
記住我


The concept of Neuromodulation is defined by the International Neuromodulation Society (INS) as the use of advanced medical device technology to enhance or suppress the activity of the nervous system for the treatment of disease. This concept has evolved to “Bioelectronic Medicine” when using electrical current delivered to the nervous tissue (central or peripheral) to achieve targeted therapeutic benefits (Birmingham et al., 2014; Pavlov and Tracey, 2019, 2022; Cho et al., 2020). While this is a recent concept, early reports of the use of electricity as a therapy date back to thousands of years BC. The Ancient Egyptians and later the Greeks and Romans identified that electrical torpedo fishes generate electrical shocks that provide pain relief (Kane and Taub, 1975; Dolhem, 2008; Heidland et al., 2013). In the 18th and 19th centuries, these natural processes of electricity generation were replaced by man-made electrical devices. In the 18th century, Galvani’s momentum marked an era of discovery with his observations on frog’s leg contractions when applying electrical current over nerves or tissue (Kane and Taub, 1975; Loeb, 2014). Then, Benjamin Franklin led the generation of static electrical currents with his electrostatic machine and demonstrated the therapeutic effects of electroconvulsive shock therapy. This era was documented as Franklinism (Beaudreau and Finger, 2006; Finger, 2006; Finger and Zaromb, 2006).
The 19th century was the “golden age” of electrotherapy. In 1831, Michael Faraday fabricated the first electric generator apparatus, that produced intermittent current. Then Wilhelm Holtz developed the Holtz machine, a generator of static electrical current, which was implemented in medical practice to relieve pain and migraine (Al-Khalili, 2015). At the end of the 19th century, electricity was used for countless dental, neurological, psychiatric and gynecological conditions. However, at the beginning of the 20th century, electrotherapy lacking a scientific basis lost credibility. Furthermore, the development of effective analgesic drugs decreased the interest in electricity (Fodstad and Hariz, 2007; Dolhem, 2008). In the second half of the 20th century, electrotherapy underwent a revival. Based on animal experiments and clinical investigations, its neurophysiological mechanisms were elucidated in more detail. The pain-relieving action of electricity was explained in particular by two main mechanisms: first, segmental inhibition of pain signals to the brain in the dorsal horn of the spinal cord and second, activation of the descending inhibitory pathway with enhanced release of endogenous opioids and other neurochemical compounds (e.g., serotonin, noradrenaline, gamma aminobutyric acid, acetylcholine and adenosine) (Dolhem, 2008; Vallejo et al., 2017; Chakravarthy et al., 2018).
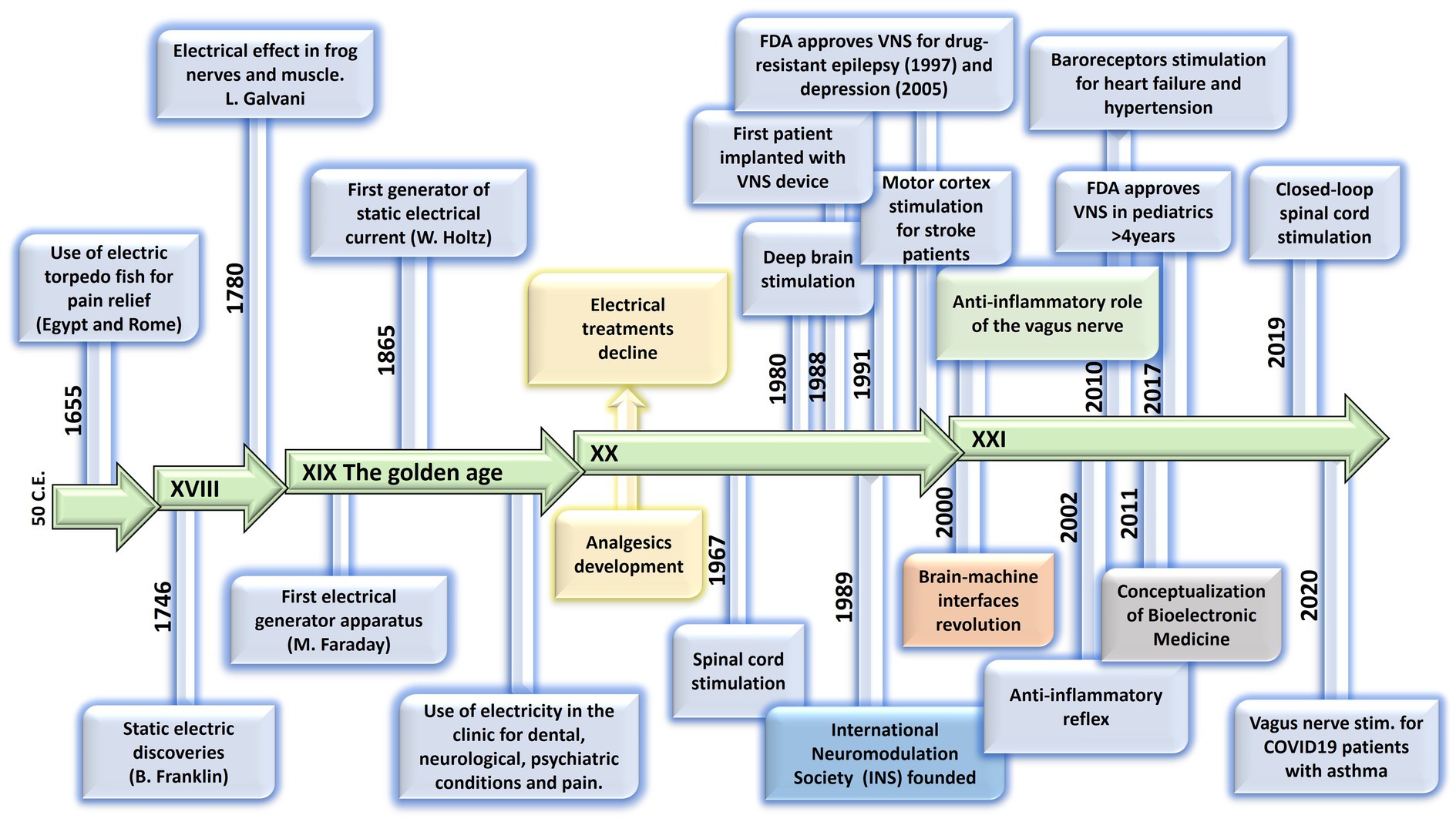
The notion of using the stimulation of large nerve fibers in the dorsal columns of the spinal cord for addressing chronic pain emerged during the 1960s (Shealy et al., 1967). This idea reached its pinnacle in 1968 with the development of the first commercial implantable stimulator designed for managing chronic pain and laid the foundation for modern neuromodulation techniques (Gildenberg, 2006). Another milestone in the neuromodulation field was the development of deep brain stimulation (DBS). While first used for the study of the physical manifestations of human emotion (Oliveria, 2018), it was when used for stimulation of the basal ganglia to improve symptoms of Parkinson’s disease that revolutionized the field evolving afterwards