
記住我




With few exceptions, terrestrial organisms require iron for growth, but the limited solubility of the predominant aqueous form of iron, ferric oxyhydroxide [Fe(OH)n; KSP = 2.8 × 10−39; Neilands, 1974], obstructs its biological utilization. Consequently, microorganisms produce siderophores (Neilands, 1995) that solubilize Fe3+ from its insoluble precipitates, liberating the metal for uptake and incorporation into cellular metabolism. The tricatecholate Gram (−) bacterial siderophore enterobactin (Ent) is an avid iron chelator [KA = 1052 M−1; Harris et al., 1979]. Some pathogenic bacteria glucosylate Ent (GEnt; also called salmochelin Hantke et al., 2003) to evade siderocalin (Doneanu et al., 2004; Holmes et al., 2005), an innate immune system protein that tightly binds Ent (but not GEnt) (Kumar et al., 2022) and removes it from circulation. The phenolic chelation groups of Ent and GEnt may oxidize to quinones, and their trilactone scaffolds may hydrolyze to yield mono- and dicatecholates (Ent*). In response to these phenomena, Gram (−) bacteria may utilize both FeEnt and FeGEnt, their dicatecholate hydrolysis product ferric (dihydroxybenzolyserine)2 (Fe(DHBS)2; FeEnt*), and monocatecholates (dihydroxybenzoic acid (FeDHBA3); FeDHBS3). The E. coli cell envelope transport systems for ferric catecholates encompass outer membrane (OM) receptors (Fiu, FepA, Cir, IroN Klebba et al., 2021), periplasmic binding proteins (FepB Thulasiraman et al., 1998; Sprencel et al., 2000), inner membrane (IM) ABC-transporters FepCDG (Shea and McIntosh, 1991), and Ent hydrolases/membrane reductase activities (Fes, IroD, IroE) (Brickman and McIntosh, 1992; Lin et al., 2005; Zhu et al., 2005; Caza et al., 2015). Monocatecholate Trojan horse siderophore antibiotics (Budzikiewicz, 2001; Wencewicz et al., 2013; Simner and Patel, 2020) enter bacterial cells by the same pathways.
Gram (−) bacterial OM receptors for metal complexes are also called TonB-dependent transporters [TBDT; (Schauer et al., 2008)] or ligand-gated porins [LGPs; (Rutz et al., 1992)]. The initial LGP crystal structures of FepA (Buchanan et al., 1999) and FhuA (Locher et al., 1998) revealed C-terminal, ~650 a.a., 22-stranded, transmembrane β-barrels filled by N-terminal, ~150-a.a. globular domains (Locher et al., 1998; Buchanan et al., 1999; Ferguson et al., 2002; Shultis et al., 2006; Buchanan et al., 2007; Grinter and Lithgow, 2019a). The external loops of LGP β-barrels initiate iron acquisition (Smallwood et al., 2014) by binding particular iron complexes (Klebba et al., 2021; Kumar et al., 2022). The internal N-domain controls the ensuing TonB-dependent stage of ligand passage into the periplasm. TonB (Noinaj et al., 2010) is a cell envelope protein that facilitates active OM iron transport, through connection with electrochemical proton motive force (PMF) across the IM (Bradbeer, 1993; Jordan et al., 2013). TonB anchors in the IM but spans the periplasm to interact with ligand-bound LGP at the periplasmic interface of the OM. Overall, LGPs use PMF, mediated by TonB, to transport ferric siderophores through the OM (Jordan et al., 2013). The structural features of LGP raised the possibility of ligand transport by a “Ball and Chain” mechanism (Armstrong and Bezanilla, 1977), in which the N-terminus dislodges from the channel into the periplasm (Ma et al., 2007). Molecular dynamics simulations suggested that this process may occur by unfolding of the N-terminal globular domain (Faraldo-Gómez et al., 2003; Gumbart et al., 2007), but subsequent experiments concluded that the N-domain of FepA remains within the transmembrane channel and undergoes conformational rearrangements during FeEnt uptake (Majumdar et al., 2020). At present, the molecular mechanism of ferric siderophore movement through OM LGP is an open question.
High-affinity ferric siderophore uptake by TonB-dependent LGP enables Gram-negative bacterial iron acquisition in iron-deficient environments, including within human and animal organs, tissues, fluids, and secretions, where transferrin, siderocalin, and ferritin sequester iron as a defense mechanism (Cornelissen and Sparling, 1994; Abergel et al., 2008; Correnti and Strong, 2012). Consequently, siderophore biosynthetic systems and TonB-dependent ferric siderophore transport systems are determinants of bacterial pathogenesis, including in the ESKAPE organisms (Russo et al., 2002, 2011, 2015): disruption of iron uptake reduces virulence. In Acinetobacter baumannii, for example, iron acquisition strongly correlates with bacterial pathogenesis (Gaddy et al., 2012; Penwell et al., 2015; Fleming et al., 2017; Luna et al., 2019; Ramirez et al., 2019; Escalante et al., 2023). Furthermore, orthologs of E. coli Fiu and Cir (PiuA and PirA, respectively) were implicated in the uptake of the novel Trojan horse antibiotic cefiderocol (FDC) in both Pseudomonas aeruginosa (Luscher et al., 2018; Gupta et al., 2022) and A. baumannii (Luscher et al., 2018; Malik et al., 2020; Asrat et al., 2023; Smoke et al., 2023; Tiseo et al., 2023). Siderophore antibiotics use native, TonB-dependent iron uptake pathways for entry into bacterial cells (Miller et al., 1991; Ji et al., 2012; Miller and Liu, 2021).
Fiu [Ferric iron uptake; (Hantke, 1983)] is an LGP (Grinter and Lithgow, 2019a) that was implicated in the uptake of ferric monocatecholates (Curtis et al., 1988; Hantke, 1990; Nikaido and Rosenberg, 1990). It occurs in commensal E. coli and in clinically relevant members of Enterobacterales, such as pathogenic E. coli, Klebsiella pneumoniae, and A. baumannii (Klebba et al., 2021). While E. coli FepA (EcoFepA) was originally identified as the cognate receptor for FeEnt, Fiu and Cir were proposed to transport ferric monocatecholates, such as FeDHBS and FeDHBA (Hantke, 1990). Other studies showed Fiu-mediated uptake of catecholate siderophore antibiotics (Curtis et al., 1988; Nikaido and Rosenberg, 1990). However, previous research did not define Fiu’s ligand preferences, binding affinities, nor the mechanism by which it captures and internalizes ligands. In this report, we characterized the selectivities of known and putative OM catecholate receptors of E. coli (Fiu, EcoFepA, and Cir) and A. baumannii (PiuA, AbaFepA, PirA, and BauA), for different catecholate iron complexes, using sensitive fluorescent technology to monitor their binding reactions (Hanson et al., 2016; Nairn et al., 2017; Chakravorty et al., 2019; Kumar et al., 2022). Fiu-FM bound FeEnt, FeGEnt, FeEnt*, FeDHBS, FeDHBA, FeFDC, and FeMB-1. The fluorescent constructs defined the hierarchy of Fiu binding affinities among these iron complexes: FeDHBS (KD = ~0.1 μM); FeGEnt (KD = 0.34 μM); FeEnt* (KD = 0.7 μM); FeDHBA (KD = 0.7 μM); FeEnt (KD = 0.83 μM). However, despite its broad binding activity, Fiu only catalyzed the uptake of FeEnt*, the dicatecholate degradation product of FeEnt: Fiu did not transport FeDHBA, FeDHBS, FeEnt, or FeGEnt. Finally, in silico docking experiments and molecular dynamic simulations found 3 FeEnt* binding sites in Fiu’s vestibule and a fourth site deeper in its interior. We used Ala scanning mutagenesis to study the outer sites and identified residues in each location that affected the binding and transport of FeEnt*. Double mutations involving R142A in site 2 created a 20-fold decrease in overall binding affinity and abrogated FeEnt* uptake. Overall, the analysis explained the pathways of ferric catecholate uptake in E. coli and A. baumannii and provided insight into the general mechanism of metal transport through TonB-dependent LGP.
