
記住我









Fully differentiated, pigment producing melanocytes are integral cellular components of human skin, and they are considered essential in UV protection. They differentiate from the neural crest, and the master transcriptional regulator in its full differentiation is the microphthalmia-associated transcription factor (MITF). MITF regulates three major pigment enzymes, tyrosinase, and tyrosinase related protein-1 and 2 (TRP-1 and TRP-2) (Widlund & Fisher, 2003). Fully differentiated, pigment producing melanocytes are localized in the interfollicular epidermis surrounded by keratinocytes, while precursor melanoblasts are found in the hair follicle, and melanocyte lineage stem cells reside in the bulge area (Nishimura et al., 2002). Mast/stem cell growth factor receptor Kit (c-Kit), apoptosis regulator Bcl-2 (BCL2) positive, and TRP1 negative (Kit+Bcl-2+TRP1−) cells in the follicular infundibula represent precursor melanocytes (Grichnik et al., 2006). c-Kit is essential in melanocyte differentiation, and anti-KIT antibody inhibits melanocyte proliferation and differentiation (Botchkareva et al., 2001). Activation of c-Kit by stem cell factor results in MITF phosphorylation (Hemesath et al., 1998). Notch signaling is required for maintaining melanoblasts and melanocyte stem cells (Moriyama et al., 2006). Transcription factor SOX10 (SOX10) is required for maintaining melanocyte precursors in the bulb (Harris et al., 2013).
In previous work, we have established an in vitro culture for human adult epidermal melanocytes in which melanocytes dedifferentiated. The culture lacked specific mitogens, the tumor promoter 12-O-tetradecanoyl phorbol-13-acetate (TPA or PMA), the intracellular cyclic adenosine 3′, 5′ monophosphate (cAMP) enhancer cholera toxin (CT), and 3-isobutyl-1-methylxanthine (IBMX), generally used in in vitro melanocyte cultures. Under our culture condition, melanocytes became bipolar, unpigmented, and highly proliferative in vitro. c-Kit and TRP-1 expression disappeared, and nestin and epidermal growth factor receptor (EGFR) expression was induced in the cells. These cells could be passaged up to 15 passages before they reached complete senescence (Szabad et al., 2007). We also showed that this in vitro dedifferentiation was reversible, and both addition of CT, PMA, and indirect UVB irradiation, using irradiated keratinocyte supernatant, induced redifferentiation.
Phenotype switching in melanoma is well established and thought to play a crucial role in therapy resistance, dormancy, and metastasis (Rambow et al., 2019). Our cultured cells represent a good system to investigate melanocyte phenotypic plasticity; therefore in this work, we aimed to further characterize these in vitro dedifferentiated melanocytes and to detect intrinsic mechanisms governing cellular dedifferentiation by comparing the dedifferentiated cells to conventionally cultured pigment producing melanocytes using RNAseq.
2 MATERIALS AND METHODS 2.1 Skin samples and ethicsSkin samples from healthy adult donors undergoing plastic surgery procedures (n = 11; years: 23–52) were collected after written informed consent in accordance with the ethical standards of the Helsinki Declaration and corresponding to the EU Member States' Directive 2004/23/EC on tissue isolation. Sample collection was approved by the Human Investigation Review Board of the University of Szeged (PSO-EDAFN-002, 34/2015, 3517, 23 February 2015, Szeged, Hungary).
2.2 Cell cultureM254 Medium was used as a commercially available melanocyte medium (Biosciences, Dublin, Ireland) supplemented with Human Melanocyte Growth Supplement (HMGS) containing 0.2% bovine pituitary extract (BPE), 0.5% fetal bovine serum (FBS), 0.01 µg/ml recombinant human insulin-like growth factor-I, 5 µg/ml bovine transferrin, 3 ng/ml basic fibroblast growth factor, 0.18 µg/ml hydrocortisone, 3 µg/ml heparin, 10 ng/ml phorbol 12-myristate 13-acetate (PMA) (all from Biosciences, Dublin, Ireland), and 1% Antibiotic/Antimycotic solution (AB/AM) consisting of 100 U/ml penicillin, 100 µg/ml streptomycin, and 0.25 µg/ml amphotericin B (Biosciences).
Mel-mix culture medium was previously described (Kormos et al., 2011) as an alternative culture method in which melanocytes underwent dedifferentiation. Mel-mix medium contains 5% FBS (Euroclone), 47.5% Keratinocyte Serum Free Medium (SFM) supplemented with 2.5 µg epidermal growth factor and 25 mg BPE (Biosciences), 1% L-glutamine (Biosera), 1% AB/AM (Biosciences), and 47.5% AimV medium (Biosciences) supplemented with 1% L-glutamine and 1% AB/AM solution (Biosciences).
