
記住我
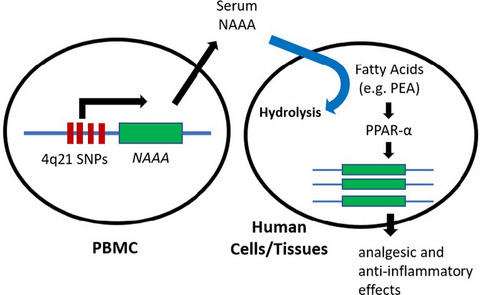



The prevalence of allergies is rising in the westernized world and is associated with urban life style.1
This is in contrast to children growing up in a farm environment and drinking farm milk who suffer less frequently from allergies and asthma.2 This effect is referred to as the “protective farm effect.” Farm life boosts gut microbiome maturation in children3 with some bacterial species (Acinetobacter lwoffii and Lactococcus lactis), and their metabolites have been associated with the farm effect.4-8 Particularly the exposure to cow sheds,9 stable dust and close contact with farm animals have been identified as main factors changing the epidemiology of allergy.10, 11
Raw milk consumption is an independent factor contributing to the protective farm effect. A broad range of factors with potential allergy protective effects have been studied to explain the protective effect of raw milk such as bovine TGF-beta, IL-10, IgG, microRNA, unsaturated fatty acids, oligosaccharides and whey protein content.12-14 In raw milk, in particular, the whey protein content has been linked to protection against allergies.13 Milk processing, especially heat treatment, coincides with loss of asthma- and allergy- protective effect of raw cow's milk, implicating the role of heat-sensitive milk components, such as whey proteins, in the tolerogenic feature and protective effects of raw cow's milk.14-16 Notably, beta-lactoglobulin (BLG) is a major whey compound, with a great similarity to the human lipocalin-2 (LCN-2) with innate immunoregulatory potency. In allergics, LCN-2 levels are reduced and can be corrected by allergen immunotherapy.17 Hence, interference with the regulatory lipocalin system with an exogenous lipocalin could be beneficial in allergy. We also detected BLG as a major compound in cow stable dust and aerosolized around cattle farms.18
In previous studies, we demonstrated that BLG prevented allergic sensitization, when fulfilling its innate function shuttling ligands to immune cells when BLG carried micronutrients within its protein pocket such as iron-flavonoid complexes,19 or retinoic acid20-22– it acted as a tolerogen, preventing antigen presentation and activating anti-inflammatory pathways.23 Importantly, transport of these micronutrients resulted in immune resilience in an antigen-unspecific manner and prevented the onset of allergy also to other allergens, thus mimicking the farm effect.24
Here, we went one step further seeking clinical translation for the new principle in the protective farm effect. Considering that farm dust and raw milk exposure need thorough time for mucosal absorption, we developed a dietary supplement as a lozenge from compounds compatible with EU regulations for food safety.25 Besides zinc18 and retinoic acid, the flavonoids catechin and epicatechin from cocoa26, 27 were chosen for complexing iron and combined with whey protein BLG derived from organic farms in EU and Switzerland.
Here, we demonstrate that the combination of the dietary components in the lozenge activated the anti-inflammatory aryl hydrogen receptor pathway (AhR) similar to the single purified components. Indeed, IgE-binding of children allergic to milk was reduced to the ligand-filled, holo-form of BLG and also antigen-specific degranulation of human mast cells was significantly diminished. The impact of the lozenge was further tested in double-sensitized mice resulting in decreased antigen presentation, promotion of regulatory T cells and reduced proliferation that was accompanied by reduced allergic symptoms to both allergens in a challenge. The results identify the new lozenge as a successful translation of the protective farm effect transporting micronutritional ligands to immune cells that initiate tolerogenic pathways in mice. The beneficial innate and allergen-non-specific effect of the lozenge has been clinically confirmed in house dust mite allergic rhinoconjunctivitis patients.28
2 MATERIALS AND METHODS 2.1 Ethics approvalSera of milk-allergic patients (10 patients positive and 10 patients negative to oral cow's milk allergen challenge) were retrospectively collected in accordance with the Helsinki declaration of 1975 and under approval of the ethical committee of the Bambino Gesù Pediatric Hospital, Rome; individual informed consent from all donors was collected by Dr. Alessandro Fiocchi, Children's hospital Bambino Gesú, Rome, Italy. Open food challenges as described in the AAAAI/Europrevall protocol29 were performed to confirm milk allergy. Children who had no symptoms with the cumulative administration of the entire milk dose corresponding to 144 mL were considered negative.
