Chemicals
Chemicals and reagents used were obtained from Carl Roth (Karlsruhe, Germany), Alfa Aesar (Karlsruhe, Germany), Applichem, (Darmstadt, Germany), Sigma Aldrich (Schnelldorf, Germany), Th. Geyer (Renningen, Germany) or Merck (Darmstadt, Germany). Chemicals and materials for electrophoresis were from Serva (Heidelberg, Germany) and Bio–Rad (Düsseldorf, Germany).
Strains and culture methods
Pleurotus sapidus (DSM 8266) was obtained from the German Collection of Microorganisms and Cell Cultures (DSMZ, Braunschweig, Germany). The strain was cultivated in standard nutrition solution (SNS, 30 g L−1d–(+)–glucose H2O, 4.5 g L−1l-asparagine H2O, 1.5 g L−1 KH2PO4, 0.5 g L−1 MgSO4 H2O, 3.0 g L−1 yeast extract, 1 mL L−1 trace element solution: 5 mg L−1 CuSO4 5 H2O, 80 mg L−1 FeCl3 6 H2O, 90 mg L−1 ZnSO4 7 H2O, 30 mg L−1 MnSO4 H2O, and 0.4 g L−1 EDTA; pH 6.0) for 8 days (24 °C, 150 rpm, 25 mm shaking diameter). For induction of lipase production, either 4% v/v Tween® 80 or 1% v/v corn oil were added to the main culture, replacing glucose as the main carbon source. Escherichia coli (TOP10) was obtained from Invitrogen (Karlsruhe, Germany) and was used for vector propagation. Chemically competent cells were transformed by heat shock treatment (2 min, 42 °C) according to standard protocols. Recombinant cells were cultivated in sterile LB medium (10 g L−1 tryptone, 5 g L−1 yeast extract, 10 g L−1 NaCl) with 150 mg L−1 ampicillin used as selection marker (37 °C, 225 rpm).
Secretome analysis
Secretome analysis of P. sapidus was performed at Protagen Protein Services GmbH (Dortmund, Germany) by 2D electrophoresis and subsequent MALDI-MS/MS analysis. Sequence homologies of the obtained fragments were analyzed using BLAST (Altschul et al. 1990) and Pfam (Sonnhammer et al. 1997) database.
cDNA-synthesis
To isolate RNA, the mycelium of a submersed culture containing SNS + Tween® 80 was harvested on day 5 and 100 mg was ground in liquid nitrogen with mortar and pistil. Isolation of total RNA was performed with RNeasy™ Plant Mini Kits (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. The quality of the RNA was verified by an 1% (w/v) agarose electrophoresis and ethidium bromide staining. A cDNA library of P. sapidus was produced using the isolated RNA as template with the SMART™ PCR cDNA Synthesis Kit (Takara Bio Europe S.A.S., Saint-Germain-en-Laye, France) for 21 to 24 cycles (Additional file 1: Fig. S1). Amplification of a coding GDS(L)-like hydrolase sequence of P. sapidus from the cDNA library was performed by PCR. The primers were derived from a homologous P. ostreatus putative GDSL type lipase sequence (Genbank accession number: KAF7436219.1, Additional file 1: Table S1). Amplification of the specific cDNA was performed in an Alpha SC PCR Thermocycler (Analytik Jena, Jena, Germany). The following PCR protocol was used: 50 ng template, 5 × PCR-Puffer including dNTP’s (Qiagen), forward and reverse primer each 50 pmol, 1.25 U HotStar HiFidelity DNA-Polymerase (Qiagen), ddH2O ad 50 µL, 95 °C 5 min,–95 °C 1 min, 53 °C 1 min, 72 °C 90 s–for 40 cycles 72 °C 5 min. PCR products were separated electrophoretically (1.2% (w/v) agarose gel, (Additional file 1: Fig. S2), subsequently isolated from the gel using NucleoSpin Extract II Kit (Macherey–Nagel, Düren, Germany) and finally ligated (Topo TA-Cloning® Kit, Invitrogen) into the vector pCR2.1–TOPO® (Invitrogen). The plasmid DNA was replicated in E. coli TOP10 cells (Invitrogen), isolated from the cells and purified (NucleoSpin® Plasmid DNA Purification, Macherey–Nagel). Sequencing of the cloned cDNA was performed by Eurofins Genomics GmbH (Ebersberg, Germany). The codon-optimized PSA_Lip sequence can be found in the supplementary file.