Melanocytes were isolated from normal healthy adult skin as described previously (Szabad et al., 2007). Skin samples were first washed in an AB/AM solution (Biosciences). The subcutis was removed by surgical scissors, and the sample was cut into small strips. Next, the tissue was incubated overnight in Dispase II (Lonza) solution at 4°C to separate the epidermis from the dermis. Subsequently, the epidermis was trypsinized (Biosera) and was vigorously washed to release epidermal cells. The epidermal cell suspension was filtered and centrifuged. The cells were then plated onto 75 cm2 tissue culture dishes (BD Biosciences). In Mel-mix, the melanocytes usually attach in 24 hr, while keratinocytes can attach later between 24 and 48 hr after plating. We consequently washed the plates between 12 and 24 hr after plating to remove keratinocytes and supplied fresh medium to the cells. During trypsinization, the melanocytes detach from the flask at a higher rate than keratinocytes due to their different attachment properties as previously described (Kormos et al., 2011). After the 2nd passage, no keratinocytes remained in the culture. The isolated cells were grown either in M254 cell culture medium (Life Technologies) or in Mel-mix medium. The melanocytes cultured in Mel-mix reached ~90% confluency in 7–9 days, while the cells sustained in M254 reached the same confluency in 10–13 days. Melanocytes cultured in M254 medium were grown until the 3rd passage was reached, while cells cultured in Mel-mix were grown for 9–11 passages. When the cultures reached confluency, they were treated with 0.05% ethylenediaminetetraacetic acid (EDTA) in phosphate buffered saline (PBS) and cells were harvested by a short, 2–3 min trypsinization. Harvested cells were counted and were seeded into new flasks. Cultures were grown in an incubator at 37°C in humidified atmosphere containing 5% CO2.
2.3 RNAseq methodsTotal RNA was isolated from dedifferentiated and mature melanocytes cultured in 75 cm2 cell culture flasks (Corning) and collected at the 10th passage in the case of dedifferentiated melanocytes and mature melanocytes were harvested at the 3rd passage using TRI-Reagent (Molecular Research Center) according to the manufacturer's instruction. To obtain global transcriptome data, high throughput mRNA sequencing analysis was performed. Total RNA sample quality was checked on Agilent BioAnalyzer using Eukaryotic Total RNA Nano Kit (Agilent) according to manufacturer's protocol. Samples with RNA integrity number (RIN) value >7 were accepted for library preparation process. RNAseq libraries were prepared from total RNA using Ultra II RNA Sample Prep kit (BioLabs) according to the manufacturer's protocol. Briefly, poly-A RNAs were captured by oligo-dT conjugated magnetic beads; then, the mRNAs were eluted and fragmented at 94°C. First strand cDNA was generated by random priming reverse transcription, and after the second strand synthesis step, double-stranded cDNA was generated. After repairing ends, A-tailing and adapter ligation steps, adapter ligated fragments were amplified in enrichment PCR and finally sequencing libraries were generated. The sequencing run was executed on Illumina NextSeq500 sequencing platform (Illumina) using single-end 75 cycles sequencing.
2.4 RNAseq data analysisRaw sequencing data (fastq) were aligned to human reference genome version GRCh38 using HISAT2 algorithm, and BAM files were generated. Downstream analysis was performed using StrandNGS software (www.strand-ngs.com). BAM files were imported into the software, and DESeq1 algorithm was used for normalization. To identify differentially expressed genes between conditions, Moderated t test with Benjamini-Hochberg FDR for multiple testing correction was used. We used Fisher's exact test to see that genes related to melanocyte differentiation had a significant change out of all gene expression changes.
2.5 Ingenuity Pathway AnalysisTo identify the relationships between selected genes, the Ingenuity Pathway Analysis (IPA; Ingenuity Systems) was used. The Ingenuity Pathways Knowledge Base (IPKB) provided all known functions and interactions which were published in the literature. ANOVA was used to calculate a p-value to determine the probability that each biological function or canonical pathway assigned to the data set was due to chance alone. For the representation of the relationships between the genes, the “Pathway Designer” tool of the IPA software was used.
2.6 ImmunohistochemistryMel-mix-cultured dedifferentiated melanocytes in 9th passage and mature melanocytes in 3rd passage were used for cytospin preparations in the immunohistochemistry studies. Samples were made with Wescor Cytopro 7620 Cytocentrifuge (Wescor Biomedical Systems) at 600 rpm for 6 min and then fixed in 4% paraformaldehyde (PFA). The staining for intracellular transcription regulators SOX10 (BioCare Medical) and MITF (Santa Cruz Biotechnology) and for transmembrane protein E-cadherin (CellMarque) were made by a Leica Bond Fully automated Immunostainer (Leica Biosystems Inc.). Briefly, the endogenous peroxidase activity was blocked for 3 min and primary markers were applied for 20 min. The post-primary antibody (Leica Biosystems Inc.) was introduced for 8 min, and the samples were washed in BOND™ Wash Solution (Leica Biosystems) for 15 min. The samples were exposed to Mixed DAB refine solution for 10 min and then labeled by hematoxylin (all from Leica Biosystems). A Zeiss Axio Imager Z1 microscope (Carl Zeiss AG) was used for observation.
2.7 Flow cytometryMature melanocytes at the 3rd passage and dedifferentiated melanocytes from the cultures were collected at the 10th passage, and the phenotype of cells was identified by three-color flow cytometry. Briefly, cells detached by a short trypsinization, washed with PBS-EDTA (1%) solution and fixed in 4% paraformaldehyde (PFA) and stained for 30 min with fluorochrome-conjugated antibodies, and then measured on a BD FACSAria Fusion flow cytometer (BD Biosciences Immunocytometry Systems). Data were analyzed using Flowing Software (Cell Imaging Core, Turku Centre for Biotechnology), and the results were represented as mean percent of positive cells (%) ± standard deviation (SD). We used the following antibodies: anti-human cluster of differentiation (CD) 34, CD73, CD146, CD90, CD105, CD47, CD44, CD29, CD325, CD54, CD31, CD271, CD6, CD106, CD112, CD61, CD166, CD50, CD40, CD49A, CD51, CD59D, CD144, CD304, HLA-DR, and anti-human IgG1 and IgG2 as isotype controls (all from Biolegend).
