
記住我






With the average age of the World's population increasing, the prevalence of age-associated diseases including neurodegenerative disorders is increasing as well. Alzheimer's disease (AD) is the most common form of dementia and the most common neurodegenerative disease affecting 12% over 65. The major pathologic hallmark of AD includes the formation of extracellular plaques consisting of amyloid-β (Aβ) peptides as well as neurofibrillary tangles formed by hyper-phosphorylated tau protein (Masters et al., 2015).
Aβ pathology has been established to precede other downstream pathologic features, suggesting that amyloid peptide secretion and aggregation are key pathogenic events in AD pathogenesis. However, the exact chain of events underlying amyloid pathology and AD pathogenesis are still not fully understood hampering the development of any curative treatments with significant clinical and socioeconomic consequences (Masters et al., 2015; Scheltens et al., 2016).
A further challenge in understanding the biochemical basis underlying AD pathogenesis is the complex, phenotypic heterogeneity of AD pathology and Aβ plaque morphology in particular. Polymorphic Aβ pathology involves structurally different amyloid aggregates within and across different plaque phenotypes (Rasmussen et al., 2017). Most prominently, mature plaques associated with AD pathology are characterized by the formation of a compact core that was found to consist mainly of Aβ 1–40 (Michno, Nystrom, et al., 2019). With background to the complexity of plaque pathology and the still limited knowledge thereof, it comes to no surprise that major pharmacotherapeutic strategies targeting Aβ plaque pathology have not been successful so far.
This, in turn, highlights the immediate need to further elucidate the molecular architecture associated with plaque formation in AD. Indeed, key developments in bioanalytical techniques, including chemical imaging, have increased our understanding of the molecular basis of single Aβ deposit formation at subcellular scales. Particularly, mass spectrometry imaging (MSI) has gained prominent relevance for studying biochemical traits of AD pathology warranted by the molecular specificity inherent to this technique (Enzlein et al., 2020; Hanrieder et al., 2013; Kaya, Brinet, Michno, Baskurt, et al., 2017; Kaya et al., 2018; Michno, Wehrli, et al., 2019 ; Michno et al., 2018). Here, matrix-assisted laser desorption ionization (MALDI)-based MSI has been demonstrated to be a valuable approach in retrieving novel chemical information on plaque pathology in both AD patient's brain and AD mouse models. This includes also molecular information on plaque-associated neuronal lipid dynamics, which is significant in that lipids have been implicated in AD pathology and more specific in Aβ plaque formation mechanisms before (Di Paolo & Kim, 2011). This is corroborated by the fact that the ε4 allele of the apolipoprotein E encoding gene (APOE), a lipid transporter, is the most prominent genetic risk factor for developing sporadic AD (Liu et al., 2013). Although the relevance of lipids in AD pathology has long been recognized, the means for elucidating lipid biochemistry comprehensively and chemically specific at single plaque scales have not been available. Here, MALDI-MSI along with complementary chemical amyloid staining to delineate and annotate plaque polymorphism, identified plaque morphology-associated sphingolipid (SP), phosphatidylcholine (PC), lysolipid (LPC/LPA), and phosphoinositol (PI), species (Kaya, Brinet, Michno, Baskurt, et al., 2017; Kaya, Brinet, Michno, Syvanen, et al., 2017; Kaya et al., 2018; Michno et al., 2018).
The capacity of MSI has been significantly expanded due to the introduction of ion mobility into the MS system allowing further separation of structurally similar and even isomeric compounds, which is of great relevance given the complexity among different lipid classes (Gillig et al., 2000; Jackson et al., 2005, 2007; McLean et al., 2007; Trim et al., 2008).
In this study, we set out to elucidate plaque morphology-associated lipid species in a transgenic AD mouse model (tgAPPSWE) that displays polymorphic plaque pathology (Philipson et al., 2010). We used a novel MALDI-MSI setup with trapped ion mobility spectrometry (TIMS) to further resolve plaque-associated lipid species. The MSI experiments were interfaced with fluorescent amyloid staining on the same section using structure sensitive, electrooptic amyloid probes, luminescent-conjugated oligothiophenes (LCOs) (Nilsson, 2009) that allow to delineate structurally different amyloid aggregates (Nilsson et al., 2007; Nystrom et al., 2013; Rasmussen et al., 2017). This hyperspectral imaging approach identified distinct lipid localization patterns specific to polymorphic amyloid structures including mass overlapping compounds, which could be further resolved by ion mobility separation.
2 METHODS 2.1 Chemicals and reagentsAll chemicals for matrix and solvent preparation were pro-analysis grade and obtained from Sigma–Aldrich/Merck unless otherwise specified. TissueTek optimal cutting temperature compound was purchased from Sakura Finetek (Cat. no: 4583; AJ Alphen aan den Rijn). Deionized water obtained by a Milli-Q purification system (Millipore Corporation).