into a versatile therapy with applications in various neurological and psychiatric conditions (Frey et al., 2022). It was in the late 1980s that modern DBS techniques gained attention, with the advent of precise electrode placement and sophisticated neuroimaging methods. By delivering controlled electrical impulses, DBS modulates aberrant neural activity, effectively alleviating symptoms in conditions such as Parkinson’s disease, essential tremor, dystonia, epilepsy, obsessive-compulsive disorder, and even depression and schizophrenia. Ongoing advancements in electrode design, longer-lasting batteries, programming algorithms for closed-loop approaches, and the understanding of neural circuitry help refine and expand the therapeutic possibilities of DBS, offering new avenues of hope for individuals grappling with complex neurological and psychiatric disorders (Frey et al., 2022), and highlighting the necessity of interdisciplinary efforts. In 1989 the INS was funded in order to promote the dialog between clinicians, scientists and engineers (INS, 2022), and since then, national neuromodulation society chapters have been established in more than twenty countries around the globe, who meet in annual global and regional meetings to discuss advances in the field. Figure 1 summarizes the main events in the history of Bioelectronic Medicine.
Figure 1. Evolution of electrical therapies. Timeline highlighting the main historic events and evolution of the use of electricity for neuromodulation therapies. XIX century is recognized as the golden age due to impactful discoveries in electricity, including the first electrical generator apparatus (by M. Faraday) and the first generator of static electrical current (by W. Holtz). At XX century, the use of electrical therapies declined and lost credibility due to the lack of scientific basis, and to the development of the pharmaceutic industry (incorporation of analgesics). A new era begins at XXI century with the brain-machine interfaces and the discovery of the neuroinflammatory reflex.
An important advancement in the bioelectronic field was the implementation of vagus nerve stimulation (VNS). The vagus nerve is the main autonomic nerve communicating the brain with peripheral organs (Baker and Lui, 2019). The approval of VNS for drug-resistant epilepsy in 1997 and subsequently for treatment-resistant depression in 2005 stands out as important milestones within the bioelectronics field (Johnson and Wilson, 2018; Austelle et al., 2022). In the USA, the green light from regulatory authorities FDA for VNS therapy in these conditions has validated the safety and efficacy of bioelectronic interventions. However, the noncoverage determination in 2006 by the Centers for Medicare and Medicaid Services (CMS) that the VNS for depression remained unproven has limited its use (Aaronson and Conway, 2018). More recently, the discovery of transcutaneous auricular vagus nerve stimulation (tVNS) as a viable treatment option for epilepsy and depression carries immense importance for the treatment of these conditions. This innovation not only expands the range of available treatments but also opens the doors for a wider patient population to access therapy and eliminates the need for invasive procedures, reducing potential risks and improving patient comfort and compliance (Verma et al., 2021). It is worth mentioning the limitations of this approach, a customized treatment is required, and often side effects arise due to the off-target stimulation −undesired nerve fibers are recruited (Kim et al., 2022). Section 12 will provide more details on this.