Blood cells from sixteen birch and/or grass pollen allergic volunteers (ethical approval number 1370/2018) were stimulated, as described below, with apoBLG or holoBLG and analysed by flow cytometry. All subjects provided their written informed consent under institutional review board of the Medical University approved protocols before participation. This study was approved by the institutional ethics committee of Vienna and conducted in accordance with the Helsinki declaration of 1975.
2.2 AnimalsFemale BALB/c mice, 5–7 weeks of age, were obtained from Charles River Laboratories and kept under conventional housing and treated according to European Union rules of animal care, with the permission of the Austrian Ministry of Sciences (BMWF-66.009/0133-WF/V/3b/2016).
2.3 Supplement/Placebo and raw materialsThe holoBLG lozenge (ImmunoBON®) was produced and provided by Biomedical International R+D GmbH, Austria containing active ingredients (whey with BLG, iron, catechines, vitamin A and zinc); the placebo lozenge formulated without active ingredients, consisted of sorbitol, mountain herbs flavour and karo-coffee providing a similar appearance and taste like the verum lozenge.
All compounds used were pure, organic and 100% food grade. The source of flavonoids in the holoBLG lozenge is cocoa extract. The whey concentrate was obtained from an Austrian diary company, which complies to the stringent criteria applied for food product according to EU regulations. The delivered product conformed to the current EU food regulations as well as the Austrian food law. The inspection body AT-BIO-301 made sure that the EU regulation 834/2007 was fulfilled. In addition, each whey charge was controlled for the protein composition and comparability with BLG as the major component, before using it in the lozenge production. The lozenges are produced under GMP conditions in Gebro Pharma, Fieberbrunn, Tyrol, Austria, and each constituent undergoes regular and stringent quality checks.
2.4 Allergic therapeutic modelSample size for the mouse experiments were based on the literature and described in the approved protocol of the animal ethic (BMWF-66.009/0133-WF/V/3b/2016). No randomization was performed. Female BALB/c mice, 5–7 weeks of age were split in 3 groups: (n = 8 for lozenge, n = 8 for placebo, n = 6 for naïve). Animals were immunized three times with 5 µg BLG and 5 µg Bet v 1 in conjunction with Al(OH)3 in biweekly intervals, thereafter mice received oral gavages of aliquots of crushed lozenges containing active ingredients (5 mg holoBLG using iron-catechin3 complexes as ligand; molar ratio BLG: iron: catechin = 1:1:3, 10 µg retinol and 176 µg zinc) or not (placebo-group). Mice were orally gavaged on 3 consecutive days per week over the course of 6 weeks to ensure ingestion of the crushed aliquots. Subsequently, all mice were challenged intraperitoneally with 50 µg BLG, followed 1 week later by 50 µg Bet v 1.
Body temperature and movements were monitored for 20 minutes after i.p. challenge using an Imaging system.30 The allergic symptoms of challenged animals were scored as previously described31; 0 points for no symptoms; 1 for scratching and rubbing around the nose and head, 2 for puffiness around the eyes and mouth, diarrhoea, pilar erection, reduced activity and/or decreased activity with increased respiratory rate; 3 for wheezing, laboured respiration, cyanosis around the mouth and the tail and 4 for no activity after prodding or tremor and convulsion. After euthanasia with CO2, blood was collected by cardiac puncture, and sera were stored at −80°C until further processing. Spleens were collected. Results of two separate, independent experiments were compared.
2.5 Measurement of antigen-specific antibodies in mouse serum by ELISABLG/Bet v 1-specific IgG1 and IgE levels were measured by ELISA as previously described.32 Briefly, BLG (1 µg/well) or Bet v 1 (0.5 µg/well) and serial dilutions of mouse IgG1 and IgE standards (highest concentration for IgG1, 1000 ng/ml, for IgE standard 100pg/ml) were coated in carbonate buffer, blocked with 1% BSA in PBS and incubated with diluted sera (1: 100 for IgG1 and 1:15 for IgE) overnight at 4°C. Specific antibodies were detected with monoclonal rat anti-mouse antibodies IgG1 (clone A85–1) or IgE (clone R35-72) followed by polyclonal peroxidase-labelled goat anti-rat IgG (GE Healthcare). Tetramethylbenzidine (eBioscience) was used as substrate, and 1.8 M sulfuric acid was used as stop solution followed by optical density measurement at 450 nm.
2.6 Spectral analysisFor spectral analysis, deionized water was used as buffer to minimize iron contamination from the air. The pH was kept constant at pH 7 by addition of NaOH. Optical density was measured at a constant concentration of 100 µM quercetin or 300 µM catechin with incremental increase of iron. All measurements were repeated at least three times with similar results.