2.2 AnimalsTransgenic male tgAPPSwe mice, 18 and 23 months of age (n = 3 per age group), carrying the Swedish (K670N, M671L) mutations of human APP were studied. The animals were reared ad libitum and housed in groups at an animal facility at Uppsala University, Sweden under a 12/12 light cycle. The animals were anesthetized with isoflurane and sacrificed by decapitation. The brains were dissected quickly within >3 min postmortem delay and frozen on dry ice. All animal procedures were approved by an ethical committee and performed in compliance with national and local animal care and use guidelines (DNr #C17/14 at Uppsala University). For this study, no randomization, blinding, and sample size calculations were performed. All animal experiments and reporting were performed according to the ARRIVE guidelines. This exploratory study was not preregistered and not randomized. No inclusion or exclusion criteria were applied. No sample size calculation was performed, though post-hoc power calculations were included for validation (see 2.6 Data Analysis).
2.3 Sample preparationTissue sections (12 μm) were collected on a cryostat microtome (Leica CM1900; Leica Biosystems) and thaw mounted onto indium tin oxide-coated, conductive glass slides (Cat. no: 237001; Bruker Daltonics GmbH & Co. KG) and stored at −80°C until further use. For matrix deposition, samples were thawed in a desiccator under reduced pressure for 30 min. 2-(1-naphthylamino) ethylamine dihydrochloride (NEDC) (Cat. no.: 222488-5G; Sigma–Aldrich) was used as a matrix and applied to the tissue sections using a TM sprayer (HTX Technologies) equipped with a P-580 HPLC pump (Dionex). Before spraying, the solvent pump was purged with 70% acetonitrile (ACN, Cat. no.: 34851-1L; Sigma–Aldrich) at 500 μl/min for 10 min followed by a manual rinse of matrix loading loop using a syringe. A matrix solution containing 7 mg/ml NEDC in 70% MeOH (Cat. no. 34860; Sigma–Aldrich) was sprayed onto the tissue sections with the following instrumental parameters: nitrogen flow (5 psi), spray temperature (30°C), nozzle height (40 mm), 14 passes with offsets and rotations, and spray velocity (1200 mm/min), and isocratic flow of 60 μl/min using 70% ACN as a pushing solvent.
2.4 MALDI-MSI analysisMass spectrometry imaging analysis of tissue sections was performed on a timsTOF fleX instrument (Bruker Daltonics). The instrument was equipped with frequency tripled, 10 kHz, Nd:YAG laser (355 nm) (SmartBeam 3D; Bruker Daltonics). Imaging MS was collected at 10 μm spatial resolution with beam scan off, using 200 laser shots per pixel. Data were collected in negative ion mode from m/z 400–2000 Da. TIMS was performed in the range from 1/K0 0.9 to 1/K0 2.13 V∙s/cm2 with a ramp time of 200 ms. External TOF calibration was performed with red phosphorous that was spotted next to the sections. TIMS was externally calibrated in electrospray ionization (ESI) mode using Tune Mix (Agilent). Single MS/MS spectra were acquired directly from tissue for some selected precursor ions at 50 µm laser pitch. For that, the mass range was set from m/z 50 to 1300 Da using a TIMS ramp from 1/K0 0.5 to 1.7 V∙s/cm2 with 200 ms ramp time. Six hundred laser shots were acquired per pixel in negative ion mode. Quadrupole isolation windows and collision energies were adapted for each precursor individually. If required, spectra from several pixels were accumulated, and collision energies were stepped from lower to higher voltages.
2.5 Hyperspectral amyloid imagingAmyloid pathology was further delineated using double staining with LCOs and hyperspectral confocal imaging. The LCOs quadro-formyl thiophene acetic acid (qFTAA) and heptamer-formyl thiophene acetic acid (hFTAA) were used for staining polymorphic plaques (Nystrom et al., 2013). Following MALDI analysis, tissues were rinsed in absolute EtOH for 60 s, fixed in absolute EtOH (Cat no.: V002075; Sigma–Aldrich) at −20°C for 8 min, 70% EtOH at −20°C for 30 s, 70% EtOH at RT for 30 s, and rinsed for 5 min in PBS (Cat. no.: P9416; Sigma–Aldrich) both prior and after staining. For fluorescent amyloid staining, tissues were incubated with qFTAA (3 μM in PBS) and hFTAA (1.5 μM in PBS) in a humidification chamber for 30 min at RT. Tissues were mounted with Prolong Gold antifade reagent (Thermo-Fisher Scientific) and dried for 2 h at RT. Hyperspectral imaging was performed on ELYRA PS.1/LSM 780 laser scanning microscopy equipped with a 32-Channel GaAsP spectral detector (Zeiss). The objective used was Plan-Apochromat 20×/0.8 (WD = 0.55 mm), ∞/0.17. The excitation wavelength for LCOs was 457 nm. The emission spectra were acquired in the range of 405–750 nm as described before (Nystrom et al., 2013).