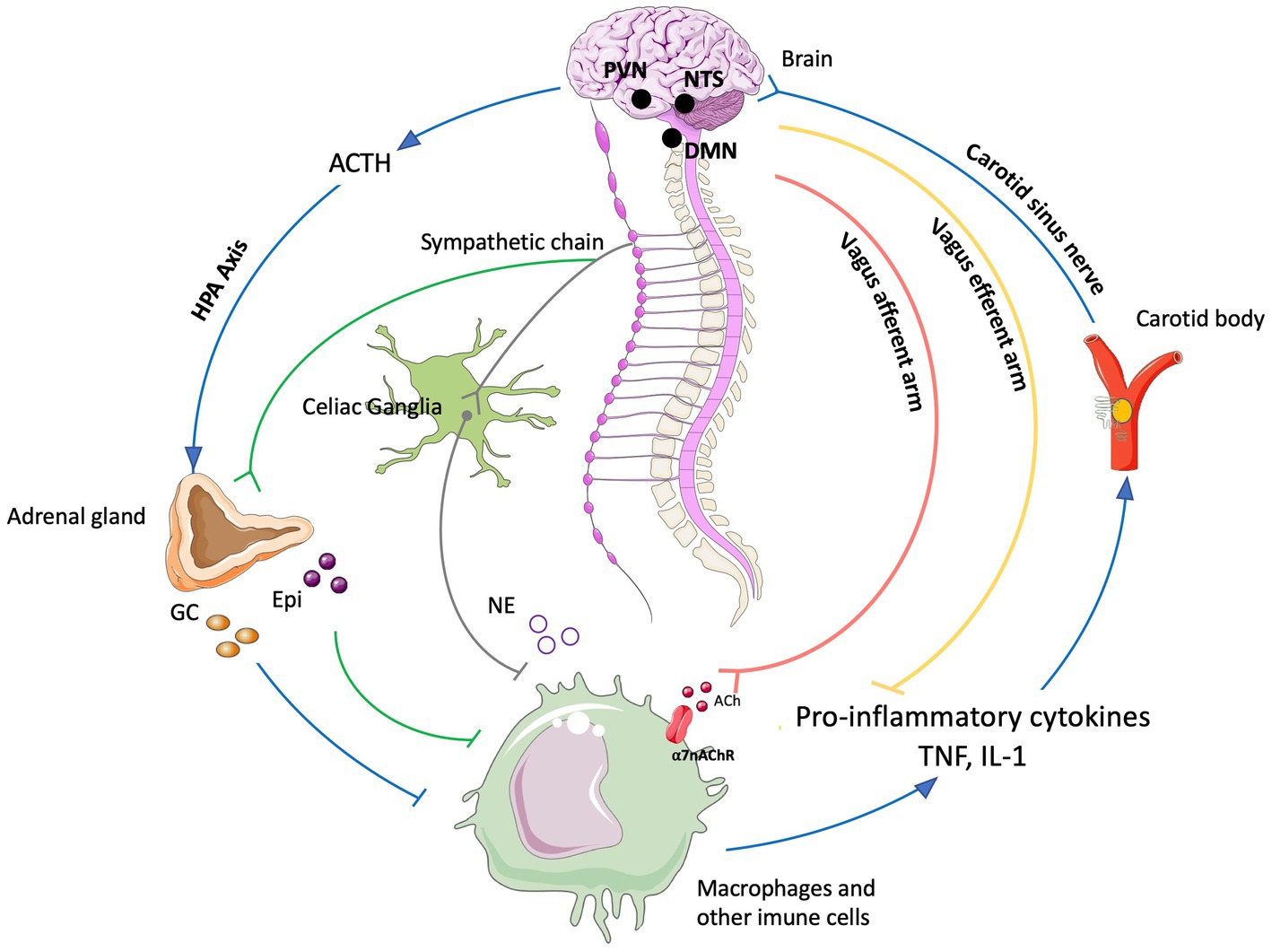
The last decades have been fruitful in the identification of new targets for the treatment of a variety of pathologies using Bioelectronic Medicine, particularly on the peripheral nervous system (PNS). One example is the neuro-immune reflex and its relevance in chronic inflammatory and immune diseases (Eberhardson et al., 2020; Pavlov et al., 2020). The mechanisms behind the neuro-immune reflex have been under debate, but the clinical output has marked a significant turning point in the Bioelectronic Medicine field (Tracey, 2002; Conde et al., 2020; Hu et al., 2020; Pavlov et al., 2020; Gonzalez-Gonzalez et al., 2021; Figure 2). Since pioneer studies in 2000, showed the decrease in inflammation followed by VNS in rats (Borovikova et al., 2000), multiple studies in animal models were performed showing that VNS decreases inflammation and mitigates disease in acute inflammatory conditions such as endotoxemia, sepsis, hemorrhagic shock, postoperative ileus, and kidney ischemia–reperfusion injury [for a review see Kelly et al., 2022] and in chronic inflammatory conditions such as rheumatoid arthritis, inflammatory bowel disease (IBD) and asthma (Levine et al., 2014; Yuan and Silberstein, 2016; Wheless et al., 2018; Austelle et al., 2022). The mechanisms have been under debate and accepted statements include the regulation of cytokine production through the nicotinic acetylcholine receptor subunit α7 (α7nAchR) in the spleen, termed “the cholinergic anti-inflammatory pathway” (Wang et al., 2003; Gallowitsch-Puerta and Pavlov, 2007; Parrish et al., 2008), through second-order vagal projections (indirect) (Ackerman et al., 1987; Berthoud and Powley, 1993) and first-order (direct pathway) vagus nerve-spleen (Gonzalez-Gonzalez et al., 2021). Another important landmark for the bioelectronic targeting of the vagal anti-inflammatory reflex has been the clinical testing of VNS devices for patients with inflammatory and autoimmune disorders, such as rheumatoid arthritis and IBD, with decreased inflammation and positive clinical outcomes (Koopman et al., 2016; Genovese et al., 2020).