2.7 Measurement of BLG-specific antibodies of milk-sensitized children by ELISAAbout 5 µg/ml apoBLG, or holoBLG diluted in 0.89% NaCl were coated on 96-well plates (100 µl/well) overnight at 4°C. After 2 h blocking at room temperature with 200 µl/well 0.89% NaCl containing +0.05% Tween 20 and 0.05% albumin (20%, 200g/L Biotest, CSL Behring), wells were incubated with 100 µl of human serum diluted 1:10 in 0.89% NaCl/0.05% Tween-20 overnight at 4°C. Detection was performed using horseradish peroxidase-conjugated goat anti-human IgE antibody (Invitrogen A18793) diluted at 1:4000 in 0.89% NaCl/0.05% Tween-20, using tetramethylbenzidine (eBioscience) as a substrate and 1.8 M sulfuric acid to stop colour development. The optical density was measured at 405 nm using an Infinite M200Pro microplate reader (Tecan, Austria). Between the steps, rigorous washing was performed with 0.89% NaCl/0.05% Tween-20.
2.8 In vitro stimulation of splenocytesIsolated splenocytes of individual mice were plated at a density of 5 × 106 cells/ml and cultured with 25 μg/ml BLG or Bet v 1 (Sigma) or medium alone for 72 h at 37°C/5% CO2.
Secreted mouse cytokines were measured with the corresponding commercial ELISAs (Invitrogen/eBioscience, for mouse IL-5, IL-10, IL-13 and IFNγ) according to the manufacturer's instructions.
2.9 AZ-AhR reporter assayAZ-AhR cells were incubated at 37°C, 5% CO2 on 96-well plates at a density of 2 × 104 cells/well for 18 h. Subsequently cells were stimulated for 18 h in triplets with 90 µM catechin/epicatechin alone or in complex with iron and increasing concentrations of BLG (10 µM). Compounds were initially incubated together for 15 minutes and the pH was adjusted to pH 7 prior addition of BLG. The positive control cells were treated with 20 nM indirubin. Cells were washed once with 0.89% NaCl, and lysis buffer was added. After a single freeze-thaw cycle, 20 μl/well of lysates were transferred into a black 96-well flat-bottom plate (Thermo Scientific) and bioluminescent reaction were started with addition of 100 μl/well of luciferase assay reagent (Promega). Chemiluminescence was measured (10 sec/well) using a spectrophotometer (Tecan InfiniteM200 PRO).
2.10 Flow cytometric analyses of murine cellsFor the evaluation of Treg cells, single-cell suspensions of murine splenocytes (0.5 million cells) were stained for CD4+CD25+Foxp3+ T regulatory cells using anti-FOXP3 PerCP-Cyanine5.5 (clone FJK-16s), anti-CD4 FITC (clone GK1.5) and anti-CD25 PE (clone PC61.5) antibodies according to the manufacturer's instructions (eBioscience). Doublets were excluded before gating the lymphocytic population, followed by consecutive gating for CD4+, CD25+ and Foxp3+.
For staining of co-stimulatory molecules on dendritic cells, splenocytes were incubated for 30 min with Calcein Violet 450 AM (Thermo-Fisher), anti-CD11c PE (clone N418), anti-MHC II I-Ad APC (clone AMS-32.1) and anti-CD86 FITC (clone GL1) in staining buffer (eBioscience) before cells were washed twice in Hepes-buffer (20 mM Hepes, 150 mM NaCl, pH 7.2). Cells were acquired on a flow cytometer, gating on CD11c+ in the living monocytic population, before plotting on MHC Class II I-Ad+CD86+ cells.
A third set was stained for CD71+ as a marker for proliferation using anti-CD71 PE (eBioscience, clone R17217) in combination with anti-CD19 APC (Biolegend, clone 6D5), and using calcein-AM (Thermo-Fisher) as a living marker. Here, first doublets were excluded, before gating on the living cells as Calcein+. Then, cells were gated on the lymphocytic population on the FSC/SSC plot, followed by gating on the CD19+ and CD71+ populations.
2.11 In vitro stimulation of peripheral blood mononuclear cellsPeripheral blood mononuclear cells (PBMCs) from sixteen birch and/or grass pollen allergic subjects were isolated via Ficoll-Paque (GE Healthcare) and washed with 0.9% NaCl, before cells were incubated with apoBLG (5 μM) or holoBLG (5 μM BLG plus 15 μM catechine and 5 μM iron) in media neither containing phenol red nor fetal calf serum for 18 hours. Only a single time-point was measured due to the technical limitations that were encountered when working with iron, as media and buffers had to be iron-free, although iron is essential for cell survival.