Results Purification and characterization of natural ferric catecholate siderophoresWe purified catecholate ferric siderophores from the pathogenic E. coli strain CP9. After culturing it to late log phase in iron-deficient T-media (McIntosh and Earhart, 1976), we clarified the spent culture supernatant by centrifugation, added FeCl3 to form ferric siderophores, and purified them by ion exchange, gel filtration, and hydrophobic chromatography (Sephadex LH20), which segregated three distinctly colored red or purple fractions (Supplementary Figure S1; Kumar et al., 2022). The third peak was FeEnt, as identified by its absorption maximum at 495 nm, and by mass spectrometry, which showed a peak at 729.6 m/z (Supplementary Figure S1), in close correspondence to the mass of FeEnt (MW = 719). FeEnt is chemically labile: in aqueous buffers, it degrades over days or weeks, even on ice. As FeEnt storage time passed, the amount of peak 3 decreased and peak 2 increased, consistent with peak 2 as a degradation product of FeEnt. Hence, we designated the 2nd LH20 fraction as FeEnt*. It had an absorption maximum at 503 nm, and mass spectrometry defined it as 554.6 m/z, near the mass of Fe(DHBS)2 (MW = 518.4). Four catechol oxygens complex iron (III) in dimeric FeEnt*; in the hexacoordinate ferric complex, two more oxygens may derive from water molecules. When included in mass calculations of FeEnt* [i.e., Fe(DHBS)2·2H2O], the water molecules raised its mass to 554.4, which matched the mass spectrometric peak at 554.6 m/z. We also characterized the recognition of FeGEnt by Fiu. The glucosylated ferric catecholate does not adsorb well to siderocalin (Kumar et al., 2022), which promotes the pathogenesis of clinical isolates of E. coli, K. pneumoniae, and Salmonella typhimurium in human and animal hosts (Russo et al., 2011, 2015). We purified FeGEnt from E. coli CP9. It comprised the rapidly migrating first peak from LH20, with an extinction maximum at 510 nm and an MS value of 1076.3 m/z, consistent with its predicted molecular mass of 1077.5. Although the peak fraction containing FeEnt* was nearly homogeneous, the fractions containing FeEnt and FeGEnt contained other chemical species, slightly different in mass. This result underscores the chemical lability of FeEnt/FeGEnt and/or their susceptibility to cellular degradative processes during biosynthesis (Lin et al., 2005). Nevertheless, the FeEnt peak fractions did not contain FeEnt*, and the FeGEnt peak fractions did not contain FeEnt. Overall, the procedures yielded FeEnt, FeEnt*, and FeGEnt for use in binding and uptake experiments.
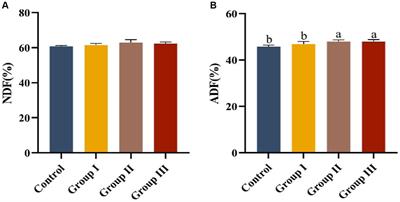
Siderophore nutrition tests of ferric catecholate transport by FiuPrevious reports of ferric monocatecholate uptake by Fiu (Hantke, 1990; Nikaido and Rosenberg, 1990) suggested that it may transport the dicatecholate degradation product, FeEnt*. To test this idea, we compared isogenic chromosomal mutants of BN1071 (Klebba, 1981; Klebba et al., 1982) lacking (Ma et al., 2007) FepA (OKN3: fiu+, ΔfepA; cir+), Cir (OKN5; fiu+ fepA+: Δcir), or Fiu (OKN9: Δfiu fepA+; cir+) to the wild-type and the triple deletion mutant (OKN359: Δfiu/ΔfepA/Δcir). We also expressed chromosomal Fiu as sole catecholate-specific LGP (OKN35: fiu+ ΔfepA; Δcir) and from the low-copy vector pHSG575 in OKN359. With these and other test strains, we screened the catecholate ferric siderophores FeEnt, FeEnt*, FeGEnt, FeDHBS, and FeDHBA in nutrition tests (Figure 1A; Table 1) that revealed a variety of uptake pathways. Strains that exclusively expressed Fiu grew around disks containing FeEnt*, with halos that were comparable in size and density to those produced by wild-type E. coli. In addition to definitive growth around disks containing FeEnt*, Fiu+ strains showed very faint, almost imperceptible growth around FeEnt, but no growth around FeDHBA nor FeDHBS. Cells expressing FepA, on the other hand, acquired FeEnt, FeEnt*, and FeDHBA (confirming that the assay functioned properly with FeDHBA). These data demonstrated the uptake specificity of Fiu for FeEnt* but not the ferric monocatecholates. Bacteria singly expressing Fiu, FepA, or Cir all utilized FeEnt*, revealing three uptake pathways for the ferric dicatecholate degradation product. In summary, Fiu transported FeEnt*, very weakly transported FeEnt, but did not utilize FeDHBS nor FeDHBA. Unfortunately, nutrition tests were uninformative for FeGEnt, which produced identical small halos on all the test strains, including E. coli BN1071 (Fiu+, FepA+, Cir+) and OKN359 (Δfiu/ΔfepA/Δcir). Hence, the assay did not provide evidence of specific FeGEnt uptake by any of the ferric catecholate LGP. However, the uptake of FeGEnt requires not only an OM receptor but also the iro gene cluster, that encodes the enterobactin trilactone hydrolases IroD and IroE (Lin et al., 2005). They are present in S. enterica and certain pathogenic E. coli such as uropathogenic E. coli 563 (Hantke et al., 2003), but the laboratory E. coli host strains we employed lack the iro system needed for the transport of FeGEnt.
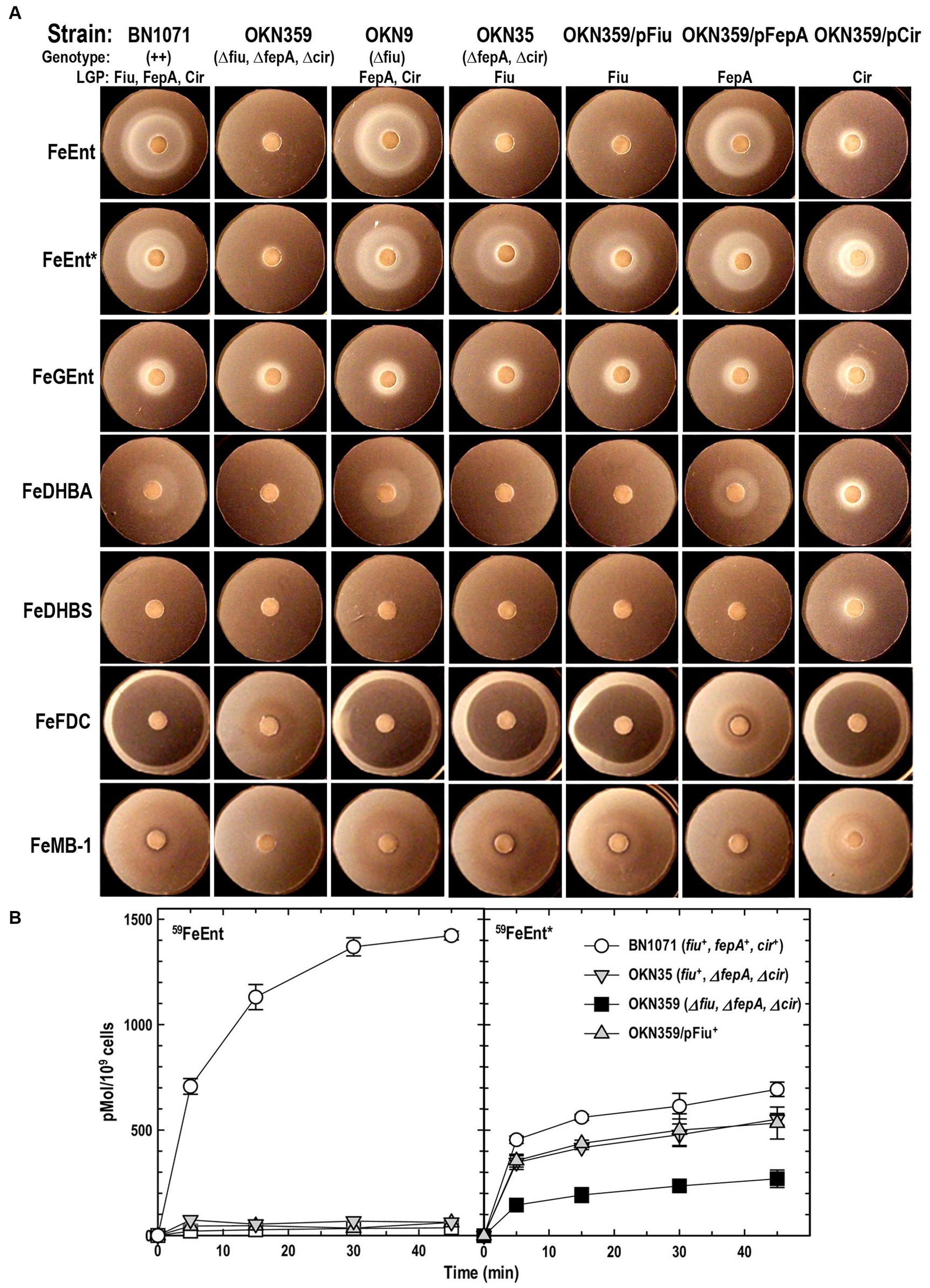
Figure 1. Uptake of ferric catecholates by Fiu, FepA, and Cir. (A) Siderophore nutrition tests. We compared strains that selectively expressed Fiu, FepA, and Cir by siderophore nutrition tests (Wayne et al., 1976), and for susceptibility to siderophore antibiotics (FeFDC or FeMB-1), by applying 10 μL of 100 μM solution of test compound to a paper disk on the agar. After overnight incubation at 37°C, we measured the diameter of the growth or killing halos of the siderophore or antibiotic, respectively, with a ruler. (B) 59Fe accumulation assays. We grew bacteria expressing Fiu, FepA, and Cir, or isogenic derivatives that only expressed Fiu, in MOPS minimal media, added 10 μM [59Fe]Ent or [59Fe]Ent* at t = 0, and collected and filtered aliquots at sequential times. We measured the radioactivity retained on the filters to evaluate the uptake of ferric siderophores. Error bars represent the standard deviations of the means of three independent trials.
