
記住我
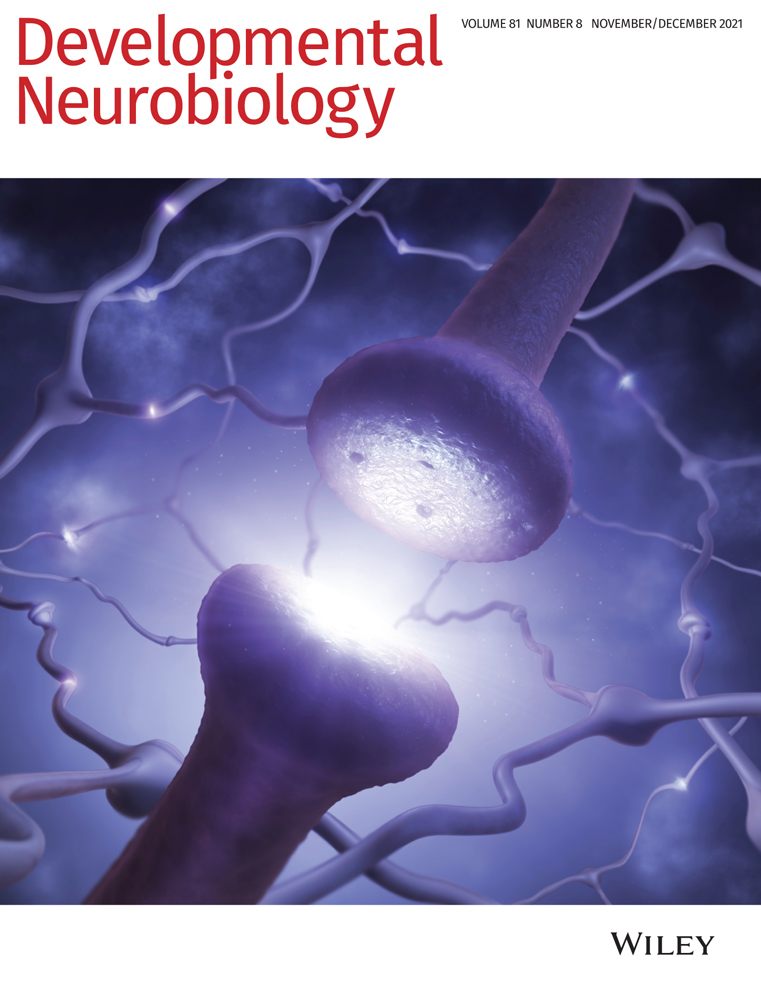
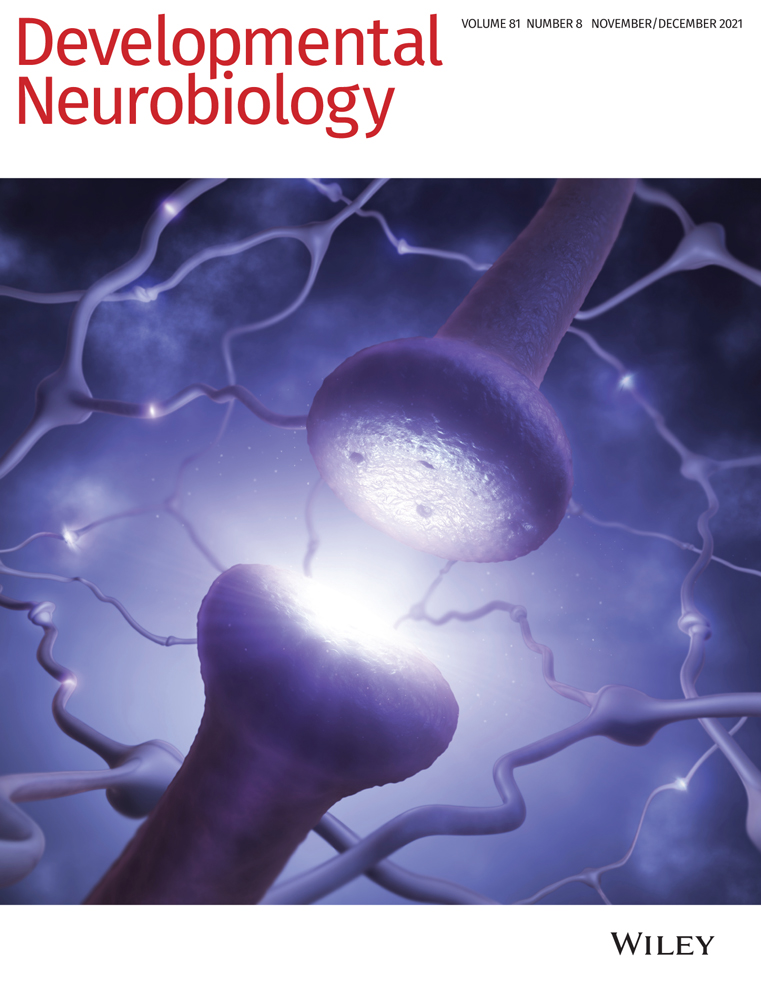
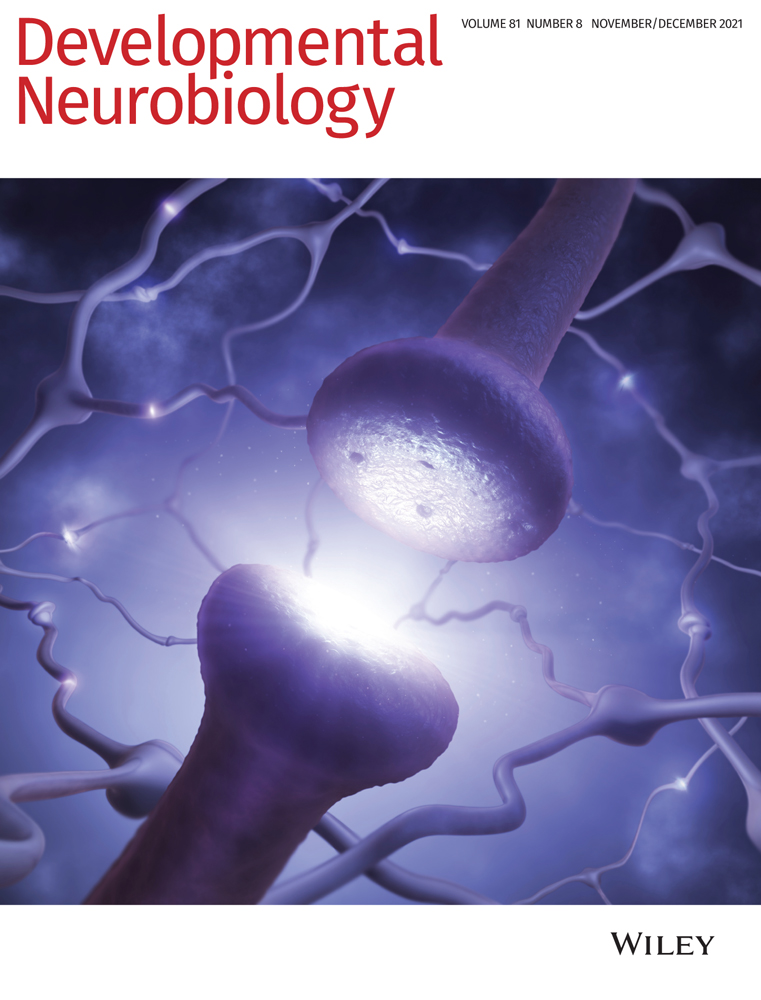
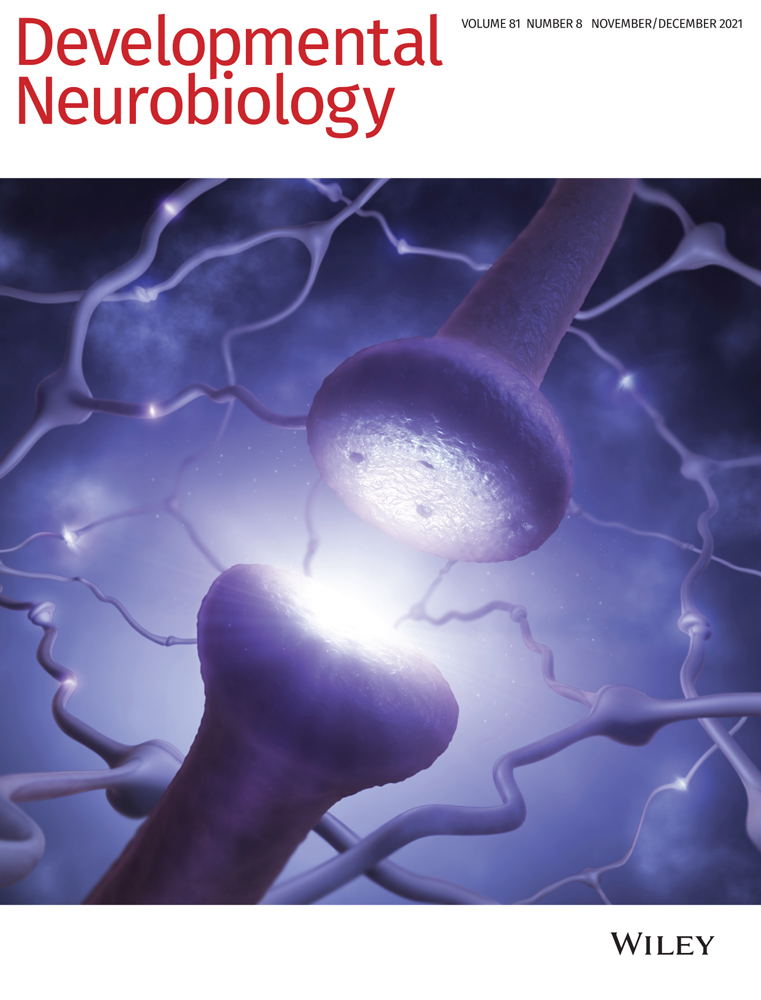
First metazoan animals populated the earth about 650 million years ago with the appearance of demospongiae (Love et al., 2009). Shortly thereafter, animals with nervous systems and muscles appeared that were able to sense and process external and internal stimuli to adjust their behavior. It is well established that neurons connect themselves in amazingly complex circuits which compute all information. To do so, neurons are able to change their membrane potential very fast which enables information transport by action potentials. The generation of action potentials depends on the expression of voltage-gated sodium and potassium channels and finally results in the release of neurotransmitters. Interestingly, the molecular machinery underlying all these basic neuronal properties are well conserved throughout evolution (Moran, Barzilai, Liebeskind, & Zakon, 2015; Senatore, Raiss, & Le, 2016; Varoqueaux & Fasshauer, 2017; Walter, Böhme, & Sigrist, 2018). Despite the undisputed importance of neurons, it is important to note that the nervous system also harbors glial cells. Glial cells are not able to change their membrane potential as fast as neurons can do, but closely interact with them and in some parts of the mammalian brain even outnumber neurons (Herculano-Houzel, 2014).