Enzyme production
For recombinant production of the PSA_Lip, the codon usage of the gene was adapted to the host organism T. reesei (a derivative of RUT C30) (Peterson and Nevalainen 2012) and an expression cassette was constructed from the adapted sequence. The expression of lipase was investigated with three different constructs. pAB500-LipPS, in which the lipase gene with its signal sequence was under control of the cbhI (cellobiohydolase I) promoter and the cbhI terminator of T. reesei, was used to generate strain RH32919. pAB510-LipPS, in which the lipase gene was under control of the same cbhI promoter and cbhI terminator and secretion was mediated by the cbhI signal sequence was used to generate strains, which are the parallel transformants from same transformation, RH32924, RH32926 and RH32927; pAB600-LipPS, in which the lipase gene without signal sequence was fused with a cbhII (cellobiohydrolase II) carrier was used to generate the strains RH32928 and RH32929. The fermentation process was investigated in batch and fed-batch mode. Efficient production of the lipase was achieved by optimization of the fermentation process used batch conditions (pH5.5, 28 °C, 161 h) in monosaccharide medium D5 (5% monosaccharides, 0.7% (NH4)2SO4, 0.3% KH2PO4) (Additional file 1: Fig. S3). The supernatants were used for determination of enzyme activity (Additional file 1: Fig. S4).
Determination of enzyme activity
Enzyme activity was determined photometrically using a temperature controlled multi-mode plate reader (Synergy™ 2, BioTek Instruments GmbH, Bad Friedrichshall, Germany) by performing esterase and lipase activity assays with various p-nitrophenol (pNP) esters (Purdy and Kolattukudy 1973; Winkler and Stuckmann 1979). For the hydrolysis of pNP-acetate (pNPA), pNP-butyrate (pNPB), pNP-valerate (pNPV), pNP-hexanoate (pNPH), and pNP-octanoate (pNPO), an esterase assay was performed using the following conditions: 130 μL of potassium phosphate buffer (80 mM, pH 7.0) supplemented with 0.5% v/v Triton X-100 (without Triton X-100 for pNPA) was mixed with 20 μL of enzyme solution in a 96-well microplate, starting the reaction by adding 50 μL of substrate solution (3.5 mM of one pNP ester). The reaction was monitored for 10 min at 405 nm and 30 °C. To determine lipase activity, 50 μL of the enzyme solution was mixed with 200 μL of the substrate solution: 0.4 mM pNPP in potassium phosphate buffer (50 mM, pH 8.0), which was supplemented with 2.2 g L−1 sodium deoxycholate and 1.1 g L−1 gum arabic. The activity was measured at 37 °C for 10 min. For both assays, one unit of enzymatic activity refers to the amount of enzyme that hydrolyzes 1 μmol of pNP ester per minute under the respective conditions (esterase assay: εp-nitrophenol = 0.00985 L μmol−1 cm−1 (Purdy and Kolattukudy 1973), lipase assay: εp-nitrophenol = 0.0183 L μmol−1 cm−1 (Winkler and Stuckmann 1979)). Besides the pNP-esters, glyceryl trioctanoate (TriC8:0) (Merck, 0.12 mol L−1 stock solution dissolved in DMSO) as a triacylglyceride was used as substrate. The activity was measured in 5 mM HEPES buffer (pH 8.0) with a non-esterified free fatty acids (NEFA) colorimetric assay kit (Elabscience Bionovation Inc., Houston, TX, USA) according to the manufacturer’s protocol after 1, 2 and 4 h of reaction at 30 °C, 950 