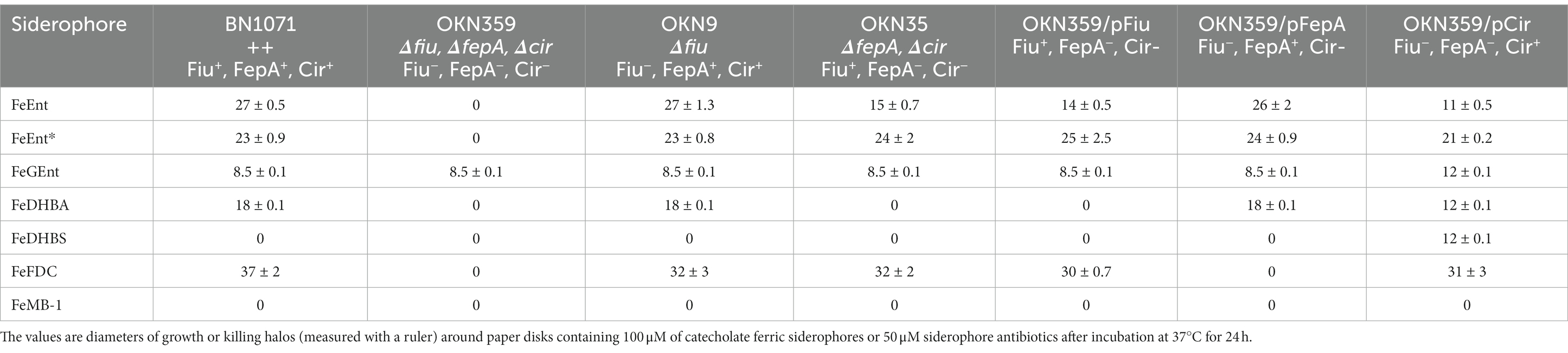
Table 1. Siderophore nutrition and antibiotic susceptibility tests.
Bacteria expressing Fiu or Cir were highly susceptible to the iron complex of cefiderocol (FDC; Figure 1), which only requires OM transport to achieve bacteriocidal activity in the periplasm. Ferric siderophores, on the other hand, require both OM and IM transport for growth stimulation. Therefore, aside from revealing OM transport, the large FeFDC killing zones are not relatable to the sizes of siderophore nutrition test halos.
Radioisotopic measurements of ferric siderophore uptakeRadioisotopic iron accumulation measurements (Figure 1B) confirmed the preference of EcoFiu for FeEnt* that was observed in microbiological assays. The left panel in Figure 1 shows the uptake of [59Fe]Ent by FepA, a high affinity (KD = 0.3 nM; Newton et al., 1999), specific process. Consequently, background uptake is negligible when we assayed FepA-deficient cells with nanomolar levels of FeEnt. The right panel, on the other hand, shows the uptake of [59Fe]Ent*, a lower affinity (KD = 0.74 μM; Figure 2) process that is accomplished by Fiu, FepA, or Cir (Figures 1A, 2). The background of FeEnt* uptake by OKN359 (Δfiu, ΔfepA, Δcir) was higher, suggesting either non-specific adsorption of FeEnt*, or the presence of another, as yet unidentified uptake pathway. Nevertheless, bacteria expressing Fiu accumulated FeEnt* to levels that were 2–3 times higher than the background, confirming its ability to transport the degradation product. In summary, the wild-type parental strain BN1071, which does not produce any siderophores (entA) but expresses Fiu, FepA, and Cir, accumulated both [59Fe]Ent and [59Fe]Ent*, whereas two strains that expressed Fiu as their sole catecholate-specific LGP, OKN35 and OKN359/pEcoFiu (fiu+), acquired only [59Fe]Ent* but not [59Fe]Ent. These data reiterated the siderophore nutrition test results that EcoFiu transported FeEnt* but not FeEnt.

Figure 2. Ferric catecholate receptor binding titrations. We determined binding affinities (see Supplementary Figure S2), by titrating OKN1359 harboring plasmids expressing the noted Cys mutant receptors with ferric catecholates and plotting the concentration dependence of the ensuing fluorescence quenching. For each trial, after measuring each data point in triplicate, we calculated the mean fluorescence quenching and its standard deviation and then plotted 1−F/F0 versus ligand concentration. Data points represent the mean of the triplicate values, with associated standard deviations for trials with FeEnt, FeEnt*, and FeFDC. We fit the data to the 1-site with a background binding model of GraFit 6.0.12 that yielded the titration curves and resulting KD values (see Table 2). The data depict a hierarchy of affinities among the different receptors for tri-, di-, and monocatecholate iron complexes. Relative to the prototypic high-affinity receptors EcoFepA and AbaFepA (Kumar et al., 2022), Fiu, Cir, and their orthologs/paralogs in A. baumannii show much lower affinity, focused on ferric di- and monocatecholates. The figure also illustrates the ability of the sensor library to monitor affinities over a 7-log range of concentrations, from sub-nanomolar to millimolar concentrations.
Fluorescence spectroscopic measurements of receptor-ligand bindingTo monitor the adsorption of ligands by EcoFiu, we engineered single Cys substitutions in its surface loops and alkylated them with FM (Supplementary Figure S2). When LGPs bind iron complexes, attached fluorophores may undergo concentration-dependent quenching, from direct contact between the ligand and the fluorophore, or conformational changes that relocate the fluor, energy transfer, or other mechanisms (Payne et al., 1997). Plots of fluorescence intensity versus [ligand] reveal binding affinity. Based on the Fiu crystal structure (Grinter and Lithgow, 2019a), we introduced substitutions S347C, T414C, S555C, and A694C in EcoFiu (Supplementary Figure S2). All the Cys mutants were active in siderophore nutrition tests with FeEnt* (data not shown). When we assessed the fluoresceination of the mutants in living bacterial cells, Fiu_A694C-FM showed the best labeling, so we used it to monitor the binding of different ligands. Titrations of OKN1359/pFiu_A694C-FM with increasing amounts of FeEnt, FeEnt*, FeGEnt, FeDHBA, FeDHBS, FeFDC, or FeMB-1 yielded KD values for the binding interactions (Figure 2; Table 2). This fluorescent sensor methodology may also be used for rapid, stopped-flow measurements that resolve the different binding stages (Smallwood et al., 2014). However, the affinity determinations that we report herein yielded the overall affinity of the binding reaction at equilibrium, in the absence of transport (i.e., in TonB-deficient host strains).