As mentioned above, neurons all use the same molecular repertoire to establish their intricate communication system. It might therefore be expected that glial cells also employ evolutionarily conserved molecular processes to exert their functions. However, vertebrate glial cells often do not have clear invertebrate counterparts. For example, neural crest-derived, myelinating Schwann cells in the peripheral nervous system (PNS) or centrally located myelinating oligodendrocytes are not known in invertebrates. Likewise, invertebrates typically have a glial blood–brain barrier whereas higher vertebrates establish an endothelial blood–brain barrier. On the other side, some aspects of glial differentiation appear conserved between invertebrates and vertebrates supporting a common evolutionary origin. To better address whether glial cells originated only once during evolution or evolved several times we first need to understand neuron–glia interaction in several model organisms before we can search for common themes and signatures.
Drosophila is a well-characterized invertebrate model system. It has a low number of very well characterized glial cells, amenable for sophisticated genetic and molecular manipulation. Due to the availability of large sets of characterized promoter elements that either drive Gal4, LexA, Flp, or Cre in specific glial subsets amazing manipulations with unprecedented spatial and temporal resolution can be conducted (Li et al., 2014). In the following, we will briefly compare the different Drosophila glial cells with those found in vertebrates to then focus on the principles underlying neuron–glia interaction in the fly nervous system.
2 DROSOPHILA GLIAL CELLSTo start, we first need to define what is glia. This is in particular relevant for the invertebrate nervous system, since it harbors three distinct ectodermaly derived cell types: Neurons, tracheal and glial cells. Neurons are electrically excitable and establish the neuronal circuits. All differentiated neurons express neuronal synaptobrevin. Tracheal cells invade the entire nervous system and establish a tubular system to provide the brain with oxygen. All tracheal cells express the FGF receptor Breathless (Klämbt, Glazer, & Shilo, 1992). The remaining cells are glial cells. To date there is no molecular marker that is expressed by all glial cells. The most specifically expressed protein is the transcription factor Repo (Xiong, Okano, Patel, Blendy, & Montell, 1994), which is found in all but the midline glial cells (Halter et al., 1995; Xiong et al., 1994). Therefore, Repo expression is generally used as marker of glial fate.
In Drosophila, glial cells come in a surprisingly large cell-type diversity, although their relative number is rather low (Yildirim, Petri, Kottmeier, & Klämbt, 2018) (Figure 1). The body plan of Drosophila larvae is segmentally organized, with three thoracic and eight abdominal segments. At the end of embryogenesis, each of these segmental units contains about 700 neurons and 77 glial cells. About 50 of these neurons are peripheral sensory neurons that project their axons through segmentally arranged nerves to the ventral nerve cord, which can be considered analogous to the spinal cord in vertebrates (Arendt & Nübler-Jung, 1999; Arendt, Tosches, & Marlow, 2016; Singhania & Grueber, 2014). At the end of embryogenesis, 12 glial cells accompany each abdominal nerve, whereas 65 glial cells reside in the segmentally organized ventral nerve cord (Beckervordersandforth, Rickert, Altenhein, & Technau, 2008; von Hilchen, Beckervordersandforth, Rickert, Technau, & Altenhein, 2008). The embryonic brain originating from the head segments is considerably more complex with about 10,000 cells total and will not be considered further here. The Drosophila nervous system (larval and adult central nervous system [CNS] and PNS) comprises seven morphologically defined glial cell types which show a surprisingly high molecular diversity (Ito, Urban, & Technau, 1995; Klämbt & Goodman, 1991). Four of these glial cell types are found only in the CNS (astrocyte-like glia, ensheathing glia, midline glia, and cortex glia), one cell type is found only in the PNS (wrapping glia), and two cell types are found in both the CNS and the PNS (perineurial glia and subperineurial glia, Figure 1).

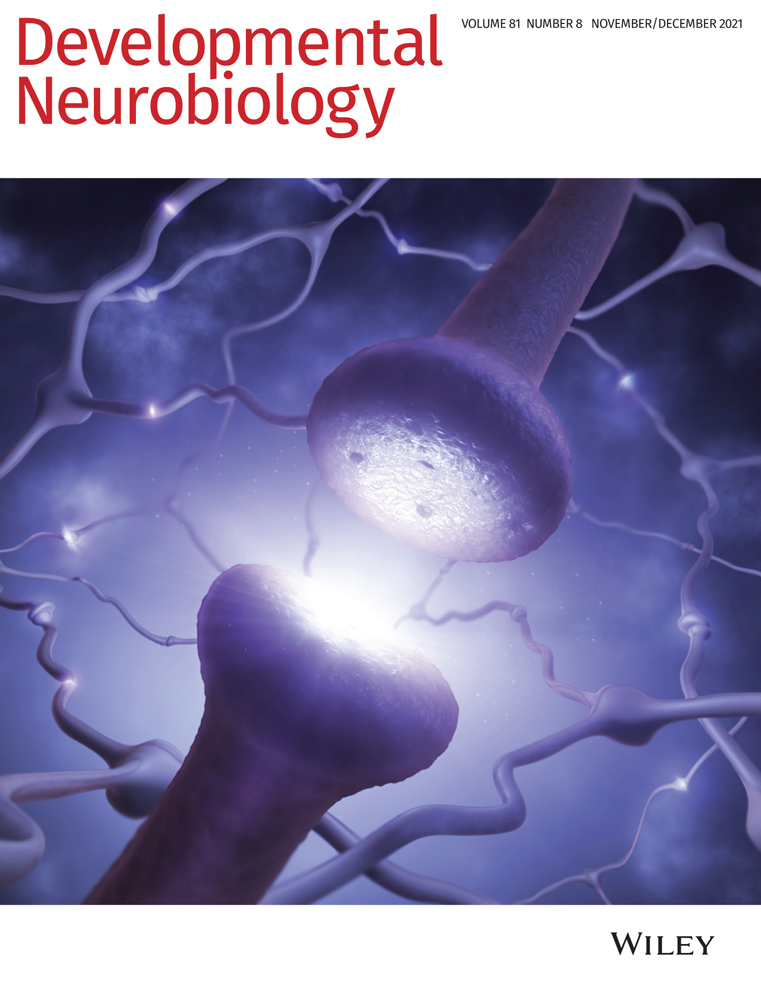
Schematic representation of neuron–glia interaction in the Drosophila nervous system. (a) Schematic view on a third instar larval brain. The relevant glial cell types are indicated and shown in the same color code throughout the figure. The ventral nerve cord (vnc) comprises three thoracic (t1–t3) and eight abdominal neuromers (a1–a8). Axon tracts are shown in dark gray, cortex regions with neuronal and glial cell nuclei in gray, synaptic neuropil in light gray. The dotted line indicates the level of the orthogonal view shown in (c). (b) Schematic view on an adult fly brain. Note that the thoracic neuromers increased in size, and the segmental nerves from the abdominal segments have fused, (c) Schematic cross section through the ventral nerve cord. Color coded glial cells are: ALG, astrocyte-like glia; CG, cortex glia; EG, ensheathing glia; MG, midline glia; PG, perineurial glia; SPG, subperineurial glia. The peripheral wrapping glia (WG) covers peripheral axons. The SPG are connected by septate junctions (SJ). The entire nervous system is covered by an extracellular matrix called neural lamella (NL). All neuron–glia contact sites found in the larval or the adult nervous system are indicated by a red line