rpm. Phospholipase A activity was examined with the EnzChek® phospholipase A2 assay kit (Thermo Fisher Scientific, Dreieich, Germany) according to the manufacturer’s manual. PSA_Lip was also tested for choline esterase activity with an acetylcholinesterase activity assay kit (Sigma-Aldrich) following the manufacturer’s instructions. Feruloyl esterase activity was examined by two methods, a photometric assay, modified from Ralet et al. (1994) and an HPLC method as demonstrated by Linke et al. (2013). The photometric assay is based on a shift of absorption maximum from 322 nm, corresponding to methyl ferulate, to 293 nm, which indicates formation of free ferulic acid at pH 6.0. The assay mixture was composed of 50 µL enzyme solution mixed with 940 µL of 1.3 mM ferulic acid methyl ester, 1 mM sodium azide and 2% (v/v) ethanol in 100 mM MOPS buffer (pH 6.0). Heat inactivated enzyme (incubated 10 min at 95 °C) served as control. After incubation at 37 °C and 200 rpm for 72 h, a UV/Vis spectrum from 280 to 340 nm was recorded. To detect feruloyl esterase activity via HPLC–DAD (LC-20 with Degaser DG-20A5R, quaternary pump LC20AD, Autosampler SIL-20ACHT, column oven CTO-20AC, Controller CBM-20A and PDA-Detector SPD-M20A, Shimadzu, Kyoto, Japan) at 325 nm, 125 µL enzyme solution was mixed with 375 µL substrate solution (100 mM tartrate–succinate buffer pH 6.0, 3% (v/v) DMSO, 2 mM 5-O-transferuloyl-arabino-furanose), incubated at 37 °C for 17 h, cooled to 4 °C, and supplemented with 500 μL acetonitrile. Heat-inactivated enzyme served as a control. 1 mM sodium azide was used to inhibit laccases. Prior to analysis, the samples were filtered through 10 kDa MWCO centrifugal filter units (Merck). 10 µL of filtrate was injected into HPLC–DAD for separation of substrate and free ferulic acid using a 250/4 Nucleosil 100–5-column (Macherey–Nagel, Düren, Germany). The mobile phase consisted of 0.05% (v/v) formic acid in water (A) and 0.05% (v/v) formic acid in acetonitrile (B). The following gradient was run: 90% A (1 min), linear gradient from 90 A to 0% A over 10 min, 0% A (5 min), linear gradient from 0 to 90% A over 1 min, 90% A (2 min) at 1.5 mL min−1 flow (Additional file 1: Fig. S5).
The acetylxylan esterase activity of PSA_Lip (0.3 U based on pNPO activity) towards d-xylofuranose tetraacetate (4 mM) and α-d-(+)-glucose pentaacetate (4 mM, in 10% ethanol) dissolved in potassium phosphate buffer 80 mM pH 8 was determined by TLC using pre-coated plates (Silica Gel 60 F254, 0.25 mm; Merck) with water/methanol/ethyl acetate, 1:2:7 (v/v), as the mobile phase. Sugars were visualized with a solution containing 120 g of (NH4)Mo7O24 and 5 g of (NH4)2Ce(NO3)6 in 800 mL of 10% H2SO4. The fully deacetylated sugars were used as control (4 mM). A reaction with an acetylxylan esterase from Orpinomyces sp. (5 U, Megazyme, Bray, Ireland) was used as positive control. The reactions were carried out at 30 °C and 600 rpm (HLC Cooling-ThermoMixer MKR 23, DITABIS - Digital Biomedical Imaging Systems AG, Pforzheim, Germany) and monitored over 24 h, while samples were taken at different time points (0.25 h, 4 h, 24 h) (Additional file 1: Fig. S6).
Protein purification and mass spectrometry-based protein identification