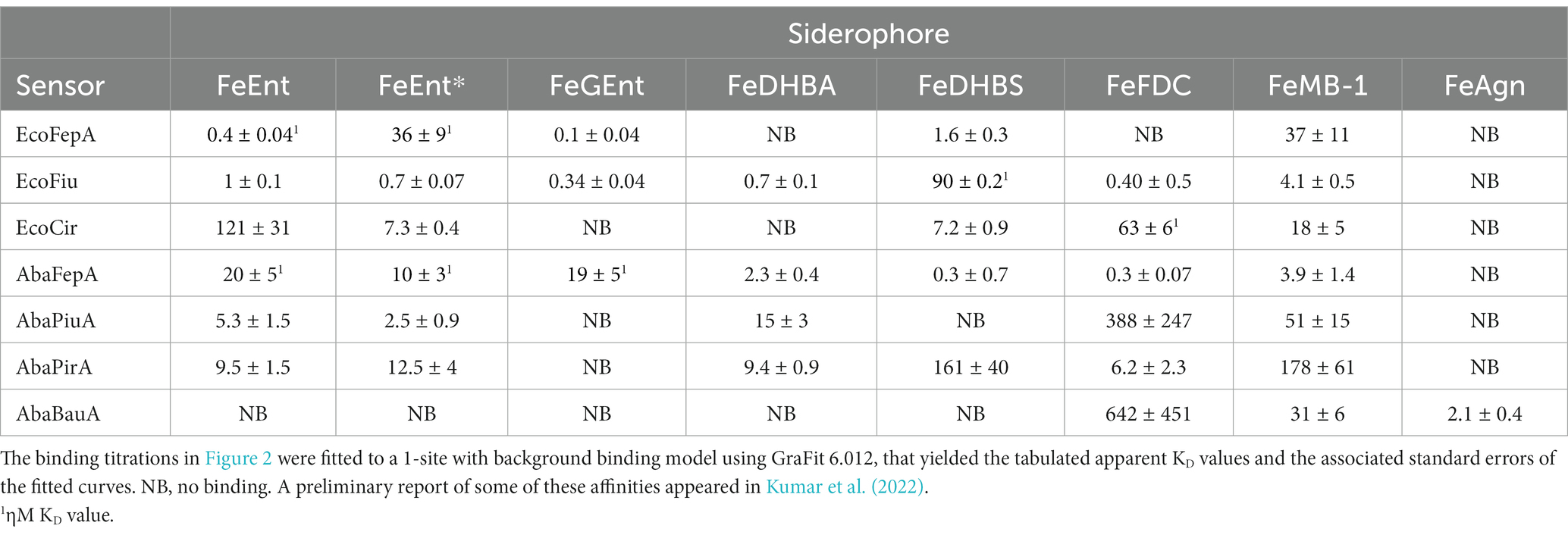
Table 2. E. coli and A. baumannii ferric catecholate receptor affinities (ηM or μM).
Escherichia coli ferric catecholate receptorsFiu_A694C-FM differentiated ferric catecholate siderophores with the following order of affinities: FeDHBS (KD = 0.1 μM) > FeGEnt (KD = 0.34 μM) > FeFDC (KD = 0.4 μM) > FeEnt* (KD = 0.74 μM) > FeEnt (KD = 1 μM) > FeMB-1 (KD = 4 μM). We did not observe significant quenching at site A694C-FM from the addition of FeDHBA, but Fiu_N554C-FM, which is closer to the binding pocket (as computed by our docking study; see below), was quenched by FeDHBA (KD = 0.7 μM). The defining aspect of Fiu’s binding activity was its ~1,000-fold lower affinity (micromolar KD values) for any of the ferric catecholates, relative to the high affinity of EcoFepA (nanomolar KD values) for FeEnt, FeEnt*, and FeGEnt (Figure 2).
Acinetobacter baumannii ferric catecholate receptorsThe susceptibility of A. baumannii to FeFDC (Zhanel et al., 2019; Delgado-Valverde et al., 2020; McCreary et al., 2021; Le et al., 2022) led us to investigate the recognition preferences of PiuA, AbaFepA, PirA, and BauA. The fluorescence spectroscopic binding determinations showed that the ligand preferences and affinities of these receptors for various ferric catecholate iron complexes were generally similar to those of their E. coli counterparts, with some exceptions.
AbaFepAThe first difference was that AbaFepA did not show the same high affinity for its ligand(s) that defines EcoFepA (48% sequence identity) and its close orthologs [i.e., KpnFepA and KpnIroN (Kumar et al., 2022)]. The ~10 nM KD value of AbaFepA for FeEnt indicated ~100-fold lower avidity for FeEnt than that of EcoFepA (0.1–0.3 nM; Newton et al., 1999; Chakravorty et al., 2019). Additionally, AbaFepA bound both FeEnt and FeEnt* with roughly the same affinity, whereas Eco FepA showed a strong preference (~100-fold) for FeEnt.
PiuAAs noted above, Fiu bound FeEnt and FeEnt* with micromolar affinities that were 100- to 1,000-fold weaker than the nanomolar affinity of EcoFepA for FeEnt. The A. baumannii ortholog of Fiu, PiuA (31% sequence identity), similarly adsorbed FeEnt and FeEnt* with lower (~1,000-fold) affinity than AbaFepA. Furthermore, overall, the affinity of PiuA for FeEnt and FeEnt* was ~10-fold lower than that of Fiu.
PirAAlthough they are 37% identical, the ligand selectivities of Cir and PirA differed in a few ways. The former preferentially bound FeEnt* and FeDHBS better than FeEnt, whereas the latter had comparable affinities for FeEnt, FeEnt*, FeDHBA, and FeFDC. Cir did not adsorb FeDHBA. Its most distinguishing attribute was its comparatively high affinity (KD = 60 nM) for FeFDC (Figure 2; Table 2).
Recognition of FeFDCAmong the E. coli receptors, Cir and Fiu showed the best recognition of FeFDC (KD values of O.06 and 0.5 uM, respectively; Figure 2; Table 2). The orthologs of A. baumannii bound FeFDC with lower affinity: AbaFepA (KD ~ 0.5 uM), PirA (KD ~ 5 uM), PiuA, and BauA (KD > 100 uM). Unexpectedly, both Cir and PirA preferentially recognized FeFDC and bound it more avidly than any other iron complex. BauA’s natural ligand is the mixed catecholate/hydroxamate ferric acinetobactin (FeAcn), but it still showed weak affinity for FeFDC and FeMB-1 (Figure 2; Table 2). In summary, the fluorescent binding titrations revealed both similarities and differences in the hierarchies of ferric catecholate recognition by the LGP of E. coli and A. baumannii that were consistent with the susceptibility of both organisms to FDC.
In silico predictions of ligand binding sites and molecular dynamics (MD) simulationsUsing AutoDock Vina (Trott and Olson, 2010; Eberhardt et al., 2021), we identified potential FeEnt* binding sites in E. coli Fiu (PDB 6BPM). This initial analysis found three binding sites: two surrounded by surface loops 6, 7, 8, and 9, and a third site deeper in the Fiu structure (Figure 3), in the interior of the β-barrel. Site 1 was situated in a vestibule cavity (Grinter and Lithgow, 2019a) bounded by the N-domain loop and external loops 7, 8, and 9. Site 2, located more centrally in the vestibule, adjoins the N-domain loop, beneath external loop 6. Site 3 sits beneath L3 and atop β-strands 5 and 6. AutoDock Vina identified the top 9 binding poses in each of the three sites, and for each site we chose the one with the highest score for further analysis (Figure 3). Another AutoDock Vina simulation using Fiu and FeDHBA (that Fiu does not transport) found two binding sites within Fiu that involved different residues and different ligand binding configurations from those that we identified (see Discussion section). The large internal cavity in Fiu, which houses site 3, was also apparent in its structural model (Grinter and Lithgow, 2019a).