2.8 In vitro scratch assayDedifferentiated melanocytes and mature melanocytes were plated onto a 48 well plate (Corning Life Sciences). Initially, 50,000 cells per well were seeded and 100% confluent cultures were scratched with a 200 µl pipette tip and were washed with PBS. To examine only the migration capacity of the cells, Mitomycin C (Sigma- Aldrich) treatment was also used. The culture medium was changed every two days. The wound closure was monitored by light microscopy with Zeiss Axiolab Vert.A1 microscope (Carl Zeiss AG).
2.9 Differentiation experimentsThe in vitro propagation of dedifferentiated melanocytes toward bone, cartilage, and fat tissues was carried out with STEMPRO®️ (Biosciences) differentiation kits. For osteogenesis and adipogenesis, 2.5 × 104 cells were plated onto each well of a 48 well cell culture plate (Greiner Bio-One) and were grown in adipogenesis and osteogenesis differentiation medium for 21 days. Finally, the cells were fixed in PFA (4%) for 30 min and stained with 4′, 6-diamidino-2-phenylindole (DAPI) (Sigma-Aldrich) to visualize nuclei. Nile Red (Sigma-Aldrich) was used for detection of lipid-laden particles and Alizarin Red S (Sigma-Aldrich) for the demonstration of calcium deposits. For chondrogenesis, micromass cultures were used, where 1.6 × 106 cells were used in a 15 ml Falcon tubes to form spheroid. Medium was changed in every alternate day. After 14 days of differentiation, the cells were fixed in PFA (4%) washed with 1 × PBS, and paraffin embedded. 2.5 µm wide sections were cut, and Alcian blue–van Gieson staining was performed. The sections were scanned with Panoramic MIDI II (3DHISTECH Ltd.).
2.10 KaryotypingFor karyotyping, dedifferentiated melanocytes were cultured in T75 flasks in the 10th passage at ~75% confluency and cells were blocked with a demecolcine solution (Sigma) for 5 hr. Cells were harvested with a cell scraper and then centrifuged at 200 g for 15 min. The samples were treated with a hypotonic solution (KCl 0.56m/m%) for 40 min, fixed with Carnoy's solution (acetic acid and methanol [1:3]) for 10 min, and centrifuged at 250 g for 10 min, and this was repeated two more times. After that, cells were incubated at −20°C overnight and fixed two more times using Carnoy's solution. The samples were dropped on a glass slide, trypsinized for 10 s with a 0.0025 m/m% solution, and then washed with PBS. Samples were ultimately stained with Giemsa (Sigma) solution for 2 min. Leica DM6 B microscope and CytoVision (Leica Biosystems Inc.) was used to capture images and sort chromosomes.
2.11 Western blotTotal protein samples were made from mature melanocytes (MM) at the 3rd passage and dedifferentiated melanocytes (DM) at the 10th passage. After aspiration of cell culture medium, the samples were thoroughly washed with PBS. Cells were harvested by scraping the cultures and then centrifuged at 540 g for 20 min at 4°C. Protein extraction was done by protein lysis buffer (20 mM HEPES, 150 mM KCl, 1 mM MgCl2, 1 mM dithiothreitol, 10% glycerol, 0.5% Triton X-100, 0.1% Igepal CA-630) containing 1% protease inhibitor cocktail and 1% PMSF (Phenylmethanesulfonyl fluoride) (all components from Sigma). Protein concentrations were determined using the bicinchoninic acid (BCA) protein assay kit (Thermo Scientific). SDS-PAGE was carried out with 30 μg protein samples, blotted to a nitrocellulose membrane (Bio-Rad). Membranes were blocked in Tris-buffered saline (TBS, 150 mM NaCl, 25 mM Tris, pH 7.4) containing 5% non-fat dry milk powder (Sigma) and incubated overnight at 4°C with mouse anti-human fibronectin [FBN11] (Thermo Scientific; dilution 1:1,000), mouse anti-human EDA+-fibronectin [IST-9] (Abcam plc; dilution: 1:1,000), and rabbit anti-human actin (Thermo Scientific, dilution: 1:1,000) primary antibodies. Membranes were incubated with horseradish peroxidase-conjugated anti-mouse or anti-rabbit secondary antibodies (Southern Biotech; dilution: 1:2,000) for 60 min at room temperature, and the bands were visualized using Pierce™ ECL Western Blotting Substrate (Thermo Scientific). Digital images were taken by C-DiGit Blot scanner (LI-COR Biotechnology), and Image Studio Digits software (LI-COR Biotechnology) was used to analyze the results.