2.6 MSI data processing and analysisImaging data were processed in MATLAB R2020b with Bioinformatics Toolbox 4.14, Signal Processing Toolbox 8.4, and Image Processing Toolbox 11.1 (MathWorks, Inc.) installed. Multivariate modeling was performed in SIMCA software (v.17.0; Sartorius Stedim Data Analytics AB—Umetrics). MALDI imaging raw data files were converted into the imzML format using SCiLS Lab (v.2021b, Bruker Daltonics) and imported into the MATLAB programming environment using the imzMLConverter by Race et al. (2012). Data were normalized to total ion count and, in addition, for multivariate modeling purposes, log-transformation was applied to address heteroscedasticity and skewness. Data were centered but not scaled for multivariate modeling. Spatial segmentation of MSI data sets was performed on score images from principal component analysis (PCA), whereby images with the most pronounced plaques or plaque core features, respectively, were used. Segmentation of plaque cores was achieved by thresholding pixel values followed by removal of off-plaque localizing pixels. Segmentation of plaque structures was achieved using Otsu's global threshold followed by a series refinement including image erosion, image dilation, active contours algorithm, and removal of off-plaque localizing pixels. Image segmentation was further guided by hyperspectral fluorescence microscopy data.
Hyperspectral imaging raw data files in czi format were imported into MATLAB using code obtained from https://github.com/CamachoDejay/czi_spec_im_load (CamachoDejay, czi_spec_im_load, 2020, github, https://github.com/CamachoDejay/czi_spec_im_load) together with the Bio-Formats software tool (Linkert et al., 2010). Binary maps of each ROI were used to subset data sets by logical indexing and averaging pixel values for statistical data analysis. Further, binary maps served as discriminator matrices for supervised multivariate analysis. Orthogonal projection to latent structure-discriminant analysis (OPLS-DA) was used to discriminate between ROIs such as plaques, plaque cores, and peripheral plaque regions. OPLS-DA is a regression method that has evolved from Partial Least Squares (PLS) where class membership is supplied in a discriminator matrix (Y-matrix) to decompose the systematic variation in the data set (X-matrix) into Y-correlated (predictive) and Y-uncorrelated (orthogonal) variation (Bylesjö et al., 2006). The separation between the predictive and orthogonal components facilitates the interpretation of the predictive loading vector and provides a direct measure of the influence each of the variables has on the model. The number of relevant components for the models was estimated through seven-block cross-validation (CV). Orthogonal components based on CV were reduced to further prevent the overfitting of the OPLS-DA models. The number of orthogonal components was then based on meaningful orthogonal variation pertaining to, for example, cores in OPLS-DA models for plaques versus nonplaque. Finally, the correlation-scaled loadings, p(corr), were examined in order to identify differences in the lipidomic profiles between areas of interest at both age groups. Post-hoc power analyses were performed in MATLAB using the sampsizepwr function (Statistics and Machine Learning Toolbox).
High-loading variables were further subjected to t-test statistics and evaluated on single-ion images. Post-hoc power analyses and t statistics were performed in Matlab using the sampsizepwr function (Statistics and Machine Learning Toolbox). Power analysis was based on a two-sample pooled t test, after assessing for normally distributed data, with unknown standard deviation and equal variances. Mean and standard deviation of sample values from image-segmented arrays were used. Although we acknowledge that the usefulness of post-hoc power analyses is controversial, we included these results as retrospective assessments to safeguard against experimental error.
Together this approach allowed for the identification of significantly altered species which were further interrogated by single-ion images based on both m/z and their collisional cross-section (CCS). For this, mass spectral and mobility information of the MALDI-TIMS imaging data were extracted using TIMS Data Viewer 1.0 (Bruker Daltonics) and visualized using SCiLS Lab 2021b.
2.7 Lipid identification 2.7.1 Lipid extraction and LC-TIMS analysesLipids were extracted according to a previously reported procedure with slight modifications (Marsching et al., 2014). Briefly, the tissue was homogenized for 2 min on ice in 2 ml methanol and 300 μl water with an Ultra Turrax T25 basic (IKA Labortechnik) at 24,000 rpm in a 15 ml polypropylene vial. The homogenate was transferred into a glass vial and the polypropylene vial was rinsed two times with 500 μl methanol. Afterward, 3 ml of chloroform (Cat. no. 650498; Sigma–Aldrich) were added to get a solvent mixture of chloroform/methanol/water (10:10:1, v/v/v). The extract was centrifuged for 10 min at 1008 g, and the supernatant was collected in a separate glass vial. Extraction was completed by repetition of this procedure twice, one time with 3 ml chloroform/methanol/water (10:10:1, v/v/v) and the second time with a 30:60:8 (v/v/v) mixture. Every extraction step included 2 min of sonication. The pooled extracts were dried under a nitrogen stream (37°C). Finally, the extract was dissolved in 100 μl chloroform/methanol/water (10:10:1, v/v/v) per 100 mg brain wet weight and stored at −20°C.
For LC-MS an ultra-high-pressure chromatography system (Bruker Elute UHPLC) was used to separate lipids on a YMC Triart C18 column (100 × 2.1 mm, 1.9 μm), packed with 1.9 μm material. The column compartment was heated to 55°C, and lipids were separated with a binary gradient at a constant flow rate of 400 ml/min. Mobile phases A and B were ACN:H2O 60:40% (v/v) and IPA:ACN:H20 90:8:2% (v/v), both buffered with 0.1% formic acid and 10 mM ammonium formate. The 20-min LC-MS experiment started by ramping the mobile phase B from 40% to 43% within 2 min, to 50% within 2.1 min, to 54% within 12 min, to 70% within 12.1 min, and finally 99% within 18 min. It was decreased to 40% within 18.1 min and kept there until 20 min to reequilibrate the column. The injection volume was 5 μl.