2.1 Astrocyte-like glial cellsThe three CNS specific glial cell types are cortex glia and two neuropil-associated glial cell types: ensheathing glia and astrocyte-like glial cells (Figure 1). Whereas in vertebrates astrocytes are the most frequent glial cell type, the Drosophila CNS harbors only a small number of these cells, with only six astrocyte-like glial cells found in each larval hemineuromere (one half of a segmental neuromere), which tile the synaptic neuropil in a characteristic manner (Stork, Sheehan, Tasdemir-Yilmaz, & Freeman, 2014). During adult stages, the number of astrocyte-like glial cells increases dramatically and about 4,600 are found in the adult brain. Although they vary in size and shape, they retain all astrocyte-like characteristics (e.g., processes and tiling behavior) (Kremer, Jung, Batelli, Rubin, & Gaul, 2017). As their vertebrate counterparts, Drosophila astrocyte-like glial cells are coupled by gap junctions and form many protrusions that invade into the neuropil to control neurotransmitter homeostasis (Ma, Stork, Bergles, & Freeman, 2016; MacNamee et al., 2016; Muthukumar, Stork, & Freeman, 2014; Peco et al., 2016; Stork et al., 2014). The elaboration of the intricate branching pattern in the larval ventral nerve cord depends on FGF-signaling. In the developing visual centers the transmembrane leucine-rich repeat (LRR) protein Lapsyn works in concert with the FGF-signaling to promote branch morphogenesis (Richier, Vijandi, Mackensen, & Salecker, 2017; Stork et al., 2014). Interestingly, and in contrast to vertebrates, astrocytic processes are often found at quite some distance of synapses and thus tripartite synapses as found in the vertebrate CNS may not exist in Drosophila (MacNamee et al., 2016; Stork et al., 2014). At the example of a defined neuron (looper neuron) 3D reconstructions of 67 synapses in three abdominal segments of a third instar larva showed that the mean distance of astrocytic processes from the synapse is 375 nm (MacNamee et al., 2016). Likewise, additional TEM studies during pupal stages did not reveal clear evidence for astrocyte-like glial cells ensheathing synapses (Muthukumar et al., 2014). Indeed, ablation of astrocyte-like glial cells resulted in a normal appearing synapse morphology but resulted in reduced numbers of synapses, indicating that astrocyte-like glial cells are involved in the modulation of synaptic plasticity (Muthukumar et al., 2014). However, during metamorphosis, astrocyte-like glial cells can activate Draper (Drpr) expression in an ecdysone-dependent manner to engulf and clear neuronal debris (Tasdemir-Yilmaz & Freeman, 2014). In conclusion, the concept of tripartite synapses remains to be demonstrated for the Drosophila CNS, while in the PNS tripartite synapses have been suggested (Danjo, Kawasaki, & Ordway, 2011; Strauss, Kawasaki, & Ordway, 2015).
To guarantee neurotransmitter homeostasis Drosophila astrocyte-like glial cells express a set of specific transporter proteins such as the excitatory amino acid transporter 1 (EAAT1) or the GABA transporter (GAT) (Muthukumar et al., 2014; Soustelle, Besson, Rival, & Birman, 2002; Stacey et al., 2010; Stork et al., 2014). Expression of EAAT1 requires Notch-mediated neuron–glia interaction (Stacey et al., 2010) and focal adhesion signaling (Cho, Muthukumar, Stork, Coutinho-Budd, & Freeman, 2018). Due to the function of EAAT1 as a chloride channel, astrocyte-like glial cells are also important players in providing ionic homeostasis in the neuropil (Parinejad, Peco, Ferreira, Stacey, & van Meyel, 2016). Additionally, Drosophila astrocyte-like glial cells express receptors like the Octopamine/tyramine receptor and the transient receptor potential channel Waterwitch (Ma et al., 2016). Forced increase of intracellular Ca2+ concentration in astrocyte-like glial cells triggers rapid endocytosis of the GABA transporter GAT from astrocytic membranes, which leads to paralysis and suppression of neuronal activity (Ma et al., 2016; Zhang, Ormerod, & Littleton, 2017). In the olfactory system, astrocytic Ca2+ signaling can regulate olfactory processing by affecting the synaptic strength between olfactory receptor neurons and downstream projection neurons (Liu et al., 2014). In vertebrates, astrocytic Ca2+ signaling is linked to the release of gliotransmitters which act on neurons and regulate neuronal spiking and synaptic plasticity (Bazargani & Attwell, 2016). However, this connection remains to be clearly demonstrated in Drosophila.