3 RESULTS 3.1 Gene expression profile of dedifferentiated melanocytes is markedly different from mature melanocytesWe performed high throughput mRNA sequencing analysis on cultured dedifferentiated melanocytes (n = 3) that we previously characterized as bipolar, unpigmented, highly proliferative c-Kit and TRP-1 negative, and nestin, EGFR positive cells (Kormos et al., 2011) and on conventionally cultured pigment producing primary melanocytes (n = 2). Comparison of the expression data showed a markedly different gene expression profile in each group between dedifferentiated melanocytes compared to mature melanocytes (Figure 1a). Among 26 genes (ADAMTS20, ADAMTS9, BCL2, CITED1, EDN3, EDNRB, ENPP1, GLI3, GNA11, HPS4, HPS6, KIT, KITLG, LRMDA, MEF2C, MITF, MREG, MYO5A, OCA2, OR51E2, RAB27A, SLC24A5, SOX10, TYRP1, USP13, ZEB2) annotated to the melanocyte differentiation pathway (https://www.gsea-msigdb.org/gsea/msigdb/cards), 15 showed significant change, suggesting that the activation of this pathway is crucial in differentiating between dedifferentiated melanocytes and mature cells (Fisher's exact test, p < .00001). Out of these 15, 13 significantly decreased (TYRP1, EDNRB, OCA2, SOX10, CITED1, LRMDA, SLC24A5, MREG, RAB27A, HPS4, BCL2, MITF, KIT) (Figure 1b) and two (ENPP1, KITLG) significantly increased in dedifferentiated cells.
 The gene expression profile of dedifferentiated and mature melanocytes. A heat map was created based on all the significant changes in gene expression in dedifferentiated (n = 3: D1, D2, and D3) and in mature (n = 2: M1 and M2) cell cultures (a). Genes associated with melanocyte differentiation were also compared (b) and the results showed that with the exception of two genes, ENPP1 that encodes the ectonucleotide pyrophosphatase/phosphodiesterase 1 protein and KITLG which encodes the KIT ligand, every melanocyte differentiation gene was significantly downregulated in the dedifferentiated cells. Analysis was performed using StrandNGS software (www.strand-ngs.com), and moderated t test with Benjamini-Hochberg FDR for multiple testing correction was applied. Fisher's exact test was used to see that genes related to melanocyte differentiation had a significant change out of all gene expression changes (p
3.2 The expression of microphthalmia-associated transcription factor (MITF) and transcription factor SOX10 (SOX10) is not detectable in dedifferentiated melanocytes
The gene expression profile of dedifferentiated and mature melanocytes. A heat map was created based on all the significant changes in gene expression in dedifferentiated (n = 3: D1, D2, and D3) and in mature (n = 2: M1 and M2) cell cultures (a). Genes associated with melanocyte differentiation were also compared (b) and the results showed that with the exception of two genes, ENPP1 that encodes the ectonucleotide pyrophosphatase/phosphodiesterase 1 protein and KITLG which encodes the KIT ligand, every melanocyte differentiation gene was significantly downregulated in the dedifferentiated cells. Analysis was performed using StrandNGS software (www.strand-ngs.com), and moderated t test with Benjamini-Hochberg FDR for multiple testing correction was applied. Fisher's exact test was used to see that genes related to melanocyte differentiation had a significant change out of all gene expression changes (p
3.2 The expression of microphthalmia-associated transcription factor (MITF) and transcription factor SOX10 (SOX10) is not detectable in dedifferentiated melanocytes
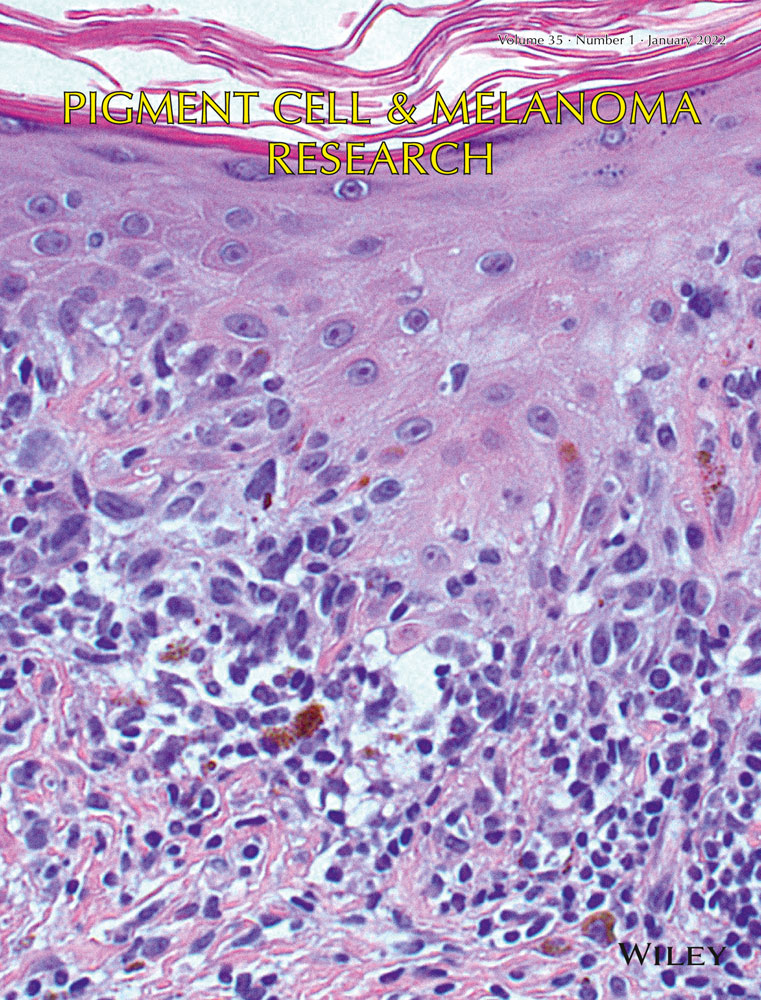
Melanocyte differentiation is mainly regulated by the MITF-SOX10 signaling pathway (Mort et al., 2015). MITF and SOX10 transcription factors are necessary to maintain differentiation and melanin synthesis (Widlund & Fisher, 2003). On immunostaining, the dedifferentiated melanocytes showed no detectable MITF and SOX10 protein expression, while the differentiated cells expressed both (Figure 2a,b, a representative staining of n = 3). This was in line with RNAseq results, which showed a considerable reduction in both MITF and SOX10 transcripts in the dedifferentiated cells compared to the differentiated, mature melanocytes (Figure 1b).