The MS analysis was performed in negative-ion mode as previously described using the PASEF acquisition mode (Vasilopoulou et al., 2020). Briefly, the LC was coupled to a hybrid trapped ion mobility-quadrupole time-of-flight mass spectrometer (timsTOF Pro; Bruker Daltonics) via an Apollo II ESI ion source (Bruker Daltonics). The voltage capillary was set to 4200 V and the TIMS ion charge control to 7.5 Mio counts. The accumulation and ramp time was set to 100 ms. Mass spectra were recorded in a range of m/z 100–1500, and the ion mobility was scanned from 0.55 to 1.90 V∙s/cm2. Precursors for data-dependent acquisition were isolated within the entire m/z mass range and fragmented with a constant collision energy of 35 eV. If necessary, the precursors were automatically fragmented multiple times within the same run to increase the quality of their MS/MS spectra. The mass calibration was performed using a mixture of sodium formate clusters with Agilent ESI LC-MS Tune Mix (50:50). The TIMS dimension was calibrated linearly using the same mixture.
2.7.2 Initial processing of LC-PASEF dataThe raw data were processed with MetaboScape (v. 2021b, Bruker Daltonics) using a mass range of 100–1200 m/z and a retention time range of 20 min. The feature finding algorithm T-ReX 4D automatically extracts all information in the four-dimensional space (m/z, retention time, ion mobility, and intensity) and assigns MS/MS spectra to the features. For detection, an intensity threshold of >500 counts in positive and negative ion modes was used. The m/z and mobility dimensions were calibrated in MetaboScape automatically. The minimum 4D peak size was set to 100 data points or 75 data points in three out of five replicates using recursive feature extraction. [M+H]+ ions were used as primary ions with [M+Na]+ and [M+NH4]+ as seed ions in positive and [M−H]− as primary and [M+HCOO]− ions as seed ions in negative ion mode. The positive and negative mode data were merged using a tolerance of 2 ppm for m/z and 6 s for retention times.
2.7.3 Lipid annotation based on LC-PASEF dataThe lipid annotations were performed on merged positive and negative mode data using a generic rule-based annotation tool implemented in MetaboScape. As a library-free approach, published fragmentation rules were applied to profile in total 42 lipid subclasses from four categories: glycerolipids (GL), glycerophospholipids (GP), sphingolipids (SP), and sterol lipids (SL). The precursor ion mass tolerance was set to 5 ppm, the isotopic pattern matching (mSigma) value to 250 (ranging from 0 = perfect match to 1000 = no match), and the MS/MS matching score to 450. The results were manually inspected and obvious false annotations were removed using a 4D Kendrick Mass Defect Plot analysis (plotting exact mass vs. KMD [H or CH2]), with a color coding used for retention times and the bubble sizes used to display CCS values. The generated lists of lipids were exported for further use.
2.7.4 Annotation of lipids in MALDI imagesLipids were annotated in MetaboScape (v.2021b, Bruker Daltonics) following the extraction of the four-dimensional data (m/z, ion mobility, intensity, and spatial coordinate). Briefly, spectra of the entire measurement region per sample were exported from SCiLS Lab (v.2021c) and imported into MetaboScape (v.2021b). Spectra from 10 × 10 pixels were averaged before feature extraction using the T-Rex3 processing algorithm. An intensity threshold of 200 was used for peak detection and [M−H]− ions were selected as a primary ion, seed ions were forbidden.
Features were then annotated using an analyte list of mouse brain lipids that were previously identified using LC-ESI-PASEF allowing a mass error of 5 ppm and a CCS value deviation of 3% if available.
For additional confidence, MALDI TIMS MS/MS fragment spectra from some selected precursors were analyzed with MetaboScape (v.2021b). Fragment spectra were converted to *.mgf format and added to the corresponding entry in the existing feature table in MetaboScape. The lipid identification is based on comparisons against a spectral library or molecular formula generation based on exact mass and fragment prediction.
3 RESULTS 3.1 Integrated MALDI TIMS TOF MSI and fluorescent amyloid imaging for multivariate classification of plaque pathologyWe here employed a multidimensional MSI strategy using a novel MALDI-trapped ion mobility spectrometry TOF instrument to delineate plaque pathology-associated lipid species with increased identification confidence. This is achieved through the ion mobility dimension of the used instrument that expands the molecular coverage of the MSI analyses at reasonable analysis times and most importantly increases molecular specificity through additional molecular parameters, that is, mass accuracy and CCS. Using this advanced MSI approach, we investigated plaque-associated changes in lipid localization in transgenic AD mice (tgAPPswe) from two perspectives, including amyloid plaque polymorphism and age. For this, we multiplexed the TIMS MSI analyses with complemental structure sensitive amyloid staining performed on the same section and followed plaque pathology and plaque polymorphism in tgAPPSWE mice at different ages (18 and 23 months).