Figure 2. The neuro-immune reflex. The activation of the hypothalamic-pituitary-adrenal (HPA) axis is prompted by various stimuli, initiating a sequence that involves the paraventricular nucleus (PVN) of the hypothalamus. This, in turn, triggers the release of cortisol releasing hormone (CRH) into the anterior pituitary. Consequently, CRH prompts the secretion of adrenocorticotrophic hormone (ACTH) into the bloodstream. ACTH stimulates the adrenal cortex to produce glucocorticoids (GC), which are potent anti-inflammatory molecules. These effects are mediated by the glucocorticoid receptor (GR), a nuclear receptor present in almost all cells, particularly innate immune cells. The vagal anti-inflammatory reflex is characterized by peripheral vagal afferent nerves sensing inflammation and conveying this information to the central nervous system (CNS). The nucleus tractus solitary (NTS) and dorsal motor nucleus (DMN) establish connections to communicate and activate efferent vagus nerve fibers. This activation leads to the release of acetylcholine (ACh), which diminishes cytokine production and inflammation. A key mechanism in this process is the binding of ACh to the nicotinic α7nAChR receptor. Furthermore, cytokines can directly activate this specific brain region through humoral control to counteract inflammation. Sympathetic fibers originating in the spinal cord directly innervate visceral organs and immune cells. Upon binding to their receptors, norepinephrine (NE) released from sympathetic fibers and at the celiac ganglia, as well as epinephrine (Epi) from adrenal chromaffin cells, collectively inhibit inflammation and the release of inflammatory cytokines. Additionally, inflammation resolution is facilitated by the actions of the carotid body. Inflammatory mediators released by immune cells trigger the activation of the carotid body, which subsequently stimulates the carotid sinus nerve. This nerve projects into the NTS, contributing to the overall process of inflammation resolution Created with BioRender.com.
Exploring the neuro-immune reflex, beyond the vagus nerve, the electrical modulation of carotid body chemoreceptors also modifies the inflammatory response during sepsis. Previous studies showed in conscious rats, that the electrical stimulation of the carotid sinus nerve, the sensitive nerve of the carotid body, attenuates the response to bacterial lipopolysaccharide (LPS) by decreasing tumor necrosis factor (TNF), interleukin (IL) 1β (IL-1β) and IL-6 levels, and increasing the levels of IL-10. These effects were mediated by both sympathetic and parasympathetic pathways, and therefore consistent with the activation of both afferent and efferent pathways of the anti-inflammatory reflex (Santos-Almeida et al., 2017). In addition, it was shown that the electrostimulation of the carotid sinus nerve in mice attenuates inflammation via glucocorticoid receptors on myeloid immune cells (Falvey et al., 2020). In 2020, a handheld VNS device was authorized by the U.S. FDA as an emergency for asthmatics with COVID-19 experiencing difficulty breathing (ClinicalTrials.gov; identifier: NCT04368156), and the consecutive randomized clinical trial demonstrated potential to mitigate some of the symptoms by reduction of inflammatory markers (Tornero et al., 2022).
The notion of joining forces to accelerate discoveries for Bioelectronic Medicine, led in 2022 to celebrate the 5th Bioelectronic Medicine Summit in NY city, where an update of recent advances was discussed (Ashe et al., 2023). These include the use of targeted ultrasound stimulation, the implementation of solid-state batteries for the miniaturization of medical devices, advances in closed-loop neuromodulation and multi-contact implantable neuronal interfaces. While we are in a revolutionary technological era, multidisciplinary dialog is imperative, including but not limited to clinicians, physiologists, data analysts, and material/electronic engineers. We must keep in mind that neuromodulation goes far beyond VNS, DBS, or spinal cord stimulation. Countless opportunities arise from the discovery of new homeostatic neural mechanisms, and more significantly, from innovative mechanisms and points of intervention. Furthermore, many of the previously identified targets, such as nerves and neuronal regions, have the potential to serve as neuromodulation targets for multiple diseases, emphasizing the importance of acquiring further insights into comprehensive neuroanatomy, neurophysiology, and biophysics needed for the intelligent design of device hardware computational models and stimulation protocols.
The present work provides and discusses core concepts required to understand the Bioelectronic Medicine field. Experts will go from the inner core of neuromodulation to the patient point of care, passing through the current neuromodulation approaches and neural interfaces available and their applications, which altogether stresses the necessity of multidisciplinary dialog to advance this field.
2 Biophysics of the neuronal membrane: the inner core of neuromodulation (R. Latorre)Neuronal membrane excitability is a fundamental property underlying function in the nervous system and it depends on a cascade of events followed by a stimulus. In electrical therapy, an external stimulus modifies the neuronal dynamics, and it is necessary to understand the underlying cellular events in order to develop high-precision treatments. This section reviews fundamental concepts of biophysics related to the neuronal membrane, focusing on the main properties of voltage-gated (dependent) channels involved in the adequate functioning of the nervous system.
2.1 How voltage-dependent channels are activated by voltageSince the advent of the patch-clamp technique, followed by ion channel cloning, and ending with the flourishing of structural methods (such as X-ray and cryo-electron microscopy), we have witnessed how the ion channel field has advanced with amazing speed. It would be impossible to examine in detail all the gating and structural properties of the myriad voltage-dependent channels present in the nervous system. Therefore, a bird’s-eye view will be provided to the main biophysical properties.