2.12 Flow cytometric analyses of human cellsTo determine the intracellular iron content and the expression of co-stimulatory molecules on CD14+ monocytic cells, cells were stained with combinations of Calcein Violet 450 AM (Thermo-Fisher), CD14-APC (Biolegend, clone M5EZ), HLADR-PE (Biolegend, San Diego, Calif, clone L243PC) and CD86-PE-CY7 (Biolegend, clone IT2.2). Doublets were excluded before gating on the CD14+ in the living monocytic population, followed by consecutive gating for HLADR+, CD86+ and calcein+ and geometric mean fluorescence intensity (MFI) were calcutated for each fluorochrome.
For Treg cells staining, a combination of CD3-APC-Cy7 (Biolegend, clone SK7), CD4-PE-Cy7 (Biolegend, clone SK3), CD25-APC (Biolegend, clone BC96), CD127-PE (Biolegend, clone A019D5) and Calcein Violet 450 AM (Thermo-Fisher) were used for flow cytometric analysis. Doublets were excluded, before gating on the living lymphocytic population for CD3+ and CD4+ cells and plotting for the corresponding CD25+ and CD127− cells. Treg is presented as fold expression normalized to medium and to the relative number of CD3+CD4+ T cells.
All acquisition was performed on a FACS Canto II machine (BD Bioscience, San Jose, CA, USA). Recorded events were analysed with the FlowJo software version 10.3 (FlowJo, LLC, Ashland, OR, USA).
For cytokine analyses, supernatants of stimulated PBMCs were flowcytometrically assessed for IL-2, IL-4, IL-5, IL-6, IL-10, IL-13, TNF-α and IFN-γ using multiplex system according to the manufacture's instruction (LEGENDplex™ Human Th1/Th2 Panel 8-plex, Biolegend).
2.13 Human mast cell generationHuman peripheral blood mononuclear cell-derived mast cells were generated as previously described by Folkerts et al.33 Briefly, peripheral blood mononuclear cells were obtained from buffy coats of healthy blood donors and CD34+ precursor cells were isolated using the EasySep Human CD34 Positive Selection Kit (STEMCELL Technologies). CD34+ cells were maintained for 4 weeks under serum-free conditions using StemSpan medium (STEMCELL Technologies) supplemented with recombinant human IL-6 (50 ng/ml; Peprotech), human IL-3 (10 ng/ml; Peprotech) and human Stem Cell Factor (100 ng/mL Peprotech, Rocky Hill, NJ). After 4 weeks, the cells were cultured in Iscove's modified Dulbecco's medium/ 0.5% bovine serum albumin with human IL-6 (50 ng/mL, Peprotech, Rocky Hill, NJ) and 3% supernatant of Chinese hamster ovary transfectants secreting murine stem cell factor (a gift from Dr P. Dubreuil, Marseille, France). The mature MCs were identified by flow cytometry based on positive staining for CD117 (eBioscience) and FcεRIa (eBioscience) using BD FACS Canto II (approximately 90%).
2.14 Human mast cell degranulation assaySera of children allergic or tolerant to milk were preincubated with BLG, catechin, iron alone or combinations thereof, or with dietary surrogates with whey, cocoa extract and iron before adding to primary human mast cells (0.8 Mio/ml, 40 000 cells/well) overnight. Subsequently, supernatants were collected and incubated with 200 μM 4-methylumbelliferyl-β-d-glucosaminide (4-MUG) in 100mM citric acid, pH4.5 for 1 h at 37 °C. Enzymatic reaction was then terminated by adding 0.1 M glycine buffer, pH 10.7. As positive control, cells were lysed with 0.2% Triton X-100. The β-hexosaminidase content was quantified by measuring fluorescence at ex360/em452 nm. Degranulation was assessed by measurement of released ß-hexosaminidase in the supernatant and of unreleased enzyme in the respective cell lysate. The presented results were calculated as percentage release of total ß-hexosaminidase content, with a release from unstimulated controls of 0.041%, from positive controls with anti-human IgE only of 35% and with ionomycin of 94%.
2.15 Statistical analysesMouse groups and murine cellular studies were compared by ordinary one-way ANOVA following Tukey's multiple comparisons test with a single pooled variance, for the symptom score non-parametric Kruskal-Wallis test with uncorrected Dunn's test was employed.
Concentration-dependent activation of AhR was compared with RM 2-way ANOVA, for the cellular studies with AZ-AHR cells and mast cells one-way ANOVA following Tukey's multiple comparisons test was employed.
Flow cytometric analysis of PBMCs were compared by repeated measures one-way ANOVA following the Tukey multiple comparisons test. For the binding of patient sera to different BLG forms Wilcoxon matched-pairs signed test was applied. All tests were two-sided and considered significant when p < .05.