In TIMS, additional data are generated across the ion mobility dimension for each mass window in addition to the m/z dimension leading consequently to very large data file sizes. To approach these complex, highly dimensional data in an unbiased way, we performed multistage image data processing and image segmentation analysis (Figure 1).

Schematic workflow for processing and statistical analysis of TIMS TOF MSI data. TIMS TOF MSI data were acquired on (a) cortical areas of transgenic AD mouse models (tgAPPSwe), (b) imaging raw data were processed, and PCA was performed in order to generate (scores) images for image segmentation. Image segmentation was performed by a combination of thresholding and active contours segmentation. Segmented image areas were either averaged for statistical analysis or were used to construct a Y-matrix for discriminant analysis. (c) OPLS-DA was performed on averaged ROI data or imaging data to examine analytes that drive class separation. (d) Results of OPLS-DA were evaluated, the identities of important loadings were determined by database search (MetaboScape), and ion images were visualized for interpretation. AD, Alzheimer’s disease; LCO, luminescent-conjugated oligothiophenes; MSI, mass spectrometry imaging; OPLS-DA, orthogonal projection to latent structure-discriminant analysis; PCA, principal component analysis; ROI, region of interest; TIMS, trapped ion mobility spectrometry; TOF, time of flight
Following data processing, we performed PCA of the data for image segmentation (Figures 1b and 2a,b). This allowed us to obtain PCA images containing plaque-specific single-ion information that we subsequently used for image segmentation. In detail, binary maps were generated for the classification of plaque structures into four regions of interest including (1) plaques and (2) background as well as (3) plaque core, and (4) plaque periphery. Binary maps were generated through a combination of thresholding and active contours segmentation to annotate the different ROI in an unbiased way across all single plaques within each tissue sample and mouse, respectively (Figure 2a–f).

Multivariate image segmentation using MSI and hyperspectral microscopy. Image segmentation for the pixel assignment was performed (a, b -zoom) on PCA scores image based on MSI data sets to generate (c, d -zoom) binary maps. This allowed for unbiased identification of Aβ plaques (e) peripheries and (f) cores. Amyloid cross β-sheet conformation-specific LCOs, including (g) q-FTAA and h-FTAA, were used for identification of more (q-FTAA) and less (h-FTAA) aggregated Aβ fibrils among Aβ plaques. Here, the (h) emission profiles of these fluorophores were verified through heat map-based hyperspectral analysis of cross-sectional emission profiles of individual deposits (i). The LCO assigned plaque (j) cores, and (k) peripheries allowed for further binary characterization of the Aβ plaques. For this purpose, the (m) LCO median spectral profiles were integrated with (n) MSI-based segmentation maps for an (o) efficient plaque structural diversity interrogation. LCO, luminescent-conjugated oligothiophenes; MSI, mass spectrometry imaging; PCA, principal component analysis
3.2 Hyperspectral microscopy with TIMS MSI reveals chemostructural heterogeneity within Aβ plaquesAlzheimer's disease pathology is highly heterogeneous, involving most prominently polymorphic amyloid deposits including senile, cored plaques, as well as diffuse plaques (Masters et al., 2015; Scheltens et al., 2016). In clinical diagnosis, a binary assessment of highly aggregated amyloid assemblies is typically performed using congo red (CR) or thioflavin. The advent of a new class of amyloid probes, LCO, offer conformation-specific detection of polymorphic amyloid structures, which revealed age-dependent changes in conformational amyloid polymorphism within individual plaques, conformation-specific properties of prions, and, most recently, variability in Aβ aggregate structures between plaques of AD subtypes (e.g., fAD, sAD) (Klingstedt et al., 2013; Magnusson et al., 2014; Nystrom et al., 2013; Rasmussen et al., 2017). Most importantly, our lab demonstrated the integration of these probes with MSI, allowing multimodal chemical and structural imaging of structurally distinct plaques (Michno et al., 2018).
In this study, we therefore multiplexed TIMS MSI with differential LCO staining for hyperspectral imaging of polymorphic Aβ at the single plaque level. Following MSI acquisition, we performed LCO double staining using q-FTAA and h-FTAA on the same tissue sections (Figure 2g). Both probes have different emission properties, allowing hyperspectral delineation of differential LCO binding (Figure 2h). Here, q-FTAA staining was chosen to allow visualization of highly aggregated, mature fibrillary structures within Aβ plaques, while in contrast, immature, protofibrillar Aβ intermediates were visualized through preferential h-FTAA binding as illustrated by cross-sectional emission profiles (Figure 2i) (Klingstedt et al., 2013; Nystrom et al., 2013). The two fluorophores were imaged together using a hyperspectral setup, allowing subsequent image segmentation into more and less-aggregated Aβ using linear unmixing-based image analysis (Figure 2j–l).
The MSI binary maps generated through PCA were then aligned with the linearly unmixed LCO data (Figure 2m–o; Figures S1 and S2). Here, the heterogeneity in the hyperspectral imaging data, that contained highly aggregated, q-FTAA-dominant features (purple), and less-aggregated forms of Aβ fibrils that were h-FTAA dominant (green) (Figure 2j–l), were used as guidance for class assignment of subsequent orthogonal projection to latent structures-discriminant analysis (OPLS-DA). MSI pixel data within the generated ROI and that were corresponding between both imaging modalities were selected for subsequent statistical data analysis.