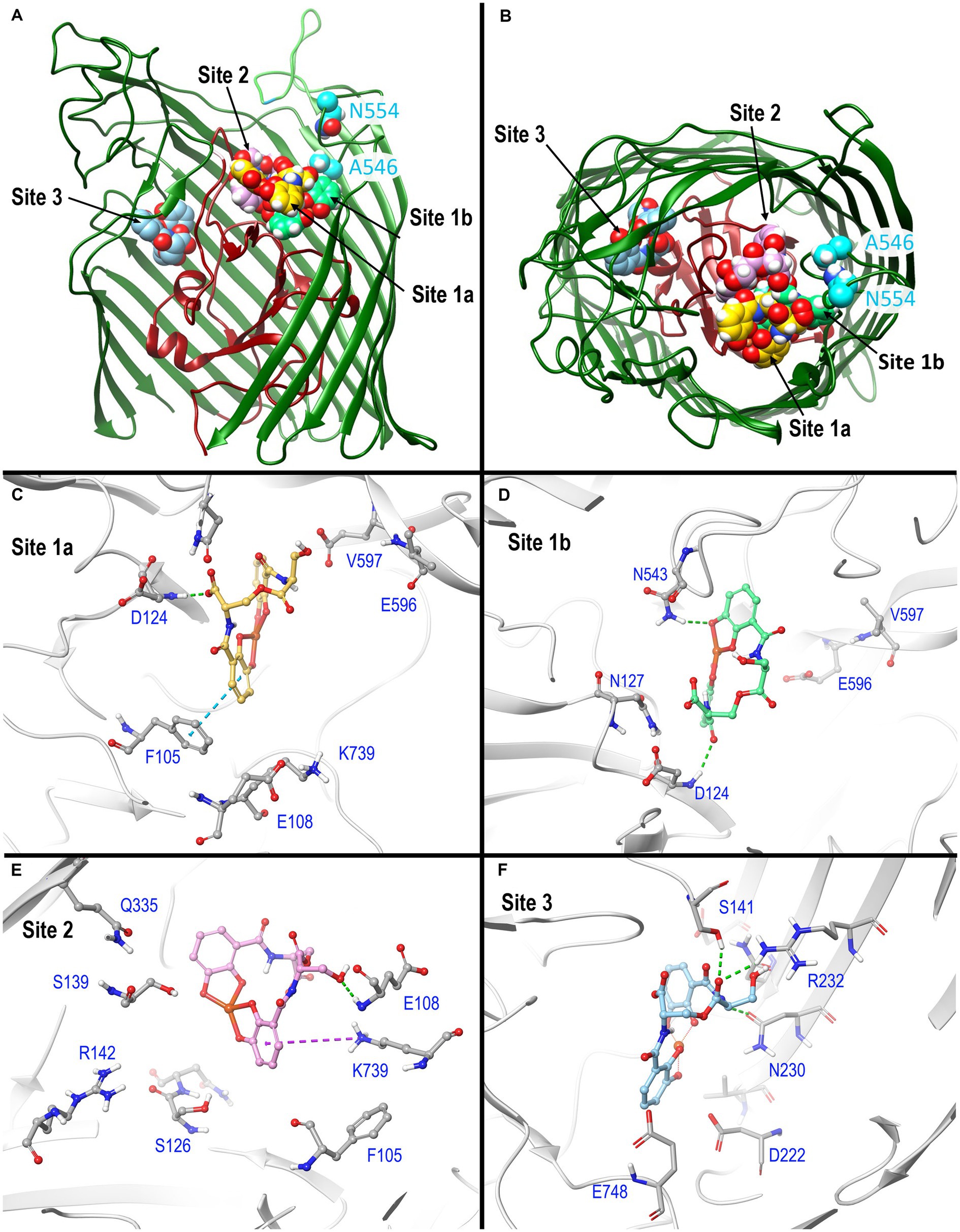
Figure 3. In silico docking/MD binding simulation. (A) AutoDock Vina Tool 1.5.6. paired Fiu (PDB 6BPM) and FeEnt*, to identify three outer binding sites (1a, 1b, and 2), and a fourth (site 3) deeper in the interior, within which the ferric siderophore is colored gold, spring green, orchid, and sky blue, respectively. We used the resulting docked model as the starting point for MD simulations that assessed the participation of individual amino acids in sites 1 and 2 during ligand recognition. In (A), the terminal 11 β-strands of the barrel were rendered transparent to reveal the ligand’s positions. (B) A 90 degree rotation on the x-axis shows a surface view of the receptor, and the binding positions of FeEnt*. The first N-domain (dark red) loop (NL1) bifurcates the interior cavity. We fluoresceinated the engineered residues N554C (cyan) near site 1a and A546C (cyan) near site 2 to study the binding reaction. (C–F) Detailed interactions of FeEnt* in sites 1a, 1b, 2, and 3, respectively (see text for detailed explanations).
After the initial identification of binding sites by AutoDock Vina, we used its resulting docked model as the starting point for MD simulations that assessed the participation of individual amino acids in sites 1 and 2 during ligand recognition. Over a duration of 500 ns, the binding interactions in site 1 were dynamic and variable, encompassing numerous positions of the iron complex, that bifurcated site 1 into sites 1a and 1b (see below) (Figure 3A). FeEnt* initially entered site 1a, near the conjunction of the first N-domain loop (NL1) and surface loop 9 (L9) of the β-barrel. The model predicted that in Site 1a, the negatively charged (−2), aromatic iron center of FeEnt* interacted with F105 (π-π stacking), D124 (H-bond), and K739 (ionic and cation-π bonds). In site 1a, FeEnt*was in proximity to NL1 (residues 119–123), L8 (residues 593–596), and L9 (residues 655–658) (Figure 3B). The siderophore’s four catecholate oxygens associated with Fe3+, along with two oxygens from extrinsic water molecules, creating an overall hexacoordinate metal complex (Figure 3).
After initial adsorption in site 1a, FeEnt* moved to site 1b, a binding pocket formed by residues 123–127 (NL1), 165–167 (NL2), and 538–544 (L7), wherein it hydrogen bonded to D124, N127, and N543. FeEnt* next occupied site 2, adjacent to and slightly beneath site 1, near the conjunction of the N-domain loop and L6. In site 2, the binding contacts were more stable. The simulations predicted interactions with F105 (π-π stacking), S126, N127, S139 (H-bonds), and K739 (cation-π interaction). The basic side chain of R142 was highly dynamic and only 4.3–9 Å away from the catecholate siderophore during the 500 ns simulation experiment. R142 may form a cation-π bond with the electron-rich aromatic ring, an ionic bond, or both, as the negatively charged ferric siderophore moves deeper in the beta-barrel. Residues 106–107 and 111–113 in NL1 were also in close contact with the ligand. When FeEnt* occupied binding site 2, it H-bonded to the amide nitrogen of E108, whose γ-carboxyl neared ionic bond distance to R486 in L6, that closed over the bound ligand (see below). Together with D124, E108 may play a key role in the translocalization of ligands from site 1a to site 1b and finally to site 2.
The movement of ligand from 1a to 1b occurred through two transition states (Supplementary Figure S4). Site 1a is ~10 Å removed from site 1b; the translocalization of FeEnt* between these sites is perhaps more accurately described as an induced fit of Fiu to capture the ferric siderophore from the environment.
Open and closed conformations of FiuIn addition to the predictions of FeEnt* binding sites, the MD simulations showed several configurations of surface loop 6 during the binding reaction (Figure 4). L6 was conspicuously open in sites 1a and 1b, but as FeEnt* progressed to site 2, L6 closed over the metal complex, with residue R486 dynamically moving toward N111, N544, or bound FeEnt*. The interactions between R486 in L6 and E108 in NL1 were particularly noteworthy because in site 2 the guanidino moiety of the R486 side chain approached the carboxylate of E108 to a distance of 5–6 Å, near salt bridge proximity. Grinter and Lithgow (2019a) reported two conformations of Fiu in the absence of a ligand that involved different forms of the N-domain loop. The changing surface loop configurations that we observed during the simulated Fiu-FeEnt* binding reaction were different, in that they encompassed the motion of L6 during ligand binding. These MD results are the first indication of loop motion in Fiu to enclose FeEnt*. However, in this respect, Fiu mimics the induced fit binding mechanisms of FepA (Scott et al., 2002; Smallwood et al., 2014) and FecA (Ferguson et al., 2002) as they close around their iron complexes. Hence, dynamic motion to capture ligands by induced fit may be a general property of LGP binding reactions.