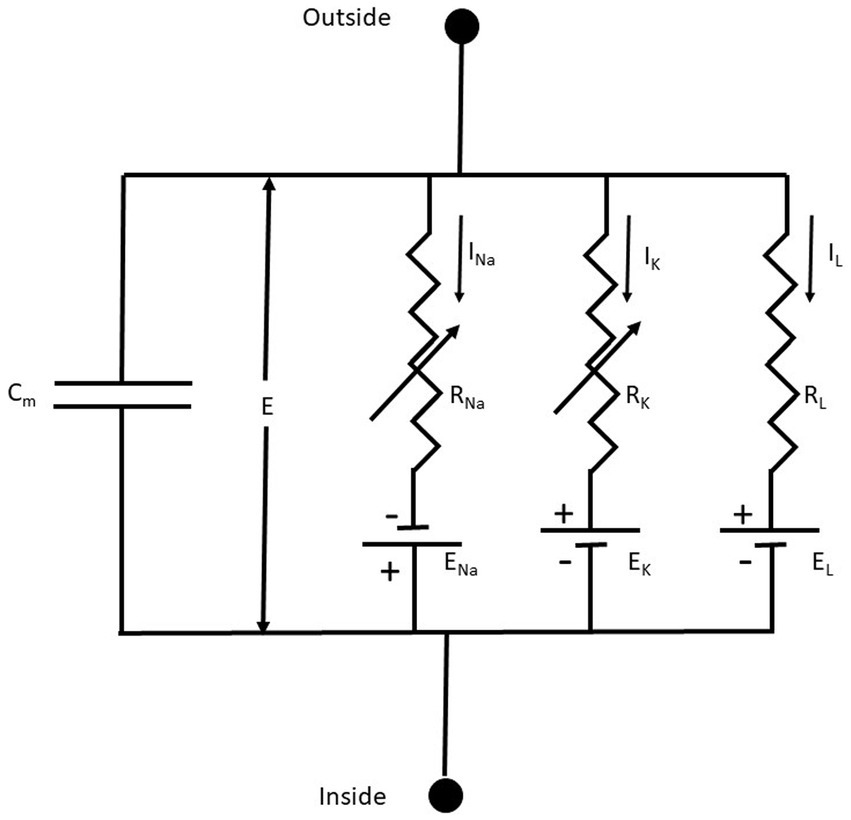
2.1.1 Membrane equivalent circuitAn excellent starting point to discuss the membrane’s electrical properties is the 1952 paper of Hodgkin and Huxley (HH) (Hodgkin and Huxley, 1952), who summarized their previous findings by representing the Na+, K+, and leak (L) conductance in an equivalent membrane electrical circuit (Figure 3).
Figure 3. The equivalent membrane electrical circuit. CM is the membrane capacitor; E is the membrane potential; ENa, EK, and EL are the equilibrium potentials for Na+, K+, and L (leakage), respectively. RNa, RK, and RL, are the membrane resistances for Na+, K+, and L, respectively. Notice that RNa, RK are variable membrane resistances. The currents IX = GX(E – EX), where X = Na+, K+ or L and the conductance GX = 1/RX. From Hodgkin and Huxley (1952), Figure 1.
The membrane conductances for Na+ and K+ (GNa and GK, respectively), are variable and are a function of the voltage-dependent membrane potential and time. The leakage conductance (GL) is constant. Thus, according to the present electrophysiological convention, we have a negative inward (IN) current when the membrane potential I < ENa, which becomes positive when E > ENa. Thus, in this seminal work, for the first time is used the concept of voltage-dependent conductances. The probability of finding the sodium or the potassium conductance operative is a function of voltage. GNa and GK increase with membrane depolarizations (i.e., membrane potentials larger than the resting membrane potential).
Hodgkin and Katz (1949), following the lead of Goldman (1943), obtained an expression for the membrane potential (Em) that takes into account internal and external concentrations of K+, Na+, and Cl- and the permeability coefficients (P) for the different ions. This equation, known as the Goldman-Hodgkin-Katz or constant-field equation, considers the non-equilibrium conditions of the cell system and is still used when describing the ion selectivity of a given conductance system (Alvarez and Latorre, 2017).
Em=−RTFlnPKKin+PNaNain+PClCloutPKKout+PNaNaout+PClClin (1)The use of Equation (1) allowed Hodgkin and Katz to determine the relative permeabilities of K+ and Na+ during rest and at the peak of the action potential. At rest, PK/PNa = 25, and at the peak of the action potential, PK/PNa = 0.05, implying that during an action potential PNa increases 500 times compared with that at rest.
2.1.2 Voltage-dependent conductancesVoltage-dependent channels are constituted by proteic subunits embedded in the cellular membrane, and every subunit is formed by six transmembrane segments (S1-6). These subunits form a central pore that allows the flow of ions, and its opening is triggered by stoichiometric modifications activated by changes in charges. The S4 transmembrane segment in voltage-dependent channels contains the gating charges. HH were fully aware that in order to have a voltage-dependent conductance, charges or dipoles contained in the plasma membrane should interact with the electric field (Hodgkin and Huxley, 1952). The movement of these charged particles (voltage sensors) should induce a current (gating currents), and they were revealed by Armstrong and Bezanilla (1973). The cloning of the voltage-dependent Na+ (Nav) and the Shaker K+ channel (Noda et al., 1984; Papazian et al., 1987; Timpe et al., 1988) gave the first hints about the molecular nature of the voltage sensor. Nav channels are formed by a protein containing four domains, each containing six transmembrane segments (S1-S6). Voltage-dependent K+ (Kv) channels are tetramers; each subunit has six transmembrane segments. Notably, in Nav and Kv channels, in the fourth S4 segment, every third residue is a positively charged amino acid residue (arginine or lysine) separated by two hydrophobic residues. Using the limiting slope method (Almers, 1978; Sigg and Bezanilla, 1997) and the total gating charge (Q) divided by the number of channels (N) contained in the same membrane area (Q/N method), it was possible to obtain the number of gating charges per channel. Twelve electronic charges (e0) are needed to be displaced in the electric field to open Nav channels from skeletal muscle (Hirschberg et al., 1995), and 12.3 e0 were obtained in the case of the Shaker K+ channel (Schoppa et al., 1992). Using these methodologies to determine the numbers of gating charges per channel and replacing the charged amino acids contained in S4 for the neutral glutamine or asparagine, Aggarwal and MacKinnon (1996) and Seoh et al. (1996) demonstrated that the first four charges from the N-terminal of S4 contribute to the Shaker K+ channel gating charge.