The release of gliotransmitters, or gliotransmission, may be mediated by proteins such as the small, secreted Ig-domain protein Noktochor (Nkt). Glial suppression of Nkt expression in fly astrocyte-like glial cells results in decreased night sleep (Sengupta, Crowe, You, Roberts, & Jackson, 2019). In addition, similar as in vertebrates, cytokine signaling affects sleep behavior. The Drosophila tumor necrosis factor-alpha homologue, Eiger, must be expressed in astrocyte-like glial cells to activate its receptor Wengen expressed by neurons which is required for normal sleep duration (Vanderheyden et al., 2018). Another evolutionarily well conserved pathway promoting sleep is associated with fatty acid metabolism. Fabp7-deficient mice show fragmented sleep (Gerstner et al., 2017). Endogenous Drosophila Fabp expression is found in the cortex glia which are also linked to sleep regulation (Farca Luna, Perier, & Seugnet, 2017; Kis, Barti, Lippai, & Sass, 2015). Expression of a mutant mouse Fabp7 protein specifically in Drosophila astrocyte-like glial cells using the alrm-Gal4 driver also results in fragmented sleep (Gerstner et al., 2017), but it is currently unknown whether astrocyte-like glial cells communicate with neurons directly or indirectly via cortex glia. In addition, astrocyte-like glial cells are linked to circadian rhythmicity of the animal (Ng et al., 2016; Suh & Jackson, 2007; You, Fulga, Van Vactor, & Jackson, 2018). In summary, one can conclude that comparable to vertebrate astrocytes, Drosophila astrocyte-like glial cells are in the center stage to modulate neuronal activity.
2.2 Ensheathing glial cellsThe ensheathing glial cells embrace the entire neuropil to form an internal barrier-like structure of still unknown function (Figure 1). Four ensheathing glial cells are found in every larval hemisegment. Three of these originate from a common lineage together with the six astrocyte-like glial cells found in every hemineuromer (lateral glioblast, Peco et al., 2016). Two ensheathing glial cells encircle the neuropil, whereas a third ensheathing glial cell covers part of the neuropil and wraps axons running in the nerve segments connecting the neuropil with peripheral nerves. A fourth ensheathing glial cell is generated by a different linage (NB1-3, Beckervordersandforth et al., 2008) and also wraps axons projecting toward the peripheral nerve. During metamorphosis, some of the larval ensheathing glial cells undergo cell death while others are able to re-differentiate and proliferate to give rise to the adult neuropil-associated glial cells which are derived from the neuroblast type II lineages (Kremer et al., 2017; Omoto, Yogi, & Hartenstein, 2015; Ren, Awasaki, Wang, Huang, & Lee, 2018).
Ensheathing glia ensure compartmentalization between the neuropil and the cortex region but the question remains why this additional barrier is established. Although ensheathing glial cells do not infiltrate the neuropil, they are able to fulfill neuromodulatory tasks. They express the sulfite oxidase Shopper that detoxifies sulfite to sulfate. High sulfite levels inhibit the function of glutamate dehydrogenase and thereby indirectly affect glutamate homeostasis (Otto et al., 2018). Possibly, ensheathing glia provide a gradient of glutamine to the neuropil which would suggest that positioning of a synapse in the neuropil reflects its general efficacy (Otto et al., 2018). Interestingly, high levels of the mammalian Shopper homologue Suox are expressed in oligodendrocytes (Zhang et al., 2014). Ensheathing glial cells also express transporters like the Excitatory amino acid transporter 2 (EAAT2) that has been shown to transport taurine across cell membranes and has a modulatory role in controlling the sleep–wake rhythm (Besson, Ré, Moulin, & Birman, 2005; Peco et al., 2016; Stahl et al., 2018). Unlike astrocyte-like glial cells, which acquire phagocytic properties during circuit remodeling (Muthukumar et al., 2014; Tasdemir-Yilmaz & Freeman, 2014), ensheathing glia express key components of the glial phagocytic machinery (e.g., the engulfment receptor Drpr, and dCed-6), to react to injury and phagocytose degenerating axons from the injured brain (Doherty, Logan, Taşdemir, & Freeman, 2009; Hilu-Dadia, Hakim-Mishnaevski, Levy-Adam, & Kurant, 2018; Purice et al., 2017).
As stated above, ensheathing glial cells embrace the entire synaptic neuropil. Importantly, axons that run within the neuropil are generally not wrapped (see below). This is a main difference to the vertebrate CNS where oligodendrocytes form myelin sheaths around segments of up to 20 large caliber axons (Simons & Nave, 2016). Therefore, no oligodendrocyte-like cells may exist in Drosophila. The notion that two ensheathing glial cells wrap axons that connect the periphery with the neuropil, however, suggests that axonal insulation by glia is indeed an evolutionarily ancient function.
2.3 Midline gliaDifferent to all other glial cells of the Drosophila nervous system, the midline glial cells are specified by the transcription factor Single minded (Crews, Thomas, & Goodman, 1988; Thomas, Crews, & Goodman, 1988). Early on the midline glial cells are required for the establishment of commissures by secreting evolutionary conserved axon guidance cues such as Netrin and Slit (Brose et al., 1999; Howard, Brown, Wadsworth, & Evans, 2017; Kidd, Bland, & Goodman, 1999; Mitchell et al., 1996; Serafini et al., 1994). In addition to its role in guiding commissural axons, the midline glia exerts a number of signaling functions to control development of lateral neurons similar to the vertebrate floorplate (Danesin & Soula, 2017; Menne, Luer, Technau, & Klämbt, 1997). During larval stages, the midline glial cells perform similar functions as the peripheral ensheathing glia that wrap axons on their way to the CNS. Axonal wrapping requires expression of the Ig-domain protein Wrapper by the midline glia that binds a neuron specific Neurexin IV isoform (Stork et al., 2009; Wheeler et al., 2009). The function of the midline glial cells during pupal and adult stages has not yet been addressed (Stollewerk, Klämbt, & Cantera, 1996). A comprehensive review of the Drosophila midline glia has been recently published (Crews, 2019).