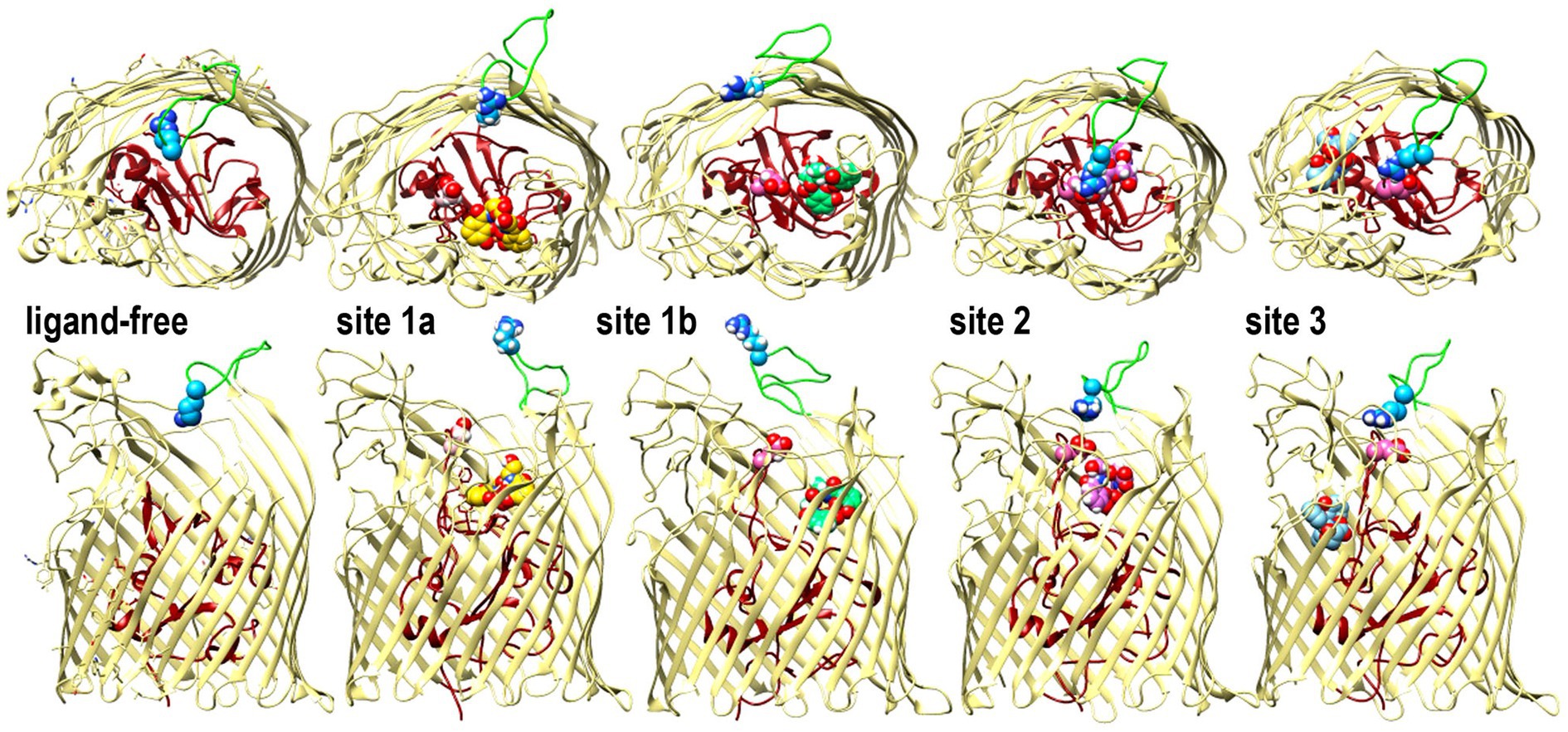
Figure 4. Open and closed conformations of Fiu. The images depict the predictions of MD simulations over 500 ns during FeEnt* binding to Fiu, as ribbon diagrams of Fiu either ligand-free, or associated with FeEnt* in sites 1a, 1b, 2, or 3. FeEnt* is colored gold, spring green, orchid, or sky blue, in sites 1a, 1b, 2, or 3, respectively. Surface loop conformations during the binding reaction are shown from above (top) or from the side (below); the N-domain is colored dark red. Although β-strands and other loops (khaki) remained relatively stationary, L6 (green loop) fluctuated among open conformations during initial FeEnt* binding to site 1a, but adopted a closed conformation above FeEnt* when the iron complex moved to site 1b. In the side view we rendered loops 3 and 10 transparent to better reveal sites 1a and 1b, see also Supplementary Video S1.
Biochemical analysis of predicted binding sites in FiuUsing fiu+ in the low-copy vector pITS23, we introduced Cys substitutions in L7 for fluoresceination, near binding sites 1a and 1b (FiuA546C) and 2 (FiuN554C). We then combined the Cys mutants with single or double Ala substitutions for residues of interest in sites 1 and 2. Our biochemical analyzes focused on sites 1a and 2 because site 3, deeper in the interior, was too distant from the L7 surface-localized fluorophores to permit analysis of its component residues by this approach. After confirmation of the mutations in sites 1 and 2 by DNA sequencing, we transformed the plasmids into E. coli OKN359 for evaluation in siderophore nutrition tests and into OKN1359 (ΔtonB, Δfiu, ΔfepA, Δcir) for fluorescent labeling and binding affinity determinations. We grew the resulting strains in iron-deficient MOPS media and subjected their engineered Cys side chains to chemical modification with FM. Fluorescence image analysis of SDS-PAGE gels of OM fractions from the mutants showed that all were expressed at or near wild-type levels and were well-labeled by FM (Supplementary Figure S3). The concentration-dependent fluorescence quenching of each construct by FeEnt* defined the binding affinities of the Fiu mutants. The experiments first showed that the Cys mutations near the EcoFiu binding pockets did not hinder the adsorption of FeEnt* (Table 3). Next, the individual Ala substitutions in binding site 1a behaved much like wild-type Fiu: They had slightly (~2-fold) lower affinity for FeEnt* than the positive control, N554C (KD = 0. 9 μM): F105A (KD = 1.2 μM); E108A (KD = 1.7 μM); D124A (KD = 2.5 μM); K739A (KD = 1.8 μM). The single mutants in binding site 2, however, were more impaired relative to the positive control A546C (KD = 1 μM). R142A showed the lowest affinity of FeEnt* binding (KD = 2.7 μM) of any single mutant. Other mutations in binding site 2 had smaller effects: S126A (KD = 1.9 μM), N127A (KD = 1.6 μM), S139A (KD = 1.7 μM), and Q335A (KD = 1.9 μM).
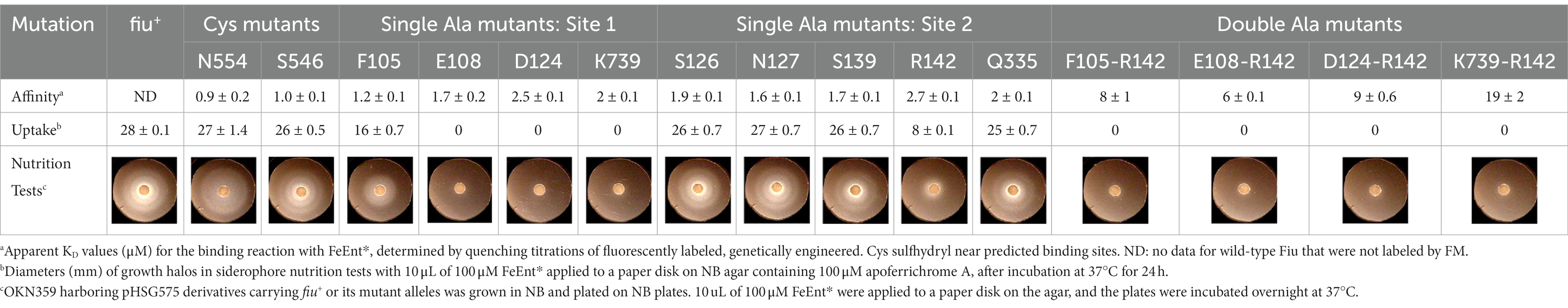
Table 3. Binding and uptake of FeEnt* by Fiu and its Cys or Ala substitution mutants.
Although single Ala substitutions in a multi-determinant binding pocket may only show small impacts on functionality, double substitutions for participant side chains often create larger reductions in ligand binding (Newton et al., 1997). Hence, we generated several double Ala substitutions, that combined single mutations in site 1a (F105A, E108A, D124A, K739A) with R142A, which created the most noticeable effects in site 2. After verifying the correct expression and fluorescence labeling of the double mutants (Supplementary Figure S3), we measured their binding affinities by fluorescence quenching assays. All four constructs had impaired binding relative to the positive controls and the single mutants, with a 7- to 22-fold increase in KD. The greatest decrease in affinity (22-fold) was from K739A-R142A (Figure 5; Table 3).
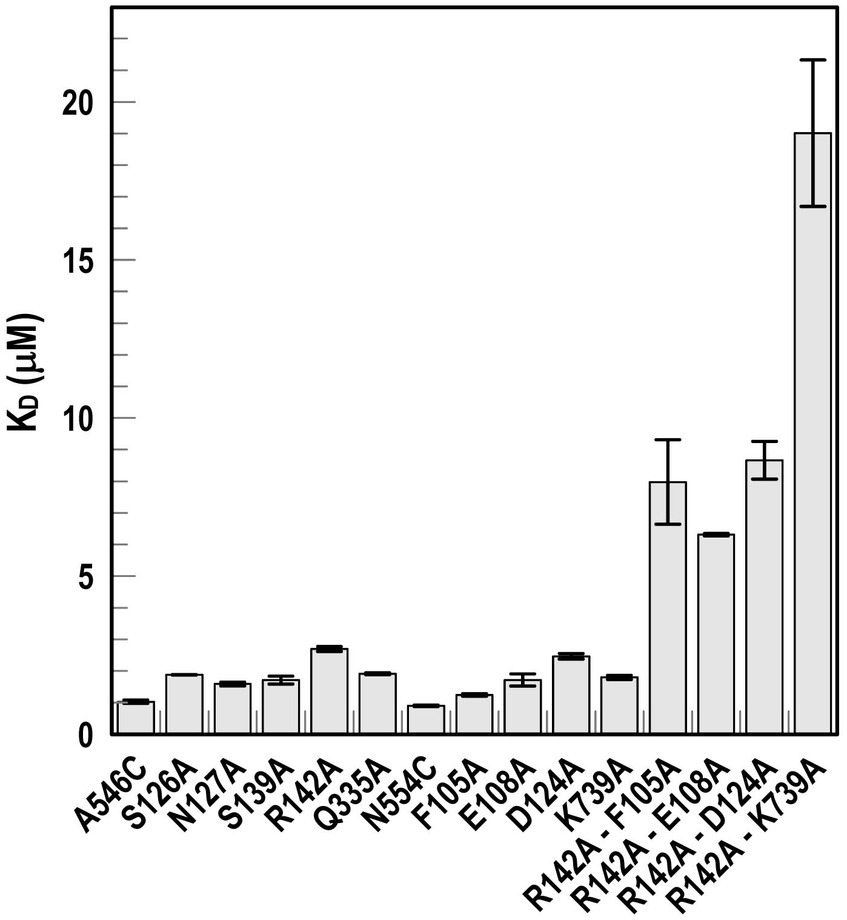
Figure 5. Ligand binding by Ala substitution mutants. In predicted binding sites 1a and 2, we engineered sensor residues A546C and N554C, respectively, and then engineered Ala substitutions for 9 other residues in the two binding cavities, as well as double mutation combinations. After the expression of the Fiu Cys-Ala mutants in OKN1359, we fluoresceinated the cells and determined their affinities for FeEnt* by fluorescence quenching.