On the other hand, state-dependent accessibility of cysteines replacing some of the gating charges and using cysteine-modifying reagents showed that the S4 segment moves upwards, exposing the charged residues to the external milieu (Yang and Horn, 1995; Larsson et al., 1996). Given the origin of what is now known as the voltage-clamp fluorometry technique, Mannuzzu et al. (1996) modified the S4-cysteines mutants with fluorescent probes showing voltage-dependent fluorescent changes that parallel the voltage dependence of the gating charge displacement. These experiments provided physical evidence that the S4 is the voltage sensor.
Gating charges move through a thin hydrophobic septum at the cell membrane. Testing the voltage-dependent accessibility of the S4 residues in Nav channels, Yang and Horn (1995) found that two of the S4 charges were accessible from the inside at hyperpolarizing voltages and accessible from the outside in response to depolarization. They moved across the whole electric field, transferring 1 e0 each. However, the following charge was only accessible from the inside regardless of the applied voltage. A parsimonious explanation for these results is that during channel activation, the S4 charges move through a thin hydrophobic septum of about 10 Å (Yang et al., 1996). The idea that the S4 is displaced between water-lined crevices separated by a thin septum was corroborated by replacing the S4 charges for histidine residues and probing the state-dependent accessibility to proton in a proton gradient (Starace and Bezanilla, 2001; Starace and Bezanilla, 2004). Defining the four arginines that contribute to the total Shaker K+ channel gating charge as R1-R4, Bezanilla’s laboratory found that when mutated to histidines, R2 and R3 behave as proton carriers implying that they move across the entire length of the electric field. Remarkably, when a histidine replaces R1, a voltage-dependent inward rectifier proton pore is produced (i.e., it conducts protons only when voltage sensors are at rest). The R4H also form a proton channel, but it conducts only when sensors are in the active position (Starace and Bezanilla, 2001; Starace and Bezanilla, 2004). Thus, all R1-R4 move from the internal to the external water-lined crevice during Shaker K+ channel gating, strongly suggesting that, as in Nav, the crevices are separated by a narrow division. Moreover, structural and electrophysiological studies indicate that the electric field is focused in a hydrophobic plug or charge transfer center of about 10 Å in thickness (Chen et al., 2010; Tao et al., 2010; Lacroix and Bezanilla, 2011). Using a fluorescent positively charged bimane derivative (qBBr) strongly quenched by tryptophan, Priest et al., (2021) show that charge displacement consists of rotation and a tilted translation during activation, but the detailed movement differs for R1 and R2.
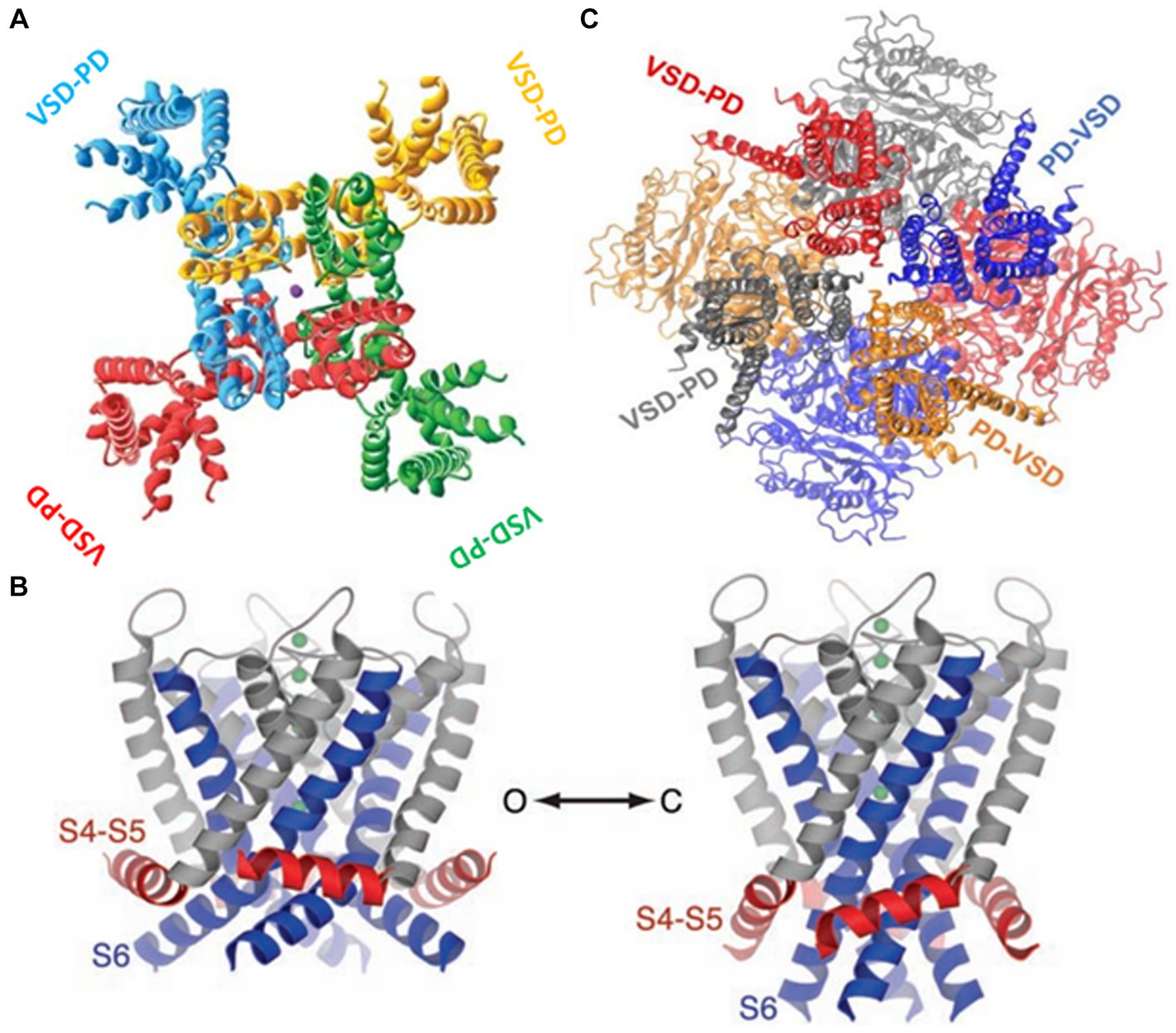
2.1.3 Electromechanical couplingNow, we turn to the question of how the electrical energy contained in the displacement of the S4 is transformed into mechanical energy, leading to the pore (or ion channel) opening. The structure of Kv in its open-inactivated conformation (Long et al., 2005a) and Nav (Shen et al., 2017) channels show they are formed by modular proteins. Kv channels are tetramers, and the Nav protein-forming channel consists of four (I-IV) domains. Each subunit in Kv or domain in Nav consist in a voltage sensor domain (VSD; S1-S4) and a pore domain (PD; S5-S6) (Figure 4). The structure of the Kv1.2 shows the VSD are in a swapped configuration (Figure 4A). The VSD makes contact with the pore domain of an adjacent subunit (domain-swapped channels). Although the structure