3 RESULTS 3.1 BLG binds to iron-catechin complexesCatechin with a complex stability constant log ß of 47.4 at physiological pH34, 35 binds strongly to ferric iron. Indeed, binding to ferric iron leads to colour formation and can be monitored by UV-VIS spectroscopy (Figure 1B). Importantly, BLG binds strongly to these iron(catechin)3 and iron(epicatechin)3 complexes with calculated affinity constants reaching the lower nM-range with 16.7 and 46.1nM respectively (Figure 1A, C). This leads to a discolouration, which is visible macroscopically and spectroscopically. The data confirm that the flavonoid catechin at physiological aerobic conditions is usually present in complex with iron and that BLG is able to bind to these compounds.

Beta-lactoglobulin binds to iron-catechin complexes. (A) Protein surface of beta-lactoglobulin, BLG, incorporating iron(catechin)3 (FeCat3) (sticks with carbons in deep blue, oxygens in red and iron shown as a grey sphere) and calculated affinities of catechin/epicatechin in conjunction with iron to BLG. (B) Optical spectra of 300 µM catechin with increasing concentration of ferric iron at pH 7.3. (C) Optical spectra and colour change of 150 µM catechin, 50 µM iron, BLG and combinations thereof
3.2 Ligand binding masks B and T cell epitopes of BLG affecting IgE binding and antigen-specific mast cell degranulationAs previously published with quercetin as ligand,23 also catechin (Figure 2) and epicatechin (Figure S1) clearly affected the experimentally deduced epitope regions of BLG. When we compared the ligand-binding site with the two described dominant IgE epitope regions at residues K75-D85 located in the loop and E127-P144 at the site of the alpha helix26 and the “weaker” IgE epitopes described at segments L31-P48, K47-K60, and L57-I78, it became apparent that also the iron(catechin)3 complex was in near proximity to the major B cell epitope 1 (Figure 2A), but not of epitope 2, which is located at the opposite site of epitope 1 on the alpha helix. Importantly, we determined the clinical relevance of epitope masking, by comparing IgE binding to BLG with or without catechin-iron ligands, in milk-sensitized children who reacted positive (n = 10) and negative (n = 10) to oral cow milk challenge, termed milk allergic and milk tolerant respectively (Figure 2B,C). Specific IgE levels of milk-allergic subjects were significantly higher than of milk-tolerant children. Interestingly, the milk-allergic subjects had reduced IgE binding to the holo-form of BLG, but not the milk-tolerant individuals, emphasizing that children tolerant to milk recognize different epitopes than children allergic to milk.

Ligand binding masks B cell and T cell epitopes of BLG affecting IgE binding and mast cell degranulation. (A) Structure of BLG-Fe(catechin)3 complex shown as cartoon (left) and as molecular surface (right). Fe(catechin)3 is depicted as sticks with carbons in cyan and oxygens in red. Iron atom is represented as a grey sphere. Major B cell epitope 1 (75–85) and epitope 2 (127–144) are marked in marine blue and light blue, respectively. The T cell epitope (101–112) is marked in green. Residues within a 3.5 Å distance from any atom of Fe(epicatechin)3 are shown as sticks with carbons in pink. Two residues within a 3.0 Å distance from any atom of Fe(catechin)3 (39 and 105) are shown as sticks with carbons in deep red. The third residue within a 3.0 Å distance from the ligand, D85, is also part of the B cell epitope 1 and is shown as sticks with carbons in violet purple. (B) IgE binding to BLG alone or in combination with iron-catechin in milk-allergic (n = 10) and (C) milk-tolerant children (n = 10) respectively. Degranulation of human mast cells sensitized with pooled milk-allergic sera and (D) crosslinked with BLG alone or in combination with iron-catechin complexes or (E) crosslinked with combinations of dietary agents such as whey containing BLG, cocoa containing catechin and iron alone. IgE binding was compared in B and C by Wilcoxon matched-pairs signed test and for D and E by one-way ANOVA following Tukey's multiple comparisons test. Mean ± SEM; *p < .05, **p < .01, ***p < .001; ****p < .0001
Ligand binding also affected the immune-dominant T cell epitope at residues 97–117 of BLG as also described for retinoic acid20 and for quercetin23 with the most important core residues spanning from Y102-E112 (YLLFCMENSAE).36, 37 As depicted in Figure 2A, catechin-iron complexes co-localized with the T cell epitope residues at position L39 and F105 being within a 3 Å proximity, and L32, V41, I56, K70, I71, N88, M107, A118 and Q120 being within 3.5 Å. The T cell epitope masking may alter T cell activation as described for the major birch pollen allergen Bet v 1 in two studies38, 39 as endolysosomal enzymes are hindered in generating antigenic peptides for antigen presentation. Among endolysosomal proteases, cathepsin S, predominantly expressed in antigen presenting cells, has two predicted cleavage sites, at BLGp99-105 and BLGp109-115.20 Both are located within the minimum essential region of BLG (YLLFCMENSAE) suggesting that also here the position F105 is occupied by the ligand and hinders cleavage.