Both Sox10 and MITF transcription factor expression disappears in melanocytes that undergo dedifferentiation. Expression of transcription factor Sox10 (SOX10) (a) is reduced and not detectable in dedifferentiated melanocytes compared to mature cells, and the same change applies to microphthalmia-associated transcription factor (MITF) (b). (n = 3, Original magnification: 40x, scale bar = 20 µm)
3.3 Mesenchymal stem cell-like characteristics of the dedifferentiated melanocytesTo see whether an epithelial to mesenchymal transition (EMT) occurred in the dedifferentiated cells, we used cell surface antigenic profiling. Flow cytometric analysis was performed using an array of markers, proposed to characterize mesenchymal stem cells (MSC) (Dominici et al., 2006). We found that mature and dedifferentiated melanocytes were negative for CD34 and HLA-DR, similar to MSC (Figure 3b). CD105 and CD146 expression was negligible in both types of melanocytes. The expression of CD73 and CD166 and CD44 increased among the dedifferentiated cells, but the most striking difference between mature and dedifferentiated cells occurred in the expression of CD90 (Figure 3a). Mature cells did not express CD90, while the majority of the dedifferentiated melanocytes were CD90 positive. Since CD90 can regulate cell migration (Schubert et al., 2013), in vitro scratching assay was applied to examine the migration of dedifferentiated melanocytes. Compared to mature cells (Figure 4b), the migration of dedifferentiated cells was increased (Figure 4a). We assume the enhanced migration capacity is at least partially related to the increased CD90 expression of the dedifferentiated cells.

Dedifferentiated melanocytes show some characteristics of mesenchymal cells. Comparison between mature and dedifferentiated melanocytes (n = 3) based on cell surface markers showed the highest difference in cluster of differentiation (CD) 90/Thy-1 expression. Representative histograms show the difference in CD90 expression using the FlowJo™ software (BD Biosciences) (a). Cell surface markers required for the definition of mesenchymal stem cells were examined with flow cytometry (b). With the exception of CD90, mature and dedifferentiated cells showed qualitatively somewhat similar expression profile, although there were quantitative differences in the expression. CD29 and CD73 is expressed by the cells in both cultures, and CD199 and CD44 is also expressed by a subpopulation in both types of cultures. CD105 which is expressed by mesenchymal stem cells was not expressed by melanocytes. Similar to mesenchymal cells CD54, CD34 and HLA-DR was not expressed by both differentiated and mature melanocytes. Figure shows mean % of positive cells ± standard error of the mean. Two-tailed Student t test was performed for statistical analysis (n = 3, **p < .005). The differentiation potential of dedifferentiated melanocytes is similar to mesenchymal stem cells (c) Nile Red staining was used to label lipid-laden particles, and 4′,6-diamidino-2-phenylindole (DAPI) was used to stain the nucleus (a). To determine the diffused calcium deposits by the end of the osteogenic differentiation, Alizarin Red S was applied (b). The chondrogenic mass was labeled by Alcian blue–van Gieson staining (c). (n = 1, original magnification: 40×, scale bar = 20 µm.) In 3 different cultures, the karyogram showed no chromosome alterations in the dedifferentiated cells compared to mature melanocytes (a representative karyogram is included, d)

Increased migration rate of the dedifferentieted melanocytes. The migration capacity of dedifferentiated (a) and mature melanocytes (b) with or without mitomycin C treatment were compared using in vitro scratching assay (n = 3, original magnification: 10×, scale bar = 100 µm). Dotted lines indicate the free area. Monitoring was performed by light microscopy. Dedifferentiated melanocytes show enhanced migration capacity compared to mature melanocytes
To test the differentiation capacity of dedifferentiated melanocytes, we used specific differentiation culture conditions (Figure 3c). Trans-differentiation toward the chondrogenic, adipogenic, and osteogenic (Figure 3d) lineages were successful, indicating similarities to MSC.
The karyogram of dedifferentiated melanocytes showed no chromosomal abnormalities (Figure 3d). We also performed soft agar assays where dedifferentiated melanocytes did not form colonies (data not shown).