2.4 Cortex gliaThe cortex glia encases all neuronal cell bodies residing in the cortex area of the Drosophila CNS (Figure 1). They engulf the progeny of individual neural stem cells, called neuroblasts, and thus keep neuronal lineages in a common encasement (Dumstrei, Wang, & Hartenstein, 2003; Pereanu, Shy, & Hartenstein, 2005; Spindler, Ortiz, Fung, Takashima, & Hartenstein, 2009). Growth of the cortex glia is regulated in part by autocrine signaling through the neurotrophin Spätzle 3 (Coutinho-Budd, Sheehan, & Freeman, 2017). Cortex glial cells participate in metabolic support of neurons by expressing the lactate/pyruvate transporter Chaski, which is essential for proper synaptic transmission (Delgado et al., 2018). In addition, these glial cells control neuroblast proliferation (Coutinho-Budd et al., 2017; Read, 2018; Spéder & Brand, 2018). The effect on neurogenesis requires the chloride channel ClC-a which is expressed by the cortex glia (Plazaola Sasieta et al., 2019) and possibly the SLC7 amino acid transporter Sobremesa (Sbm) (Galagovsky et al., 2018). Sbm is expressed in cortex glia and the subperineurial glia (see below). Glial knockdown results in reduced brain growth and reduced Ecdysone production demonstrating the potent effect of glial signaling on the entire body (Galagovsky et al., 2018). In addition, cortex glial cells are able to modulate neuronal activity by interacting with neuronal somata. The Na+Ca2+K+ (NCKX) exchanger Zydeco (Zyd) is expressed by the cortex glia where it is required for Ca2+ oscillations in small discrete areas of about 4.4 µm2, called microdomains (Melom & Littleton, 2013). Mutations in zyd cause a seizure-like behavior by increased rates of calcineurin-dependent endocytosis that in turn impairs K+ buffering abilities of the cell by enhanced internalization of the K+ channel sandman (Melom & Littleton, 2013; Weiss, Melom, Ormerod, Zhang, & Littleton, 2019). Normal extension of the cortex glial plasma membrane requires ceramide phosphoethanolamine generated by the enzyme ceramide phosphoethanolamine synthase and is needed to suppress seizures induced by visual stimulation (Kunduri et al., 2018). In addition, cortex glial cells are required for several behavioral traits including sleep, larval motor control and response to ethanol (Coutinho-Budd et al., 2017; Farca Luna et al., 2017; Lee, Mathies, & Grotewiel, 2019). Cortex glia are also able to clear cell bodies of dead neurons via the phagocytosis receptor Drpr (McLaughlin, Perry-Richardson, Coutinho-Budd, & Broihier, 2019; Nakano et al., 2019). Although, no clear vertebrate homolog exists of this cell type, cortex glia share some functional similarities with astrocytes (Melom & Littleton, 2013), exert some microglia-like functions (McLaughlin et al., 2019; Nakano et al., 2019), and even wrap proximal axons on their way to the neuropil similar to oligodendrocytes (Coutinho-Budd et al., 2017; Spindler et al., 2009). Finally, cortex glial cells resemble satellite cells found in the vertebrate PNS (Hanani, 2005).
2.5 Peripheral glial cellsThe PNS of Drosophila is organized as the vertebrate PNS. Sensory neurons detect environmental stimuli and transmit the information into the CNS via segmentally repeated nerves. Peripheral glial cells in vertebrates originate from the neural crest (Jessen & Mirsky, 2005). In contrast, peripheral glial cells of Drosophila are either generated in the CNS and migrate toward their final destination in the periphery or they are generated by peripheral sensory organ precursor cells (Hilchen, Beckervordersandforth, Rickert, Technau, & Altenhein, 2008). During Drosophila development, glial cells are generally born after neuronal growth cones have navigated to their target tissues, and thus can use axonal tracts to guide migration (Aigouy, Lepelletier, & Giangrande, 2008; Berzsenyi, Kumar, & Giangrande, 2011; Klämbt, 2009; von Hilchen, Hein, Technau, & Altenhein, 2010). This process presumably requires adhesion proteins. In flies, the homophilic Ig-domain adhesion protein Fasciclin 2, the orthologue of the neural cell adhesion molecule NCAM, is expressed on both the migrating glia as well as on the neuron and plays important roles during migration of embryonic peripheral glial cells (Silies & Klämbt, 2010). Adhesion proteins might provide the means for an intensive neuron–glia cell–cell contact but they do not necessarily explain how directionality of migration is achieved (Silies & Klämbt, 2011). This could be determined by a differential expression of adhesion proteins which allows to find the final target position by maximizing (or minimizing) adhesive contacts. In the embryonic PNS of Drosophila, a graded expression of Fas2 provides an adhesive gradient to help positioning of glial cells along the axonal tract (Silies & Klämbt, 2010), whereas in the wing model repulsive N-cadherin signaling works as a molecular brake to adjust glial migration (Kumar, Gupta, Berzsenyi, & Giangrande, 2015). In leg imaginal discs, repulsive Semaphorin signaling emanating from the imaginal disc epithelium keeps migrating glial cells close to the axonal fascicle (Sasse & Klämbt, 2016). In addition, directionality of glial migration can be provided by diffusible factors. Attractive Netrin signals guide peripheral glial cell migration in the Drosophila wing (Gupta, Kumar, Cattenoz, Vijayraghavan, & Giangrande, 2016), whereas repulsive Netrin signaling from the midline directs the outward migration of the peripheral glia in the embryo (Hilchen, Hein, Technau, & Altenhein, 2010).