We also addressed whether the ligands had an impact on the effector phase, using human mast cells. Pooled sera of children allergic to milk were preincubated with BLG alone or in combination with iron-catechin to form IgE-antigen complexes prior to addition to human mast cells. Similarly, we used dietary agents such as whey as a source for BLG, cocoa for catechin and iron to form IgE-antigen complexes prior to addition to the cells. Antigen-specific degranulation was significantly reduced when crosslinking was conducted using BLG in conjunction with iron-catechin implying again that the holo-form of BLG filled with ligand was less allergenic than the empty apo-form. Also, when using dietary agents as surrogate for BLG, catechin and iron, a similar and significant reduction in mast cell degranulation was observed.
Hence, B cell epitope masking resulted in reduced IgE binding and mast cell degranulation, whereas T cell epitope masking interferes with cathepsin S cleavage upon endosomal entry and alters antigen processing and in consequence proper antigen presentation and potentially T cell activation.
3.3 BLG facilitates AhR activation by (epi)catechin-iron complexesAs our dietary supplement lozenge contained cocoa extract as a source for BLG ligands, which is rich of the flavonoids epicatechin and catechin, we assessed whether anti-inflammatory pathways such as the cytoplasmic arylhydrocarbon receptor (AhR) were activated by these flavonoids. AhR is described to interact with many exogenous ligands40 and to mediate primarily anti-inflammatory feedback mechanisms41 that promote regulatory T cells,39, 42, 43 while preventing Th2-skewing44, 45 and impairing antigen presentation.46 Using reporter cells, we could demonstrate that catechin and epicatechin could concentration-dependently activate AhR and interestingly activation was significantly enhanced when they were present as a complex with iron (Figure 3A) implying active transport of the complex. Similarly, to our previous report, addition of BLG significantly enhanced (epi)catechin-dependent activation of the AhR pathway suggesting synergistic pathways which directed targeting to the putative BLG receptors (Figure 3B). We also tested whether the single constituents of the lozenge and combinations thereof were able to activate the AhR pathway (Figure 3C). Indeed, similar to other reports,47 the whey concentrate alone was able to activate AhR, pointing towards already present ligands in the whey concentrate, whereas neither cocoa extract, iron nor the cocoa-iron combinations were able to initiate this pathway. Importantly, combination of all mentioned constituents again significantly enhanced activation of the AhR pathway compared with whey alone, confirming that BLG serves as a carrier for flavonoid-iron complexes able to activate AhR.

The AhR activation by (epi)catechin-iron complexes is increased upon addition of BLG. AZ-AhR cells were treated with (A) increasing concentrations of catechin (left graph) or epicatechin (right graph) alone (0−60 µM), or with addition of iron (B) with 60 µM of catechin (left) or epicatechin (right) in combination with 20 µM iron and/or 2 µM BLG, (C) equivalent concentrations of dietary agents as source for iron (iron citrate, 10 µM), epicatechin/catechin (cocoa extract, aprox. 30 µM catechin/epicatechin) and BLG (whey concentrate containing approx.10 µM BLG) for 18 h before luciferase-activity was measured in the supernatant. Representative data from two independent experiments normalized to medium alone. Concentration-dependent activation of AhR in graph A was compared with 2-way ANOVA, graph B and C with one-way ANOVA following Tukey's multiple comparisons test. Mean ± STD; *p < .05**p < .01, ***p < .001, ****p < .0001
3.4 Dietary supplementation with verum lozenge containing holoBLG reduced clinical reactivity in vivoWe analysed whether holoBLG with catechin-iron complexes as ligands in a dietary supplement could be used therapeutically. A lozenge containing BLG, iron-catechins as well as zinc and vitamin A were formulated to be suitable also for human use.
Mice were simultaneously sensitized with alum against BLG and Bet v 1, before treating them orally with crushed aliquots of verum lozenges containing 5mg BLG with iron-catechin complexes, 10 µg retinol and 175 µg zinc, or placebo lozenges devoid of active ingredients. Allergic reactivity was analysed by systemic challenges first with BLG and in a second step also with Bet v 1 (Figure 4A).