3.4 The largest relative normalized expression changes in the dedifferentiated cells compared to mature melanocytesThe ten largest negative and positive fold changes (Table 1) show that the genes involved in melanocyte differentiation (TYRP1, TYR, SOX10, GPR143, PMEL, OCA2) are downregulated in the dedifferentiated cells. Furthermore, keratins (KRT14, KRT6A, and KRT5) and P-cadherin (CDH3) are also among the genes that possess the largest negative normalized fold change. The most upregulated gene in the dedifferentiated cells was FGF7 (KGF, keratinocyte growth factor), followed by TBX15. TBX15 plays a crucial role in the development of the musculoskeletal system, and it is associated with tumor suppressor functions such as apoptosis and inhibition of cell growth (He et al., 2002). PRR2C2, GREM2, SHISA9, and RFX8 have not been associated so far with melanocyte development or melanoma; however, ITGBL1 is known to be contained in extracellular vesicles that promote distal metastasis formation through fibroblast-niche formation (Ji et al., 2020). SULF1 was included in a 200-gene collection that is proposed as a gene signature in melanoma (Liu et al., 2013). ZEB1 is a transcription factor that plays role in melanocyte differentiation. ZEB2 MITF and ZEB1 create a transcription factor network that is crucial in melanocyte biology. In differentiated melanocytes from healthy, adult, human donors, ZEB1 could not be detected whereas ZEB2 and MITF were expressed. ZEB1−, ZEB2+, and MITF+ melanocytes are considered differentiated, mature melanocytes (Denecker et al., 2014). ADAM33 is a member of the gene family of disintegrins and metalloproteases (ADAMs). ADAMs are involved in several processes (physiological and pathological) in the skin from which ADAM33 is known to have effects in dermal immunology and inflammation (Kawaguchi & Hearing, 2011).
TABLE 1. The top 10 genes exhibiting the largest positive and negative fold change (FC) in expression in dedifferentiated melanocytes compared to mature melanocytes Largest positive fold change (FC) Largest negative fold change (FC) Gene symbol FC Gene symbol FC FGF7 568.57 TYRP1 −22500.035 TBX15 408.58 TYR −14825.283 RP11−334J6.7 371.95 KRT14 −14467.146 ITGBL1 341.56 KRT6A −11465.412 GREM2 341.22 SOX10 −10182.477 SHISA9 315.29 GPR143 −10102.731 SULF1 292.41 PMEL −9607.309 ZEB1 282.90 KRT5 −8736.197 RFX8 267.12 CDH3 −8225.915 ADAM33 259.42 OCA2 −7834.8706 3.5 Ingenuity Pathway Analysis reveals that the most upregulated genes converge in the FGF2 signaling pathwayBased on the most upregulated genes, IPA analysis revealed that FGF2 signaling is the most affected in the dedifferentiated cells. Several downstream and upstream regulators were found which were linked to cellular processes involving cell migration, cytoskeletal organization, and mechanisms involved in angiogenesis such as endothelial cell movement, endothelial cell migration, development of vasculature, growth of blood vessels, muscle cell migration, and movement (Figure 5).

IPA analysis shows FGF2 in the center of signaling pathways of the most upregulated genes in dedifferentiated melanocytes. Graphical summary of the regulatory Ingenuity Pathway Analysis (IPA). Based on the most upregulated genes, the affected signaling pathways merge around fibroblast growth factor 2 (FGF2) in dedifferentiated melanocytes. The dominant regulatory processes that are involved in the dedifferentiation are cytoskeletal and cytoplasmic organization, cell movement, cell migration, angiogenesis, and hepatic fibrosis. Cell specificity of these pathways shows the involvement of endothelial cells, and vascular and smooth muscle cells, indicating the central importance of angiogenesis and vasculogenesis. (Data analysis was performed on RNAseq data of dedifferentiated (n = 3) and mature (n = 2) melanocytes)
3.6 Expression of E-cadherin is lost in dedifferentiated melanocytes, while fibronectin (FN) and EDA containing fibronectin (EDA+FN) expression increasedDifferentiated cells show a strong staining for the E-cadherin expression (Figure 6a), while dedifferentiated cells show no visible staining (Figure 6a) indicating that cells lost E-cadherin expression. At the same time, increased expression is detected for FN and EDA+FN using Western blot (Figure 6b).

E-cadherin expression is reduced, while EDA+FN and FN expression levels are elevated in dedifferentiated melanocytes. E-cadherin, oncofetal fibronectin (EDA+FN), and fibronectin (FN) are important markers in dedifferentiation and epithelial–mesenchymal transition (EMT). (a) In dedifferentiated melanocytes, the expression level of E-cadherin was reduced compared to differentiated, mature melanocytes (n = 3). (b) The opposite change can be observed regarding EDA+FN and FN. Dedifferentiated melanocytes increased the synthesis of EDA+FN and FN, whereas in mature melanocytes their expression was reduced (n = 1)
3.7 The protein kinase C pathway is altered in dedifferentiated melanocytes compared to mature cellsOf the genes related to the protein kinase C pathway (Biocarta PKC Pathway), significant changes in gene expression occurred in PRKCB, which encodes the protein kinase C β (PKC-β); PRKCA, which encodes the protein kinase C α (PKC-α) protein; in the PLCB1 gene encoding the 1-phosphatidylinositol-4,5-bisphosphate phosphodiesterase beta-1 enzyme; and in NFKB, the gene of the p105 subunit of nuclear factor kappa B (NFKB). Differentiated melanocytes highly expressed PRKCB, while the expression of PRKCA, PLCB1, and NFKB1 was lower. In the dedifferentiated cells, PRKCB expression decreased considerably, while PRKCA and PLCB1 increased. Change in NFKB1 gene expression was not as strong between the two cell types (Figure 7).