Together, this allowed for annotation and validation of both amyloid-positive features in general (plaque vs. background) as well as plaque structural diversity based on the conformational state of the amyloid-β aggregates that predominate within a single plaque (core vs. periphery, Figure 2m–o).
Following ROI annotation and classification, the MSI pixel data were either averaged for statistical analysis or were used to construct a Y-matrix for discriminant analysis. OPLS-DA was used to discriminate between regions of interest both within and between data sets (Figure 3). Here the aim was to interrogate the lipid localization diversity accounting for differences between plaque versus background and between plaque cores and peripheries of individual plaques, respectively, and finally for differences between plaques and plaque regions of animals of 18 and 23 months of age.

OPLS-DA modeling of timsTOF MSI data on a single image basis and of plaque-averaged data. OPLS-DA on image basis for age groups 18 months (a–d) along with plaque-averaged data (number of animals n = 3, e, f). Similarly, OPLS on image data was performed for 23-month-old mice (number of animals n = 3, g–l). (a, g) Score images and corresponding (b, h) scatter plots as well as (c, d, i, j) loading plots for both the predictive components deduced from image-based OPLS-DA. (e, f, k, l) OPLS-DA modeling of plaque-averaged TIMS TOF MSI data: (e) 18 months, plaque versus nonplaque, (f) 18 months, periphery versus core, (k) 23 months, plaque versus nonplaque, (l) 23 months, periphery versus core. MSI, mass spectrometry imaging; OPLS-DA, orthogonal projection to latent structure-discriminant analysis; TIMS, trapped ion mobility spectrometry; TOF, time of flight
For this, the data were analyzed through two independent approaches. First, the data were interrogated in a Y2 matrix, where either plaque versus background or periphery versus core of the plaques was analyzed across all animals both for 18- and 23-month-old animals (Figure 3e,f,k,l; Figure S3). This analysis allowed for the extraction of key features separating the respective groups for all animals within a given age group. A limitation of this approach is the lack of possibility to visualize the unique features responsible for the separation of the fine plaque subcomponents in the context of a single animal. Therefore, we performed a second round of analysis that was based on a Y3-matrix. This approach allowed for the extraction of unique features that underlie the separation of background from plaque cores and periphery within individual animals. Here, for the features that allow classification into the respective region of interest (ROI; i.e., plaque, core, periphery), no major difference was observed between the different age groups and between the different animals, confirming the validity of the first analysis based on averaged ROI data.
This approach, however, did allow to visualize the distribution of the subgroup separating components in the form of component ion images (Figure 3a,b,g,h). The generated score images further validated the accurate ROI annotation and classification procedure and were well in line with the OPLS-DA results obtained for the averaged pixel data that were used for groupwide comparisons.
Following OPLS-DA model generation and validation, interrogation of the OPLS-DA results allowed us then to examine which analytes drive the class separation of the different ROI, within individual plaques, and between plaques in respective data sets and ages. These significant OPLS loadings (variables) were further confirmed through univariate statistics and validated by inspecting their mobility resolved single-ion localization patterns (Figures 4 and 5; Figure S4).

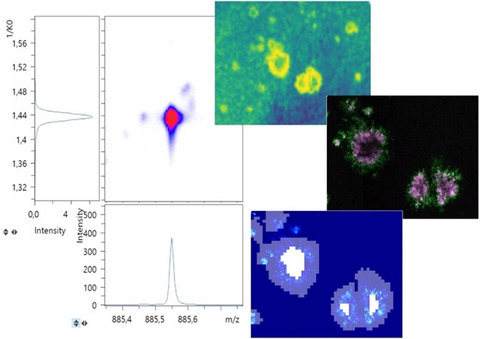
Plaque-associated lipid patterns. Extraction of collision cross-sections of higher mass range (m/z 1400–2000) from the trapped ion mobility mass spectrometry imaging allows for annotation of distinct cardiolipin (CL) (1499.996 ± 0.05 1/K0 1.987 ± 0.02 V∙s/cm2) were specifically depleted within Aβ plaques. This trend was prominent for multiple CL species in between (b) m/z 1449–1459, (c) m/z 1469–1479, and (d) m/z 1499–1509. Shorter mono-sialated gangliosides (GMs), including (e) GM3 (d36:1) (m/z 1179.728 1/K0 1.686 V∙s/cm2) and (f) GM2 (d36:1) (m/z 1382.818 1/K0 1.750 V∙s/cm2) did also display a general Aβ plaque trend with localization to these Aβ inclusions. These species were well resolved without the need for TIMS dimensionality. Finally, TIMS MSI allowed for identification of Aβ plaque-associated depletion patterns of phosphatidylserine (PS) species that otherwise partly overlapped with phosphatidylethanolamine (PE) species. These included (g) 1. PS (18:0/22:6) (m/z 834.525 1/K0 1.431 V∙s/cm2), 2. PE (22:6/22:6) (m/z 834.51 1/K0 1.564 V∙s/cm2), 3. ST (d18:1/20:0) (m/z 834.5771 1/K0 1.461 V∙s/cm2 g), and (h) 1. PE (22:6/22:1) (m/z 844.586 1/K0 1.475 V∙s/cm2g), 2. PS (22:0/18:1) (m/z 844.607 1/K0 1.461 V∙s/cm2), 3. unassigned. Number of animals n = 3. MSI, mass spectrometry imaging; TIMS, trapped ion mobility spectrometry