of the closed Kv channel has not been elucidated, all the results to date indicate that the four S6 transmembrane segments form a bundle crossing that hinders the passage of ions. The question is how this bundle separates to allow ion conduction during activation. The first indications regarding the coupling between the VSD and the pore opening were given by the group of Lu et al. (2001) and Lu et al. (2002), who demonstrated that the S4-S5 linker interacts with the C-terminal of S6. The structure of the Kv1.2/Kv2.1 (Long et al., 2007) showed that C-terminal of S6 contacts the S4-S5 linker N-terminal of the same subunit and the C-terminal of the S4-S5 linker of the S6 of the adjacent subunit. Since only the open Kv structure has been determined, we start with the S6 bundle crossing in its open configuration. When the S4 moves downward upon closing, the S4-S5 linker pushes the S6 toward the closed configuration of the gate (Long et al., 2005b, 2007; Jensen et al., 2012; Yarov-Yarovoy et al., 2012; Figure 4B). Above, we have described the canonical electromechanical coupling. However, several recent reports provide evidence that the interactions of amino acid residues at the interface of VSD and the pore domain contribute to electromechanical coupling in voltage-dependent channels (Fernández-Mariño et al., 2018; Kalstrup and Blunck, 2018; Carvalho-de-Souza and Bezanilla, 2019; Bassetto et al., 2021). This non-canonical coupling can play an essential role in those voltage-dependent channels lacking a well-defined S4-S5 and, particularly in those channels in which the VSD is adjacent to the P-domain of the same subunit (non-domain swapped channels). This is the case of the voltage-gated potassium channels (KCNH) family (Whicher and MacKinnon, 2016; de la Peña et al., 2018; Cowgill and Chanda, 2021) and the Ca2+ and voltage-activated (BK) channels (Hite et al., 2017; Tao et al., 2017; Carrasquel-Ursulaez et al., 2022; Figure 4C).
Figure 4. (A) The swapped configuration of the KV 1,2 channel. Each subunit is represented in a different color. The sphere in the center is a potassium ion ([2]PDB ID 2A79). (B) The non-swapped configuration of the BK channel. Notice the VSD is contacting the pore of the same subunit. The c terminals in the background are shown forming the gating ring [3]. (C) Left. The open structure of the K1.2 channel. S6 is showed in blue and the S4-S5 linker in red. Right. A model for the closed structure of the Kv1.2 channel (Long et al., 2005b).
2.1.4 Voltage-dependent ion channels and neuromodulationDifferent types of Nav channels are found in the nervous system (Nav1.1-Nav1.9) coded by nine genes (SCNA1A-SCNA5A; SCNA8A-SCNA11A) (Cox et al., 2006). Nav channels can be distinguished by their sensitivity to tetrodotoxin (TTX). Nanomolar concentrations of TTX block Nav1.1-Nav1.4, Nav1.6, and Nav1.7, whereas Nav1.5 (cardiac Nav), Nav1.8, and Nav1.9 are TTX-resistant channels. Nav1.3, Nav1.7, Nav1.8, and Nav1.9 are involved in nociception in sensory neurons (Cox et al., 2006). Loss-of-function mutations of Nav1.7 have been found in families with congenital insensitivity to pain (Kaplan et al., 2016). Nav1.1 and Nav1.2 are present in GABAergic neurons of the central nervous system; missense and loss-of-function mutation of these channels create neuronal hyperexcitability and various forms of epilepsy (Kaplan et al., 2016). The most abundant Nav in the brain is Nav1.6, and loss-of-function mutation promotes gait disorders, ataxia, and dystonia. Nav1.6 is enriched in the node of Ranvier of myelinated excitatory neurons and plays an essential role in initiating and propagating the action potential by lowering the threshold voltage (Wagnon et al., 2016).