Gene changes in the protein kinase C signaling pathway in dedifferentiated melanocytes. The expression of the protein kinase C β gene (PRKCB) is significantly decreased, while the expression of protein kinase C α gene (PRKCA) and 1-phosphatidylinositol-4,5-bisphosphate phosphodiesterase beta-1 gene (PLCB1) is significantly increased in dedifferentiated cells. The expression of nuclear factor NF-kappa-B p105 subunit gene (NFKB1) was elevated in dedifferentiated cells, but not significantly compared to matured cells. Figure shows the average gene expression values ± standard error of the mean (SEM). Two-tailed Student t test was performed for statistical analysis (n = 2 mature, n = 3 dedifferentiated cells, *p < .05, **p < .005)
4 DISCUSSIONIt is increasingly clear that cellular differentiation is not unidirectional and permanent. Many terminally differentiated cells retain the capacity to dedifferentiate into stem cell or transdifferentiate into another differentiated cell type due to changes in extracellular signals (Bonfanti et al., 2012). This plasticity depends on environmental or extrinsic factor mediated reprogramming of the cell and is referred to as extrinsic plasticity, as opposed to the intrinsic plasticity that is induced by forced expression of transcription factors (Gupta et al., 2019). In vitro cell culturing can result in phenotypic changes of the cells, and dedifferentiation and transdifferentiation may occur depending on the culture environment. Extrinsic reprogramming of cells has significant implications for regenerative medicine and cancer therapy.
Previously, we described a novel in vitro culture technique for human melanocytes derived from interfollicular adult epidermis in which cells dedifferentiated. In our medium, which did not contain specific mitogens, the cells gradually lost pigmentation, TRP1 and c-Kit expression became bipolar and highly proliferative, their life span increased, and they expressed EGFR and nestin. The dedifferentiation of the cells was reversible with the addition of specific mitogens (CT and PMA) or UVB irradiated keratinocyte supernatants. As expected and proved in this work by RNAseq and immunohistochemical staining MITF, a crucial regulator of melanocyte differentiation was also lost in the dedifferentiated cells.
Melanocytes develop from the neural crest in vertebrate embryos. During development, neural crest cells undergo an epithelial to mesenchymal transition, migrate, and at specific locations differentiate into diverse cell types, peripheral neurons, Schwann cells, osteocytes, chondrocytes, adipocytes, smooth muscle cells, melanocytes, and keratinocytes. Increasing evidence suggest that many differentiated cells regain the ability to undergo dedifferentiation due to extrinsic reprogramming. In vitro cell cultures can serve as model systems to investigate cellular dedifferentiation. Understanding cell plasticity is relevant to tissue regeneration, wound healing and importantly to cancer initiation, progression, and therapy resistance. Stem cell-derived models of neural crest and melanoblast cultures have been used to understand melanoma progression (Bosserhoff et al., 2011).
Melanoma is one of the deadliest human cancers. Besides genetic changes resulting in tumor cell heterogeneity, environmentally regulated epigenetic reprogramming seems to be an important driver of disease progression (Rambow et al., 2019). Switching phenotype through dedifferentiation is well described in melanoma, and it is known that melanoma dedifferentiation is closely related to the development of metastasis, therapy resistance, and long-lived dormant cancer stem cell development. Based on the relative SOX10 and MITF expression, six different melanoma phenotypic states have been described and each specific phenotype has been assigned to functions of proliferation and migration. These six melanoma phenotypic states are characterized as MITFhigh differentiated cells (hyper-differentiated state), MITF positive proliferating cells (melanocytic state), intermediate cell state consisting of migrating neural crest melanoblasts, therapy-induced starved melanoma cells (SMC) state, MITFlow neural crest stem cell (NCSC), and the MITF negative undifferentiated state. SOX10 expression distinguishes between Sox10 positive NCSC and Sox10 negative invasive/mesenchymal SMC. Data indicate that undifferentiated, NCSC and SMC state cells exhibit both proliferative and invasive features. The complete loss of MITF and Sox10 characterizes the most undifferentiated melanoma cells. Tsoi et al. categorized melanoma differentiation as four distinct stepwise changes using consensus hierarchal clustering of expression profiles from a panel of 53 human melanoma cell lines (Tsoi et al., 2018). Melanocytic, transitory neural crest-like cells all expressed Sox10, and loss of Sox10 expression was characteristic for cells in the undifferentiated cluster. Since dedifferentiation of melanoma is induced by targeted therapy and during immunotherapy in response to inflammatory signaling, inhibition of dedifferentiation is of great importance in preventing therapy resistance.
While dedifferentiation of melanoma has been extensively studied, much less is known about the dedifferentiation of human melanocytes. While most human melanoblast cultures use foreskin melanocytes (Cook et al., 2003), the advantage of our culture is the use of adult skin derived cells. The lack of chemical mitogen in the culture also makes our system unique. Based on the lack of both MITF and Sox10 expression, these dedifferentiated melanocytes resemble undifferentiated melanoma cells. While TYRP1, EDNRB, OCA2, SOX10, CITED1, LRMDA, SLC24A5, MREG, RAB27A, HPS4, BCL2, MITF, and KIT, all well-characterized melanocyte differentiation genes, significantly decreased in the cells, KITLG and ENPP1 showed a significant increase. KITLG plays an essential role in stem cell maintenance, in hematopoiesis, gametogenesis, mast cell development, and in melanogenesis. Meta analyses of studies comparing GWAS data in nevus and melanoma indicate that KITL has no effect on melanoma risk among nevus genes (Duffy et al., 2018). ENPP1 encodes the ectonucleotide pyrophosphatase/phosphodiesterase 1, a transmembrane enzyme, important in ATP to AMP hydrolysis. By generating inorganic pyrophosphate, it regulates tissue calcification and bone development. In tumors, ENPP1 can promote metastasis by degrading extracellular cGAMP, an intracellular second messenger that activates the innate immune STING pathway in response to cytosolic double-stranded DNA (Ritchie et al., 2019). Increased ENPP1 expression in human tumors correlates with suppressed anti-tumor immunity and resistance to PD1 inhibitor therapy (Li et al., 2020). ENPP1 is highly expressed in stem-like tumor cells in Grade IV glioma and has been shown to be functionally related to cell cycle progression (Bageritz & Goidts, 2014). ENPP1 knockdown decreases the transcriptional activity of E2F transcription factor 1 resulting in accumulation of cells in G1 phase.