Effects of Ala substitutions on FeEnt* uptakeIn addition to binding affinity titrations, we assessed the transport efficiencies of the Ala substitution mutants with siderophore nutrition tests. The experiments further illustrated the contributions of residues in sites 1a and 2. For example, despite the fact that none of the four single substitutions in site 1a substantially changed binding affinity (≤ 2-fold), three of those single mutants (E108A, D124A, and K739A) abolished the transport of FeEnt*. Even at 100 μM, FeEnt* did not produce growth halos on the lawns of these mutants. Similarly, F105A, also in site 1a, showed weaker growth than the wild-type control. The outcomes were similar but different in site 2. As in site 1 the single Ala substitutions only slightly decreased binding affinity (2- to 3-fold), and most had little impact on FeEnt* uptake: neither S126A, N127A, S139A, nor Q335A reduced growth on FeEnt* relative to the positive control FiuN554C. Only R142A in site 2 severely diminished the halo around FeEnt*. Overall, these data showed that changes in either predicted site could disrupt FeEnt* transport. Analysis of the double Ala substitution mutants supported this interpretation because all of the four site 1a–site 2 double mutants were completely defective in FeEnt* uptake. Thus, Ala substitutions in either site 1a or site 2 affected FeEnt* recognition, binding, and uptake.
Discussion Acquisition of ferric catecholatesThe findings clarify the transport attributes of E. coli Fiu, that was initially thought to be responsible for the uptake of FeDHBS, a monocatecholate degradation product of FeEnt (Hantke, 1990). Our results indicate that although Fiu adsorbs FeDHBA, FeDHBS, FeFDC, and FeMB-1, ferric monocatecholates are not its primary transport target. Rather, Fiu preferentially transported the dicatecholate degradation product, FeEnt*. Among numerous ferric catecholates that we tested, Fiu only transported FeEnt* and the siderophore antibiotic FeFDC. Cir had identical ligand specificity for FeEnt* and FeFDC. Aside from the initial reports of its uptake of FeDHBS (Hantke, 1990) and catecholate-β-lactam antibiotics (Nikaido and Rosenberg, 1990), the transport specificities of Cir were largely unexplored. Our data confirm its transport of FeDHBS and monocatecholate antibiotics (FeFDC); our findings additionally demonstrate, for the first time, its recognition and transport of FeEnt*. Despite its highest affinity and specificity for FeEnt, FepA also bound and transported both FeEnt* and FeDHBA, further illustrating the importance of these degradation products to overall iron acquisition. However, cells expressing only FepA were not susceptible to the monocatecholate FeFDC. These preferences of the three chromosomal ferric catecholate receptors of E. coli strategically balance to accommodate the tendency of FeEnt to decompose/degrade from the susceptibility of its lactone backbone to acid or base, or cellular hydrolases that cleave the lactone ring. The latter process sequentially produces linear trimeric and then dimeric ferric catecholate species that FepA, Fiu, and Cir recognize and transport. FepA is most important in the overall cell envelope transport system, because its higher affinity for both FeEnt and FeEnt* allows their uptake at even nanomolar concentrations. Fiu and Cir, on the other hand, add additional iron transport capability when ferric catecholates are present at much higher, micromolar levels.
Regarding the ferric catecholate recognition specificities of A. baumannii, relative to E. coli, genomic annotations from sequence identities among the various receptors generally agreed with their biochemical activities. That is, AbaFepA was most like EcoFepA, in sequence identity and its specificity for FeEnt, etc., with one exception: PirA was more identical to EcoFepA (56%) and AbaFepA (53%) than to Cir (36%), its purported ortholog (Supplementary Figure S6). This higher level of sequence identity between PirA and EcoFepA or AbaFepA inferred that its true ligand is FeEnt. However, the fluorescent binding titrations did not support this prediction: PirA showed no selectivity for FeEnt, but instead, similar micromolar affinity for FeEnt, FeEnt*, FeDHBA, and FeDHBS. Its affinity for FeEnt was 105- and 103-fold lower than that of EcoFepA and AbaFepA, respectively.
Biological relevance of Fiu, Cir, and FepA specificitiesIn response to iron deprivation, Fiu, FepA, and Cir are over-expressed to a level that is 10- to 20-fold higher than in iron-replete media, and the kinetics of their de-repression are virtually identical (Klebba et al., 1982). This coordinate, Fur-regulated enhancement of the three ferric catecholate transporters accompanies and mimics the Fur-regulated overproduction of Ent that also ensues from iron deprivation. The synchronous regulation of the Ent biosynthetic and the FeEnt/FeEnt* uptake systems concurs with our finding that all three receptors participate in the acquisition of FeEnt or its degradation products. However, despite their similar patterns of overproduction during low-iron stress, the different affinities of the three ferric catecholate receptors dictate their transport physiology: Only FepA (KDFeEnt = 0.4 nM) acquires FeEnt at low (nanomolar – micromolar) concentrations, but all three proteins may transport FeEnt* (FepA: KD = 40 nM; Fiu: KD = 0.7 uM; Cir: KD = 7 uM), which may accumulate at much higher (mM) concentrations during prolonged growth in iron-deficient conditions (Kumar et al., 2022) (Figure 6).