Oral supplementation with a lozenge containing holoBLG reduces clinical reactivity in mice. (A) As depicted in the treatment scheme, mice were sensitized 3x with BLG and Bet v 1 in combination with Alum, before receiving in 6 cycles aliquots of crushed lozenges containing active ingredients (5 mg holoBLG in verum lozenge) or not (placebo lozenge). Subsequently, mice were challenged with BLG and Bet v 1 one week apart; (B) BLG- and Bet v 1-specific antibodies before and (C) after oral treatment; (D) clinical response after BLG and Bet v 1 challenge of double-sensitized mice after treatment with immune or placebo lozenges. Data from one experiment are shown (n = 8 for lozenge groups, n = 6 for naïve group). Groups were compared by one-way ANOVA followed by Tukey's multiple comparison, for symptom score analyses ANOVA followed by Kruskal-Wallis test was used. *p < .05, **p < .01, ***p < .001
Treatment in the verum lozenge group resulted in a significant decrease of BLG-specific IgG1 only, and Bet v 1-specific IgG1 and IgE antibodies (Figure 4B,C). Though BLG-specific IgE did not differ after verum supplementation, allergic mice treated with holoBLG, but not with the placebo substances, were protected against anaphylaxis to both antigens BLG and Bet v 1 (Figure 4D).
3.5 Lozenge treatment decreased immune reactivity by promoting tolerance: less lymphocytic and B cell proliferation, more Tregs and decreased antigen presentation in vivoIn line with a suppressed immune response, treatment with the verum lozenge rendered comparable splenocyte numbers as the naïve group (Figure 5A). Also, the expression of the proliferation marker CD71 on lymphocytes and B cells did not significantly differ from the naïve group (Figure 5B,C), in contrast to the group treated with the placebo lozenge. However, splenocytes of double-sensitized mice still had the capacity to respond similarly to in vitro stimulation with BLG and Bet v 1, irrespective whether the groups were treated with the verum or placebo lozenges (Figure S2). Regulatory T cells were significantly elevated after verum lozenge treatment compared to the placebo lozenge-treated group (Figure 5D) presumably in an antigen-unspecific manner. However, the data on the verum lozenge treatment were accompanied by impaired antigen presentation, in line with a decreased expression of co-stimulatory markers (Figure 5E). Hence, supplementation with the verum lozenge established immune resilience in an ongoing Th2-response.

Treg promotion, decreased antigen presentation and decreased B cell proliferation upon treatment with the verum lozenge. (A) Total splenocyte numbers; (B) Lymphocyte proliferation using CD71 as proliferation marker (C) B cell activation (D) Foxp3 expression of CD4+CD25+ splenocytes; (E) % of CD11c+CD86+MHCII+splenocytes. Groups were compared by one-way ANOVA followed by Tukey's multiple comparison. *p < .05, **p < .01, ***p < .001, ** p < .0001
3.6 Iron transport by holoBLG into CD14+ monocytic cells is accompanied by decreased antigen presentationIn a next step, we assess whether similar results could be obtained when incubating PBMCs from pollen allergic patients with apoBLG or holoBLG for 18h. Co-stimulatory molecules and intracellular iron content of CD14+ cells were assessed flow cytometrically. Similar to previous published data using iron-quercetin complexes as BLG’s ligand,23, 24 here we demonstrated that transport of iron-catechine complexes by BLG increased the intracellular iron content of CD14+ populations, reduced relative numbers of CD14+ monocytic cells (Figure 6A) and significantly suppressed the expression of the co-stimulatory molecules, HLADR+and CD86+ (Figure 6B,C) compared with cells stimulated with apoBLG. The data further support our therapeutic murine model showing that dietary supplementation with holoBLG and catechin-iron complexes was sufficient to ameliorate clinical reactivity in vivo irrespective to which allergens mice were sensitized to. It emphasizes the function of BLG in providing antigen presenting cells with micronutrients and thereby blocking immune activation. Mechanistically, targeting antigen presenting cells and shuttling anti-inflammatory ligands into these cells lead to reduced antigen presentation and less proliferation in an antigen-non-specific manner.

HoloBLG stimulation of peripheral blood mononuclear cells increase the intracellular iron levels of CD14+monocytes, while decreasing antigen presentation and the relative number of Th2 cells. Flow cytometric analyses of PBMCs (n = 16) stimulated with medium, iron-catechine, apoBLG alone or in combination thereof (holoBLG). (A) Relative numbers of CD14+ cells in the monocytic gate and the respected mean fluorescence intensity (MFI) of the calcein signal. An increase of intracellular iron results in a lower calcein signal due to quenching. (B) HLADR expression as well as calcein signal of CD14+HLADR+cells (C) CD86 MFI of CD14+ HLADR +cells and the respected calcein signal of these cells. (D) The relative number of CD3+CD4+ cells and (E) calcein MFI of CD3+CD4+ cells as well as (F) n-fold expression of CD25+CD127− cells in CD3+CD4+ cells compared with cells incubated to medium alone. Groups were compared by repeated measures one-way ANOVA following the Tukey multiple comparisons test. *p < .05; **p < .01; ***p < .001; ****p < .0001; ns = non-significant
Moreover, in line with previous reports,19 the relative number of CD3+CD4+ T cells was significantly up-regulated in cells stimulated with apoBLG (Figure S3a). Relative CD3+CD4+ numbers did not differ upon holoBLG treatment from medium control, whereas the iron-catechine complex alone was sufficient to reduce CD3+CD4+numbers which may indicate that particularly T helper cells were sensitive to the catechine-iron complex. Also, overnight incubation with holoBLG significantly promoted regulatory T cells, (Figure S3b) though also here the data suggest a role of the catechine-iron complex in this process.