2.6 Wrapping gliaIn the PNS, the wrapping glia wraps individual axons or axon bundles (Stork et al., 2008). The glial cells resemble the nonmyelinating vertebrate Schwann cells, which engulf bundles of single axons to form Remak fibers. Wrapping and insulation of axons is thought to be a necessary morphological specialization to allow fast conductance of action potentials along the axon. This is most beautifully seen in myelinated nerves in the mammalian CNS and PNS which are formed by an interaction of axons with central oligodendrocytes or peripheral myelinating Schwann cells (Nave & Werner, 2014; Taveggia, 2016). The spiral wrapping of myelinating glia requires actin and is only found around large caliber axons which show fast saltatory nerve conduction (Nave & Werner, 2014; Nawaz et al., 2015; Zuchero et al., 2015). Although the developmental origin of wrapping glia and Schwann cells is clearly distinct, both glial cell types use similar molecular mechanisms to exert their function (Matzat et al., 2015). In addition to the wrapping of the axons, these glial cells have a broad range of functions in the CNS from axonal metabolic support (see below), to the control of neuronal plasticity (Fünfschilling et al., 2012; Lee et al., 2012; McKenzie et al., 2014; Tomassy et al., 2014).
Interestingly, compacted myelin is not found in invertebrates but similar morphological specializations have been identified in several invertebrate species such as copepods, earthworms, or shrimps (Davis, Weatherby, Hartline, & Lenz, 1999; Günther, 1976; Roots & Lane, 1983; Xu & Terakawa, 1999). Therefore, it appears possible that the evolutionary origin of myelin formation might have to be placed before the split of invertebrates and vertebrates (see Rey, Zalc, & Klämbt, 2020 for further information, this issue). This is also supported by the molecular mechanisms that regulate simple wrapping in the segmental nerves of the Drosophila PNS. Molecules underlying the differentiation of Drosophila wrapping glia have been identified by studying two models, the developing eye and the segmentally arranged peripheral nerves. In the developing compound eye about 50 wrapping glial cells wrap the 4,800 photoreceptor axons which project in fascicles of eight toward the first optic center, the lamina (Franzdóttir et al., 2009; Keene & Sprecher, 2012; Silies et al., 2007). On average, every wrapping glial cell wraps 12 fascicles and participates in the formation of a proper retinotopic map. Wrapping glial differentiation is controlled by a complex set of transcription factors including the homeodomain protein Cut which eventually activates EGF-receptor and FGF-receptor pathways (Bauke, Sasse, Matzat, & Klämbt, 2015). The combined activity of these receptor tyrosine kinases dictates the differentiation of glial wraps in a still unknown manner (Franzdóttir et al., 2009; Matzat et al., 2015; Sieglitz et al., 2013). Glial cells produce insulin-like peptides in response to photoreceptor-derived EGF to induce neuronal differentiation in the lamina, the first visual center in the fly brain (Fernandes, Chen, Rossi, Zipfel, & Desplan, 2017). Wrapping glial differentiation is triggered by activation of the Drosophila FGF-receptor Heartless (Htl) by the neuronally expressed FGF8-like protein Thisbe (Chang, Tsao, & Sun, 2018; Franzdóttir et al., 2009). In addition, adhesion proteins are required during neuron–glia interaction in the Drosophila eye. The heterophilic adhesion protein Borderless of the Ig-domain family is expressed only by early wrapping glial cells while they follow photoreceptor axons. These neurons in turn express the Ig-domain Turtle which participates in extension and wrapping of the axon together with FGF-receptor activity (Cameron, Chen, & Rao, 2016; Chen, Cameron, Chang, & Rao, 2017). During early wrapping glia development in the compound eye the extracellular matrix receptor Integrin is required (Xie, Gilbert, Petley-Ragan, & Auld, 2014). The integration of this pathway into the FGF-signaling pathway is currently unclear. A late acting adhesion system between photoreceptor neurons and glia is established by homophilic adhesive interactions of the Ig-domain protein Klingon (Klg) (Matsuno, Horiuchi, Ofusa, Masuda, & Saitoe, 2019; Shimozono et al., 2019). Klg is additionally expressed in a punctate pattern around the neuropil suggesting an expression on either ensheathing glia or more likely on astrocyte-like glia (Matsuno et al., 2015). Knockdown of klingon by RNA interference suppressed excessive synapse formation in the developing eye (Shimozono et al., 2019), whereas the function of Klg in ensheathing glia or astrocyte-like glial cells has not yet been determined. Moreover, Klg was found to interact with the common Dpr-interacting protein (cDIP), a secreted LRR family protein that also interacts with most members of the large Dpr Ig-domain superfamily (Özkan et al., 2013). This adds some additional levels of complexity to the interaction network established at neuron–glia interfaces.