The T. reesei supernatant containing recombinant PSA_Lip was purified using size-exclusion chromatography (HiLoad 16/60 Superdex 200, Cytiva, Uppsala, Sweden) with 0.6 mL min−1 flow with potassium phosphate buffer (80 mM, pH 7.0) as mobile phase. The fractions with the highest hydrolytic activity against pNPO were collected for gel electrophoresis and subsequent mass spectrometric analyses. The fractions were concentrated via ultrafiltration using a centrifugal filter unit (10 kDa MWCO, Merck). The purified and filtered fractions were mixed with semi-native loading buffer and semi-native sodium dodecyl sulfate poly acrylamide gel electrophoresis (SDS-PAGE) (semi-native refers to: no heat denaturation, no mercaptoethanol, 50% SDS concentration in all buffers) was performed adapted from Laemmli using a 4% stacking and 12% resolving gel and Coomassie Brilliant Blue R250 staining (Additional file 1: Fig. S7) (Laemmli 1970). Respective proteins were cut from the gel and mixed with RapiGest™ SF solution (Waters GmbH, Eschborn, Germany), approx. 10 µL per 15 µg protein, and incubated for 20 min at 80 °C. After cooling, 5 µL 100 mM dithiothreitol (DTT) was added and the mixture was heated to 60 °C for 20 min. Then 5 µL of 200 mM 2-iodoacetamide (IAA) was added and the mixture was incubated for 30 min in the dark at room temperature. Afterwards, the solution was mixed with 1.5 µL trypsin (1 µg L−1, Promega GmbH, Mannheim, Germany) and incubated overnight at 37 °C. The samples were purified using ZipTip® filter tips as described in the instruction manual (Merck, Darmstadt, Germany). After vacuum drying, the samples were resuspended in 3% acetonitrile/0.1% formic acid (v/v in H2O). The resulting peptides were separated by UltiMate 3000 RSLC UHPLC system (Thermo Fisher Scientific) equipped with a Kinetix C18 column (Phenomenex, 2.6 µm, 100 Å, inner diameter 2.1 mm, 100 mm length) coupled with a Q Exactive HF-X Orbitrap (Thermo Fisher Scientific, Bremen, Germany) mass spectrometer. Chromatographic analysis was performed at a flow rate of 250 μL min−1 with water/0.1% formic acid (mobile phase A) and acetonitrile/0.1% formic acid (mobile phase B). The gradient elution of 60 min was performed as follows: 3% B for 5 min, followed by 3−40% B over 40 min, 40−50% B over 5 min, 50−98% B over 5 min, and re-equilibration in 3% B. The mass spectrometer was operated in data-dependent acquisition mode (top-15 DDA) with the following parameters in full MS scans: a mass range of m/z 350–1800, mass resolution of 120,000, automatic gain control (AGC) target of 3 × 106, injection time (IT) of 50 ms, and MS/MS scans with mass resolution of 30,000, AGC target of 1 × 105, IT of 120 ms, isolation window m/z 1.3, and dynamic exclusion of 60 s. The raw files were processed using Proteome Discoverer version 2.4 (Thermo Fisher Scientific) with SEQUEST and MS Amanda search engines against the PSA_Lip databases (Additional file 1: Fig. S8).
Biochemical characterization