Cytokine analysis (Figure S4) revealed that, in particular the release of pro-inflammatory cytokine TNF-α was significantly reduced upon overnight stimulation with holoBLG but not with apoBLG. Additionally, released IL-4 and IL-6 levels tended to be lower when cells were incubated with holoBLG, whereas IFN-γ, IL-10 and IL-5 levels did not differ in cells treated either with apo- or holoBLG.
Summing up, the increased iron levels in antigen presenting cells went along with decreased expression of co-stimulatory markers and an increase of T cells with a supposed regulatory phenotype upon treatment with holoBLG. In this study, iron-catechine complexes were used as ligands for BLG to provide antigen presenting cells with micronutrients and to promote an immune resilience state.
4 DISCUSSIONWe have previously shown that sensitization to BLG can be prevented when BLG is filled with ligands, as they initiate anti-inflammatory pathways leading to tolerance induction and immune resilience.23
Here, we went a step further and addressed whether the concept of holoBLG, which is a novel player within the protective farm effect, may be applied and exploited for allergic subjects by using BLG as a carrier for micronutrients, nourishing antigen presenting cells in a non-antigen-specific manner. We formulated a lozenge containing food and dietary supplements approved for human use: whey concentrate as a source of BLG, cocoa as a source rich of epicatechin and catechin, furthermore ferric iron, retinoic acid and zinc.
We confirm here that catechins are strong iron chelators, activate the AhR pathway and most importantly that BLG can bind to these complexes and facilitate activation of AhR pathway, irrespective of whether it is co-applied with whey concentrate or applied in pure form. Similarly, as with other ligands, we show for the holo-form of BLG reduced IgE binding and mast cell degranulation with serum from children allergic to milk. Importantly, the ligands mask a major IgE epitope48 and a well-described T cell epitope.20, 35, 47 This suggests that children allergic to milk at one point were sensitized to BLG-devoid of any ligands, the only setting when IgE can be generated to a region which otherwise would be masked by the ligand.
Oral supplementation of allergic mice with the lozenge containing holoBLG led to reduced allergic reactivity in an antigen-unspecific manner as clinical symptoms improved not only against BLG but also towards Bet v 1. This effect was accompanied by reduced proliferation, antigen presentation and promotion of regulatory T cells. Importantly, this is in agreement with our previous results, when the single component BLG complexed with iron-quercetin applied via the nasal route prevented allergy development in an antigen-unspecific manner in vivo.24 The data are also in line with our in vitro generated data with human immune cells23 showing that holoBLG shuttled catechin-iron complexes specifically into monocytic cells and thereby hindered their activation and maturation. It provides also a rational for the antigen-unspecific effect against allergies as the monocytic cells provided with micronutrients are the antigen presenting cells that link the innate with the adaptive immune branch.
Particularly, the provision of iron seems pivotal in modulating an immune response. The iron status deeply affects immunity, with iron deficiency known to cause immune hyperactivity49, 50 prone initially towards Th251-53 and being associated with atopic dermatitis,54 allergic rhinitis55, 56 and asthma.57-61 In contrast, in iron sated conditions, immune cells seem to shift towards a more resilient state with macrophages polarizing towards an anti-inflammatory M2 sub-type in vivo and in vitro,62 antigen presentation being decreased,23, 63 CD4+ counts64 and to a lesser extent CD8+ cells being negatively affected.65
The lozenge was also spiked with vitamin A and zinc, essential contributors to immunity: Vitamin A supports mucosal regulation and predominantly anti-inflammatory pathways,22 whereas zinc deficiencies potentially cause lymphopenia, as zinc functions as a second messenger for innate immunity. Zinc efflux is required for dendritic cell maturation and zinc influx for macrophage activation.66 Importantly, we have described vitamin A and zinc have been described as natural ligands for BLG before.18, 20
Similarly, numerous studies have shown that initiation of the aryl hydrocarbon receptor–highly expressed in immune cells capable of antigen presentation such as monocytes,67 dendritic cells, macrophages68 and B cells
留言 (0)