 Trapped ion mobility mass spectrometry imaging allows for extraction of distinct morphologic features associated with Aβ plaque cores. Here, ion mobilogram allowed for identification of core-specific localization phosphatidic acids (PA), including (a) PA (16:0/16:0) (m/z 647.464 1/K0 1.221 V∙s/cm2) and (b) PA (16:0/18:1) (m/z 673.479 1/K0 1.244 V∙s/cm2). Further, a similar localization was observed for phosphatidyl-ethanolamine species (PE), including (c) PE (34:0) (m/z 718.534 1/K0 1.289 V∙s/cm2), (d) PE (18:1/18:0) (m/z 744.554 1/K0 1.315 V∙s/cm2). Interestingly, trapped ion mobility mass spectrometry imaging allowed for identification of plaque-specific depletion of a phosphatidylserine species, (e) 1. PS (16:0/22:6) (m/z 806.501 1/K0 1.404 V∙s/cm2), and a unique localization of another lipid species putatively assigned as 2. ST (d36:1) (m/z 806.546 1/K0 1.415 V∙s/cm2). We verified the identity and side chain composition of both the PAs and the ST (d36:1) species using on tissue MS/MS analysis (Figures S5 and S6). Data indicate mean ± SD, number of animals n = 3, N = 5–10 plaques. *p ≤ 0.05, paired ratio t-test
Trapped ion mobility mass spectrometry imaging allows for extraction of distinct morphologic features associated with Aβ plaque cores. Here, ion mobilogram allowed for identification of core-specific localization phosphatidic acids (PA), including (a) PA (16:0/16:0) (m/z 647.464 1/K0 1.221 V∙s/cm2) and (b) PA (16:0/18:1) (m/z 673.479 1/K0 1.244 V∙s/cm2). Further, a similar localization was observed for phosphatidyl-ethanolamine species (PE), including (c) PE (34:0) (m/z 718.534 1/K0 1.289 V∙s/cm2), (d) PE (18:1/18:0) (m/z 744.554 1/K0 1.315 V∙s/cm2). Interestingly, trapped ion mobility mass spectrometry imaging allowed for identification of plaque-specific depletion of a phosphatidylserine species, (e) 1. PS (16:0/22:6) (m/z 806.501 1/K0 1.404 V∙s/cm2), and a unique localization of another lipid species putatively assigned as 2. ST (d36:1) (m/z 806.546 1/K0 1.415 V∙s/cm2). We verified the identity and side chain composition of both the PAs and the ST (d36:1) species using on tissue MS/MS analysis (Figures S5 and S6). Data indicate mean ± SD, number of animals n = 3, N = 5–10 plaques. *p ≤ 0.05, paired ratio t-test
Inspection of the multidimensional MSI data showed plaque ROI-associated mass peak localizations with overlapping isobaric lipid species as resolved by TIMS. Here, we made use of the additional separation dimension, trapped ion mobility, which allows us to visualize isobaric or isomeric compounds (Figures 4 and 5). Peak separation of isobaric can in theory also be achieved with high resolving power instrumentations such as FTMS. However, acquisition at these mass resolutions requires long scan times of about 2–3 s/mass spectrum. The TIMS MSI setup allows fully resolving those peaks at much shorter scan times allowing for MSI experiments to be conducted at 10% of the time.
Further, TIMS MSI provides another layer of molecular identification by providing analyte-specific CCS data along with the accurate mass. For our analysis, we made use of this feature for the identification of lipid features that were significantly different in the OPLS-DA and univariate analysis. The m/z/CCS data obtained for each statistically different lipid compound were further validated through the offline analysis of mouse brain tissue extracts using liquid chromatography coupled to ESI TIMS TOF MS/MS. The TIMS MSI data (m/z/CCS) were matched against a generated database consisting of both the m/z/CCS and MS/MS data obtained by the LC-ESI TIMS experiments (Table S1). Together, this approach generated a list of confidently identified lipid species that were statistically significant in between the different groups compared, including plaques versus background, plaque core and plaque periphery, as well as 18- and 23-month-old mice.
3.3 TIMS resolves plaque-associated lipids beyond massNext, we proceeded to inspect the OPLS-DA-derived and -identified lipid species that underlie a general Aβ plaque-associated localization pattern. Here, TIMS MSI allows for the generation of clean, single-ion images without any interfering compounds as there is no other signal visible in the heat map display. The results showed a depletion of multiple species in the higher mass range (m/z 1400–2000). Investigation of the trapped ion mobility MSI allowed for the identification of distinct cardiolipins (CL) (Figure 4a–d). Further, a general plaque-associated localization of mono-sialated gangliosides (GM), including GM2 and GM3 species was also observed (Figure 4e–f). TIMS MSI further allowed for identifying plaque-associated depletion patterns of phosphatidylserine (PS) species, PS 40:6 and PS 40:1, that partly overlapped with phosphatidylethanolamine (PE) and sulfatide (ST) species (Figure 4g,h).