For detailed characterization of P. sapidus lipase (PSA_Lip), photometric esterase and lipase assays (see: Determination of enzymatic activity) were performed. For determination of temperature and pH optima, as well as stability, pNPO was used as model substrate. The optimum reaction temperature was determined by incubation of assay solutions and using the heater of the micro plate reader at the respective temperature. To evaluate the temperature stability, portions of 100 µL enzyme solution were incubated for 60 min at the respective temperature. The graphical analysis of measured activities in relation to the temperature formed the typical sigmoid curve. Its turning point can be calculated by fitting the Y-values and is equivalent to the T5060 value of PSA_Lip, which indicates a decrease of enzymatic activity by 50% after incubation at this temperature for 60 min. To determine the pH optimum, the esterase assay was performed using 100 mM Davies buffer (Davies 1959), which was set to the desired pH. PSA_Lip was incubated for 60 min at different pH values. The data obtained with different concentrations of each substrate for the Michaelis–Menten kinetics were analyzed with OriginPro® 2021 (category: enzyme kinetics, function: Michaelis–Menten, iteration algorithm: Levenberg Marquardt, without weighting). Equation for the Michaelis–Menten fit:
$$y=\frac_ \cdot x}_ + x}$$
Protein concentrations were determined by Bradford assay, ROTI® Nanoquant (by Carl Roth), following the manufacture’s manual.
Bioinformatic and phylogenetic analysis
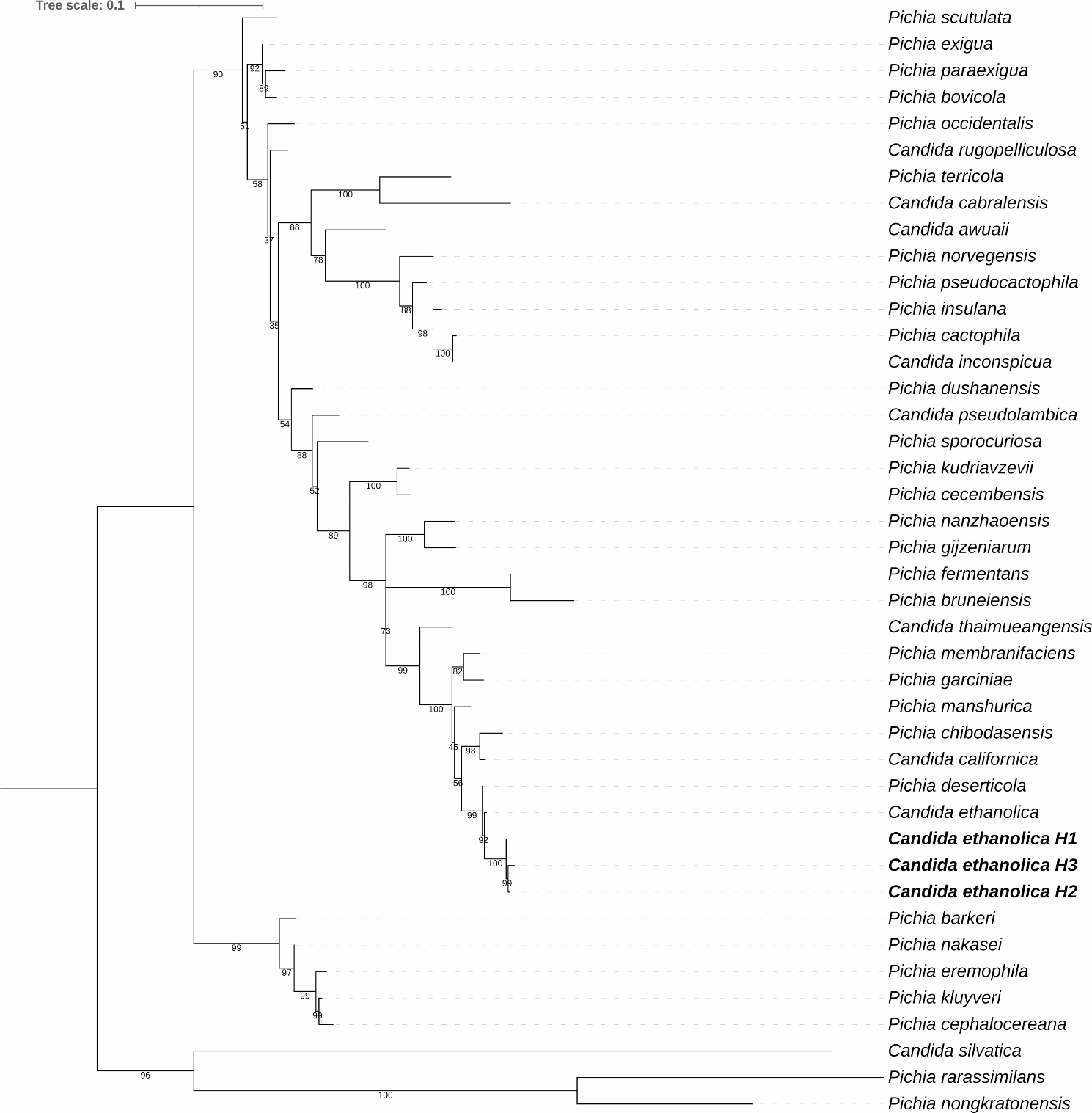
For better understanding of the potential classification of PSA_Lip, a PSI-BLAST (Altschul et al. 1990) search with the non-redundant protein database was conducted, where the scoring matrix BLOSUM62 and gap cost of 11 for existence and 1 of extensions with a PSI-BLAST threshold of 0.005 were used. The twelve most similar sequences to the PSA_Lip, 27 GDSL lipases from Akoh et al. (2004), and seven CE16 family proteins from http://www.cazy.org/ as well as seven α/β-lipases from Basdiomycota and yeast were used to perform a multiple sequence alignment with ClustalW (Thompson et al. 1994). The phylogenetic tree generated using Geneious v.9.1 (Biomatters, Ltd., Auckland, New Zealand), was inferred under the maximum likelihood (ML) (Guindon et al. 2010) by using the PhyML plugin, substation model (Le Gascuel) LG with 100 bootstraps, all other parameters were set as default (Guindon et al. 2010). The calculation of identities and similarities of the PSA_Lip was done with the EMBOSS Stretcher online tool (.ebi.ac.uk/Tools/psa/emboss_stretcher) (Myers and Miller 1988) and the consensus sequence was visualized with WebLogo v.3 (Crooks et al. 2004).
A hybrid homology model was built based on the PSA_Lip sequence using the automatic homology modelling script of YASARA v.17.1.28 (Krieger et al. 2002). This script automatically performs sequence alignments, loop building, side chain modelling, and energy minimization (Krieger et al. 2002). Herein, the script ran three PSI-BLAST (Altschul et al. 1997) iterations to build a total of 25 models based on five template structures (PDB codes: 6UQV, 6JLZ, 5XTU, 3KVN and 4HYQ). Out of the 25 models, the best parts were automatically chosen to construct a hybrid model. Structural alignments using the homology model and structures from the protein database were performed with Pymol™ v.2.3.4. The ExPASy compute pI/Mw tool (Gasteiger et al. 2005) was used to calculate molecular mass and isoelectric point of the protein. Potential homologies to carbohydrate-active enzymes were analyzed by searching the dbCAN meta server (Zhang et al. 2018) based on the PSA_Lip sequence.







留言 (0)