The dedifferentiated melanocytes did not express CD34 and HLA-DR, similar to mesenchymal stem cells; however, the expression of CD105 and CD146 that are considered mesenchymal cell markers was negligible in the cells. The expression of CD73 and CD166 and CD44 increased among the dedifferentiated cells, but the most striking difference between mature and dedifferentiated cells occurred in the expression of CD90. CD90/Thy-1 is a member of the immunoglobulin superfamily, a cell surface glycoprotein, originally described in mouse T cells. It plays a role in cell–cell, cell–matrix interaction through binding to integrins αv/β3, αx β 2, syndecan-4, CD90 itself, and CD97 (Sauzay et al., 2019). It is considered a cancer stem cell marker, and it regulates tumor cell migration and metastasis. It is strongly expressed in melanoma in blood vessel and lymphatic endothelial cells (Schubert et al., 2013). In a recently published article, Thy-1 was used as a marker to detect cancer associated fibroblasts (CAF) before and after immunotherapy (Wong et al., 2019). It is not generally described in melanoma cells, although Thy-1 expression was detected in 12 of 19 primary melanomas and only one of 23 melanoma metastases (Hersey et al., 1983), and in desmoplastic melanoma, a rare melanoma subtype (Winnepenninckx et al., 2003).
Beside characteristic expression of cell surface molecules, mesenchymal stem cells are defined by their ability to adhere to plastic, by their morphology being fibroblast-like, and by their differentiation potential toward osteoblast, chondrocytes, and adipocytes (Gebler et al., 2012). Our dedifferentiated melanocytes fulfill these criteria, indicating the potential to transdifferentiate toward mesenchymal stem cells. Epithelial–mesenchymal transition (EMT) is considered a crucial step in tumor progression. It is closely linked to induction of stem cells, and the possible association between EMT and stem cell formation is not entirely clear (Mani et al., 2008). In melanoma, it is not clear whether melanoma cells would dedifferentiate first into a highly undifferentiated state and subsequently adopt an alternative cell fate (Rambow et al., 2019). Evidence suggest that in tumors, a full mesenchymal transition is not required for tumor cell metastasis, and hybrid epithelial/mesenchymal populations exist that contribute to higher metastatic potential (Pastushenko et al., 2018). Although the cell population of our dedifferentiated cell culture is not entirely homogenous, dedifferentiated melanocytes in our culture indicate a partial mesenchymal transition. In fact, it has been proposed that partial EMT in cancer cells could result in a more tumorigenic and chemoresistant phenotype compared to cancer cells that have predominant mesenchymal or epithelial features (Jolly et al., 2019).
Loss of E-cadherin in cancer cells leads to activation of EMT and to metastatic dissemination (Onder et al., 2008). ZEB1 is one of the transcriptional pathways activated in E-cadherin loss, and ZEB1 is among the top ten genes showing the largest positive fold change in dedifferentiated melanocytes compared to differentiated cells. Our dedifferentiated melanocytes lost E-cadherin expression, and at the same time, both fibronectin (FN) and its splice variant, EDA+ FN (also known as oncofetal fibronectin) showed increased expression in the cells. Loss of E-cadherin inhibits the activity of tumor infiltrating CD8+ CD103+ T cells and reduces checkpoint inhibitor effectiveness in melanoma (Shields et al., 2019). Increased expression of EDA+ FN is detected in metastatic melanoma (Natali et al., 1995).
It is somewhat surprising that we detected the largest fold change increase in FGF7 (fibroblast growth factor7/KGF, keratinocyte growth factor) in the dedifferentiated melanocytes. So far, FGF7/KGF expression has not been described in melanocytes and there are little data available on its role in melanocyte biology. It has been proposed that FGF7/KGF promotes melanosome transfer acting on keratinocytes by stimulating phagocytosis (Cardinali et al., 2008) and that it may play a role in solar lentigo formation (Kovacs et al., 2010). In previous work, we proposed that FGF7/KGF could play a role in the splicing regulation of EDA+FN in fibroblasts. We also investigated the mRNA expression of the different splice variants of fibroblast growth factor receptor (FGFR2) in keratinocytes, fibroblasts, and melanocytes. Human fibroblasts in culture expressed the FGFR2-IIIc, and keratinocytes and melanocytes expressed the FGFR2-IIIb splice variant (Gubán et al., 2016). Further work is needed to better understand the role of FGF7/KGF in melanocyte biology.
Ingenuity pathway analysis of the RNAseq data revealed that the top pathways that changed during dedifferentiation merged around FGF2. This is not surprising, since the family of FGF and FGFR has a crucial role in embryonic development. They have an important regulatory function in various cancers, including melanoma (Czyz, 2019
留言 (0)