3.4 Plaque polymorphism is associated with distinct lipid localization patternsInvestigation of the TIMS mobilograms underlying the core- and periphery-enriched components revealed core-specific localization of several phospholipid species, such as phosphatidic acids (PA), including PA (32:0) and PA (34:1) (Figure 5a,b). Validation through LC-MS/MS and in situ MALDI TIMS MS/MS identified the side chain composition for those PA species to be 16:0/16:0 and 16:0/18:1, respectively (Figure S5). The results further show a morphology-specific localization of PE. In particular, PE (34:0) and PE (36:1) were observed to localize to the Aβ plaque cores (Figure 5c,d). Due to the TIMS capabilities, we were able to observe a unique colocalization of PS (16:0/22:6) and a second lipid species (m/z 806.5) putatively assigned as sulfatide (ST), in particular ST (d36:1) (Figure 5e; Figure S6). We have previously reported the localization of the ST (d36:1) to the periphery and outside of the Aβ plaques (Michno et al., 2018); however, definitive assignment of the ST species was not possible due to the presence of isobaric compounds. Here thanks to the CCS value, we were able to attribute the signal depletion at the plaque to PS (16:0/22:6) species and confidently annotate the specific localization toward the plaque rim to ST (d36:1), as further by on-tissue MALDI TIMS MS/MS as ST (16:0/20:1) (Figure S6).
3.5 Progressing plaque pathology is characterized by distinct lipid accumulationFollowing these analyses, we proceeded to investigate age-related changes in plaque lipid microenvironment in 23 -months-old mice compared with 18-month-old animals. To assess changes in relative lipid localization, we compared plaque versus nonplaque tissue as well as core versus periphery within each group using OPLS-DA. Here, no differences were observed between plaque-associated and plaque-depleted lipid localization patterns when comparing the significant loading values of the models generated for 18 and 23 months, respectively. Similarly, no absolute differences of lipid changes were observed for polymorphism-related models, core versus periphery, generated for 18- and 23 -months-old animals, though differences in loading values for certain lipid species were detected. To further assess these relative changes between the two age groups, we then compared the ratio of the core/periphery signal across the ages. Here, we observed mainly changes in phosphatidylinositols (PI). In more detail, lyso-phosphatidylinositol, LPI (18:0), did not display any significant, age-related changes (Figure 6a,b), while the larger, arachidonic acid-conjugated PI (18:0/20:4) species showed a trend toward a decrease in the core/periphery signal ratio with age (p = 0.06) (Figure 6c,d). This pattern was even more prominent for a previously unreported, docosahexaenoic acid-conjugated PI (18:0/22:6) (Figure 6e,f). This lipid showed a significant difference in relative localization in between the plaque regions with a relative increase in the periphery compared with the core in 23-month-old mice compared with this ratio in 18-month-old mice. The relative signal intensity of those inositol species in between core and periphery within the age groups showed that for all three species, a more pronounced relative localization to the periphery compared with the cores in the 23-month-old compared with the 18-month-old mice (Figure 6g–l).

TIMS imaging allows for the identification of distinct phosphoinositol lipid changes associated with Aβ plaque maturation. Lyso-phosphoinositol LPI (18:0) (m/z 599.32 1/K0 1.142 V∙s/cm2) (a, b) and PI (18:0/20:4) (m/z 885.54 1/K0 1.440 V∙s/cm2) did not display age-related changes in localization to Aβ deposits (c, d), though PI (18:0/20:4) showed a trend (p = 0.06) toward relative increase in plaque periphery in older animals. PI (18:0/22:6) (e.2) (m/z 909.55 1/K0 1.455 V∙s/cm2) showed a relative increase in the periphery compared with a core of Aβ plaques in 23-month-old animals compared with 18-month-old animals. This is illustrated by a decrease in the core/periphery signal ratio (d, f). Comparative statistics within each age group show relative higher peripheral localization of PI at 23 months but not 18 months for both PI (18:0/20:4) (i, j) and PI (18:0/22:6) (k, l), while no age differences were associated for LPI (18:0) (g, h), b, d, f: Data indicate mean ± SD, number of animals n = 3/age, N = 5–10 plaques. *p ≤ 0.05, Student's t-test. g–l: Data indicate mean ± SD, number of animals n = 3/age, N = 5–10 plaques. *p ≤ 0.05, paired ratio t-test
4 DISCUSSIONThe molecular mechanisms underlying AD pathogenesis are still not fully understood. Lipids have been implicated to play a central role in AD pathogenesis. Indeed, a global alteration is observed in cerebrospinal fluid, blood, and brain tissue extracts from AD patients before (Di Paolo & Kim, 2011). Multiple studies have also demonstrated alterations of various lipids and lipid metabolites both between Aβ plaque microenvironment and even within single Aβ plaques making use of the unique capabilities of MSI (Kaya, Brinet, Michno, Baskurt, et al., 2017; Kaya, Brinet, Michno, Syvanen, et al.,
留言 (0)