The segmentally organized abdominal and thoracic nerves each harbor about 30 motor axons and 50 sensory axons. They are engulfed by wrapping glial cells (Matzat et al., 2015). Along the entire length of the segmental nerves only three to four of these cells are positioned. At the interface of two wrapping glial cells, some intermingling is noted (Matzat et al., 2015). Some axon bundles are followed by one glial process, whereas neighboring axons are wrapped by processes of the neighboring glial cell. Wrapping in segmental nerves is in part controlled by the activity of the Drosophila EGF-receptor which is activated by the glially expressed neuregulin-like protein Vein (Matzat et al., 2015). Interestingly, the mammalian EGF-receptor homolog as well as the activating ligand Neuregulin are both required to trigger normal myelin formation in mice (Brinkmann et al., 2008; Michailov et al., 2004; Taveggia et al., 2005). In vertebrates, however, glially expressed Neuregulin is only used during regenerative processes (Stassart et al., 2013). Also similar to vertebrate Schwann cell development, wrapping in Drosophila appears to be dependent on Integrin (Myospheroid/ßPS Integrin) and laminin (LanB2) signaling (Petley-Ragan, Ardiel, Rankin, & Auld, 2016; Xie et al., 2014). Additionally, four proteins involved in the sphingolipid biosynthesis pathway are involved cell autonomously in wrapping glial morphogenesis, namely Lace, Schlank, Infertile cresent (Des1) and Phosphoethanolamine cytidylyltransferase (Pect), whose silencing induces defects in wrapping, characterized by hypowrapping and bulge formation or swellings (Ghosh et al., 2013). Moreover, it was demonstrated that nerve conduction velocity depends on proper wrapping and is reduced upon panglial silencing of lace (Ghosh et al., 2013).
The prime task of axonal ensheathment is to allow fast propagation of action potentials. In general, neuronal signal transduction occurs via electrical impulses which travel down the axon toward the synapse to then elicit responses in other neurons or non-neuronal target cells. It has been worked out some 70 years ago that the speed of the propagation depends on axonal diameter, the resistance of the axonal cytoplasm, the capacity of the axonal membrane, and the temperature (Hodgkin & Huxley, 1952). In unmyelinated axons, a uniform distribution of the voltage-gated ion channels is assumed along the entire axon. Therefore, the action potentials have to be generated continuously along the whole length of the axon which limits conductance speed. However, the exact position of voltage-gated ion channels has not yet been determined in the Drosophila nervous system.
Another form of glial ensheathment is observed around peripheral sensory neurons that are located at exposed positions in the body wall to detect mechanical stimuli, temperature or light (Singhania & Grueber, 2014). At the example of the dorsal dendritic arborization (daIV) neurons it was shown that the wrapping glia exclusively covers axon and soma but never reaches into the dendritic compartment, which is instead encased by subperineurial and perineurial glia (Yadav et al., 2019). Here, the daIV neurons use the Ig-domain adhesion protein Neuroglian to interact with the peripheral glia which suppresses axonal sprouting and exuberant dendritic branching (Yamamoto, Ueda, Takahashi, Saigo, & Uemura, 2006). The further ensheathment of proximal dendrites also plays a role in the determination of the initial dendrite severing point during pruning (Han, Jan, & Jan, 2011).
Finally, it is interesting to note that glial wrapping around axons is well documented in the PNS, and in the nerve segments that connect the periphery with the central neuropil (Matzat et al., 2015; Stork et al., 2008; Subramanian et al., 2017). In addition, cortex glia can wrap proximal axon segments as they project toward the CNS (Coutinho-Budd et al., 2017). Serial reconstructions of electron microscopic images have failed to detect prominent glial wrapping around axons in the neuropil areas of larval and adult brains (Eichler et al., 2017; Larderet et al., 2017; Zheng et al., 2018). In adults, however, ensheathing glial cells become more complex in morphology and further subdivide the neuropil into several distinct compartments (Kremer et al., 2017). Inside the neuropil and possibly along these boundaries, ensheathing glial cells wrap axons before they enter the synaptic neuropil (Enriquez et al., 2018). Similarly, we would predict that the neck region that connects the thoracic neuropil with the neuropil in the brain lobes harbors axons wrapped by ensheathing glia.
Formation of glial membranes around neuropil axons is only seen in adult brains of swiss cheese (sws) mutants (Kretzschmar, Hasan, Sharma, Heisenberg, & Benzer, 1997). Interestingly, these membrane structures do not appear to be made by astrocyte-like glial cells (Dutta, Rieche, Eckl, Duch, & Kretzschmar, 2016), suggesting that in sws mutants, ensheathing glial cells might be able to project processes into the neuropil to enwrap neuronal processes.
2.7 Perineurial gliaThe perineurial glial cells form the outermost cell layer of the nervous system facing the hemolymph just below the neural lamella (Figure 1). They cover all parts of the nervous system and are part of the blood–brain barrier. To date no molecular differences have been detected between CNS and PNS blood–brain barrier, neither in the larva nor in the adult. In normal development perineurial glial cells do not come in direct contact with neurons. Only one perineurial–neuron interaction is known during Drosophila development: During development of the Drosophila compound eye in late third instar larvae, perineurial glia migrate along the subperineurial glia toward nascent photoreceptor neurons (Silies et al., 2007). As soon as they contact these neurons, FGF-receptor signaling triggers their differentiation into wrapping glial cells (Franzdóttir et al., 2009; Sieglitz et al., 2013). The proliferation of perineurial glial cells is guaranteed by low Öbek expression, a component of the N-end rule pathway controlling ubiquitination and subsequent proteolytic degradation of N-terminally processed proteins (Zülbahar et al., 2018). The physiological role of the perineurial glia lies in the metabolic support of the nervous system. These glial cells are in direct contact with the hemolymph and therefore express many transporter proteins responsible for the uptake of nutrients (Delgado et al., 2018; Featherstone, 2011; Volkenhoff et al., 2015; Weiler, Volkenhoff, Hertenstein, & Schirmeier, 2017). Perineurial glia respond to ethanol and promote alcohol tolerance via the A kinase anchoring protein Akap200 (Parkhurst et al., 2018). No homologous glial cell type appears to be present in vertebrates.
2.8 Subperineurial gliaTogether with the perineurial glia the subperineurial glial cells constitute the blood–brain barrier (Limmer, Weiler, Volkenhoff, Babatz, & Klämbt, 2014). Thus, as in other invertebrates this important defense and safety line is made exclusively by glia (Bullock & Horridge,
留言 (0)