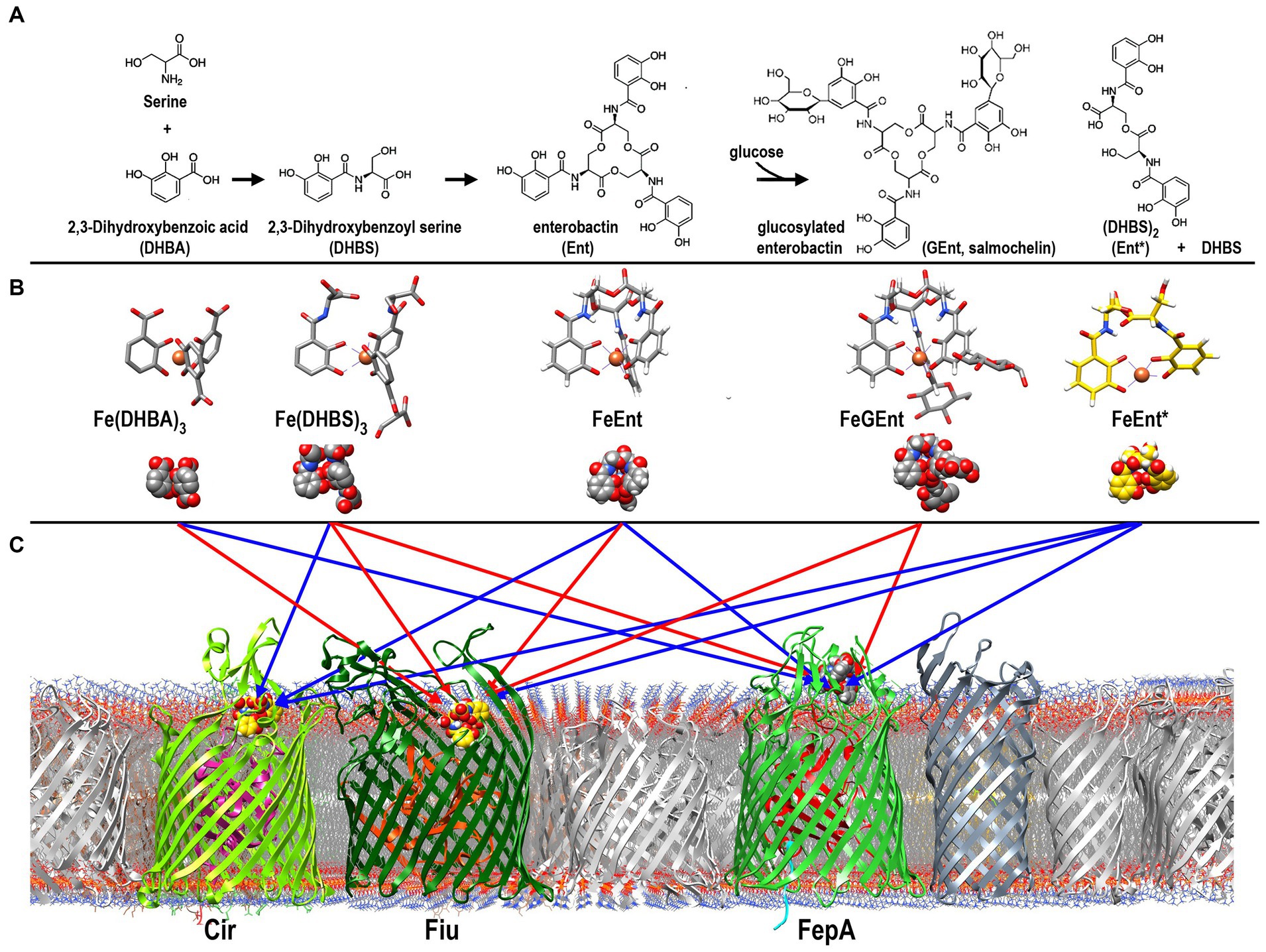
Figure 6. Pathways of ferric catecholate binding and transport in E. coli. (A). Catecholate siderophore biosynthesis and degradation. Ent is a trimer of DHBS, whose serine hydroxyl and α-carboxylate groups esterify to form a trilactone ring; the α-amino of Ser forms an amide bond with the carboxylate of DHBA. The resulting trimer may be glucosylated to form GEnt; both Ent and GEnt, and their iron complexes, may be degraded by acid, base, or esterases to form FeEnt* and DHBS. (B) Ferric complexes of catecholate siderophores. The monocatecholate biosynthetic and degradative byproducts, DHBA and DHBS, form relatively low affinity hexacoordinate, octahedral complexes with Fe3+. The tri- and dicatecholates Ent, Gent, and Ent*, on the other hand, form a higher affinity, hexacoordinate iron chelates. (C) Binding and uptake of ferric catecholates by Fiu, Cir, and FepA. The findings that we report established pathways of ferric catecholate binding only (red arrows), and binding followed by transport (blue arrows). In summary, Fiu only transported FeEnt*, but it also bound FeDHBA, FeDHBS, FeEnt, and FeGEnt; Cir transported FeEnt* and FeDHBS (weakly), but it also bound FeEnt; FepA transported FeDHBA, FeEnt, and FeEnt*, but it also bound FeGEnt, albeit with a much lower affinity.
Ligand recognition and movement of FeEnt* through FiuDespite its wide range of ferric catecholate recognition, Fiu only transported FeEnt*. The uptake of FeEnt*, but exclusion of FeEnt and Fe(DHBS)3, may likely derive from the larger size of the latter two iron complexes (555 vs. 719 and 774 Da, respectively). All three compounds manifest hydrophobicity from their peripheral aromatic rings and a negatively charged iron center (−2, −3, −3, respectively); the obvious difference among them is their overall size: FeEnt* is the smallest in mass and the most compact molecule. These considerations suggest that during uptake, FeEnt* travels a pathway that neither FeEnt nor Fe(DHBS)3 can follow, probably because of their larger size.
The identification of FeEnt* as Fiu’s natural ligand allowed AutoDock Vina to predict its binding interactions. The resulting docking model with FeEnt*, and ensuing MD simulations, showed binding positions in the Fiu vestibule and interior that delineated a putative pathway of ligand movement. These predictions had experimental support from the fact that site-directed Ala mutants in sites 1 and 2 impaired the binding and uptake of FeEnt*. The MD calculations further postulated the closing of L6 over FeEnt* when it moved to site 2 in Fiu, which mimics the two-stage kinetic model of FeEnt binding to FepA (Payne et al., 1997), and the closing of L7 in FecA during ferric citrate binding (Ferguson et al., 2002). Furthermore, when L6 changed position to cover FeEnt* in site 2, it was potentially stabilized by an ionic bond between R486 in L6 and E108 in NL1 (Figure 4). A similar phenomenon was observed in the FecA-ferric dicitrate crystal structure, as a salt bridge between residues R438 and E573 that covered the bound metal complex (Ferguson et al., 2002). In most circumstances, Fiu residues R142 and K739 have positively charged side chains that may form strong cation-π interactions with the ligand. This indicates the importance of amino acids with positively charged side chains and the associated cation-π bond in the bio-molecular complexing.
TonB-dependent transportThe process of metal transit through TonB-dependent LGP is not fully understood, but accumulating information, including current and prior findings about Fiu (Grinter and Lithgow, 2019a), suggests a mechanism. Conformational motion within the globular N-domain of EcoFepA is one aspect of the FeEnt uptake mechanism (Majumdar et al., 2020). As noted above, Fiu transported FeEnt* but not larger ferric catecholates that bound with about the same affinity. The AutoDock Vina and MD simulations projected a series of binding sites in the protein interior that ultimately position FeEnt* above a narrow opening between the N-domain and the β-barrel (Supplementary Video S1). In its initial state (Grinter and Lithgow, 2019a), this pore is too small to permit passage of FeEnt*, implying that the role of TonB activity is to transiently enlarge the channel, either allowing or compelling FeEnt* into the periplasm (Supplementary Video S1). In this context, the pathway enlarges enough to accommodate the transit of FeEnt* but not of FeEnt nor Fe(DHBS)3.
Most mechanistic studies of LGP-mediated ferric siderophore transport involved E. coli FepA (Newton et al., 1999; Scott et al., 2002; Cao et al., 2003; Chakraborty et al., 2003; Ma et al., 2007; Newton et al., 2010; Smallwood et al., 2014; Majumdar et al., 2020), FhuA (Carmel and Coulton, 1991; Braun et al., 1994; Bonhivers et al., 1996; Moeck et al., 1997; Endriss et al., 2003; Faraldo-Gómez et al., 2003; Eisenhauer et al., 2005), or BtuB (Hunter and Glass, 1982; Gudmundsdottir et al., 1989; Bradbeer and Gudmundsdottir, 1990; Fanucci et al., 2002; Cadieux et al., 2003; Chimento et al., 2003; Shultis et al., 2006; Nyenhuis et al., 2020), but the crystal structure of Fiu (Grinter and Lithgow, 2019a) opened new avenues to understanding this process. The uptake of Trojan horse antibiotics such as FDC by Fiu (this report; Ito et al., 2018, 2019) adds medical relevance to its recognition and transport properties. Previous research on FepA provides some insight into LGP transport biochemistry. FepA adsorbs FeEnt in a biphasic binding reaction (Payne et al., 1997) that involves induced fit by its surface loops around the ferric siderophore (Jiang et al., 1997; Scott et al., 2002; Smallwood et al., 2014). After reaching binding equilibrium in the surface vestibule, interactions between the TonB C-terminus and the FepA N-terminus induce a conformational change in the receptor that results in the internalization of the iron complex into the periplasm (Bradbeer, 1993; Newton et al., 2010; Klebba, 2016). Consistent with this idea, the MD-predicted sequence of binding sites through the Fiu interior suggests FeEnt* movement and ultimate deposition in a site where small changes in N-domain conformation may allow ligand transfer into the periplasm. In this sense, the MD simulations also fit with data on ferrichrome passage through FhuA (Eisenhauer et al., 2005) and FeEnt uptake through FepA (Majumdar et al., 2020), that implied conformational change in the N-terminal globule, while resident in the β-barrel, as part of the ligand uptake mechanism.
Previous findings also frame the actions of TonB-ExbBD in the uptake reaction: (i) The TonB C-terminus physically engages the N-terminal TonB-box sequence of ligand-bound LGP at the OM - periplasm interface (Pawelek et al., 2006; Shultis et al., 2006); (ii) TonB undergoes rapid, PMF-dependent motion, probably rotation in the IM bilayer (Jordan et al., 2013); (iii) ionic bonds restrict the N-terminus within the β-barrel, as a result of basic side chains in the N-domain that pair with acidic side chains on the interior of LGP β-barrels (Klebba, 2003). In Fiu, salt bridges exist between R121 – E570 and K154 - E617. These basic–acidic side-chain pairs are 90–95% conserved and evolutionarily covariant at comparable positions in dozens of other LGP (
留言 (0)