After opening, Nav channels inactivate [they have an inactivating gate (Liu et al., 2023)] and cannot be opened until the membrane potential is repolarized. While inactivated, neurons cannot fire another action potential, which is defined as the absolute refractory period. The absolute refractory period is followed by a relative refractory period in which sufficient Nav channels are open to produce an action potential. However, the threshold current to elicit the spike is increased. Thus, depending on how fast the membrane is repolarized determines the length of the refractory period and the number of action potentials a neuron can fire per unit time. It is worth mentioning that modulatory β subunits of Nav channels regulate the inactivation process. For example, β1 increases the expression, accelerates the inactivation, and leftward shifts the steady-state inactivation-voltage curve of Nav1.1 and Nav1.8 channels (Vijayaragavan et al., 2001). It has been noted that differences in expression and gating of Nav channels may have significant consequences for the generation and propagation of action potentials in nociceptive neurons (Vijayaragavan et al., 2001).
Moreover, voltage-dependent K+ channels control the threshold, the duration, and the number of action potentials a neuron can elicit per unit time. The vertebrate family of protein-forming K+ channels has twelve members (Kv1.1-Kv12; Kv5, Kv6, Kv8, and Kv9 are gating modifiers). However, given their tetrameric structure, their splice variants (e.g., K1.1-Kv1.8) can form heterotetramers, increasing the number of different Kv channels to 40 (González et al., 2012). The variety of Kv channels is further increased by their association with modulatory β subunits (Rettig et al., 1994).
A few illustrative examples of how the immense variety of Kv channels can modulate the characteristics of action potentials and, therefore, neuronal excitability are: Some Kv channels, like Nav channels, inactivate (dubbed A-type Kv channels, e.g., Kv4), but most show delayed rectifier behavior. The inactivation gate is formed by the amino terminal of A-type channels, where the first twenty amino acids bind to the internal channel vestibule once the channel opens, blocking the K+ flux [ball-and-chain model (Hoshi et al., 1990)]. Moreover, several Kv channels of the Kv1 subfamily are delayed rectifiers but show fast inactivation currents when expressed with the Kvβ1.1 subunit (Rettig et al., 1994; Heinemann et al., 1996). These inactivating Kv channels control the neuron excitability in the interspike interval. The different kinetics and voltage dependence of Kv channels determine their differential contribution to the action potential. For example, Kv1 activates at low depolarizing voltages, thus controlling the action potential threshold and the number of action potentials elicited by a depolarization induced by excitatory synaptic potentials. On the other hand, Kv channels that required larger depolarizations to open (Kv2 and Kv3) modulate the action potential duration and the firing pattern (Gola and Crest, 1993; Müller et al., 2007).
The interplay between K+ and voltage-dependent Ca2+ (Cav) channels is crucial in modulating neurosecretion. One example that illustrates the importance of the interaction of these two classes of voltage-dependent channels is the co-localization of BK and Cav channels forming nanodomains in neurons (Gola and Crest, 1993; Müller et al., 2007). The action potentials arriving at the nerve terminal open Cav channels, and both the depolarization andCa2+ entry promote the opening of BK channels, which results in a decrease in the action potential duration and a fast hyperpolarization that limit the release of neurotransmitters (Shah et al., 2022).
In conclusion, the adequate functioning of the nervous system, and most particularly of neuromodulation, is possible due to the myriad of ion channels tuned by voltage. On one direction, the rise and speed of the action potential are determined by a family of Nav channels, each with its own unique characteristics, while the duration of the action potential is determined and exquisitely modulated by the large family of Kv channels, which affects the neurotransmitter release. On the other direction, neuromodulatory inputs (detailed later on) into the distal dendrites could act to increase synaptic potential amplitudes, increase the likelihood of dendritic Na+ or Ca2+ action-potential initiation, direct action potentials to active regions of the dendrite, or simply increase action potential amplitude at the site of synaptic input, which is also crucial for neuromodulation.
3 Synapses and electrical properties of the membrane (S. C. Thébault)To understand the mechanisms of neuromodulation, it is crucial to consider the exquisite mechanisms underlying synaptic transmission. Synaptic transmission is considered the mainstay of the functioning of the nervous system. If the latter is anatomically divided into central and PNS, the basic functioning of synapses and the fact that synaptic transmission depends on the particular electrical properties of the neuronal cell membrane are valid concepts between both systems (Purves et al., 2001b; Glasgow et al., 2019).
As a brief overview of the context in which synaptic transmission is found, it has been estimated that one neuron can receive contacts from up to thousands of other cells and similarly, any one neuron can contact up to thousands of postsynaptic cells (Herculano-Houzel, 2009). A neuron can therefore be both a presynaptic and postsynaptic cell. In addition to the axon-dendrite contact, axoaxonic and axosomatic contacts happen and shape not only the input of information to the postsynaptic cell, but also the individual contribution of the cell to the local electromagnetic field. The information input determines if neurons will fire action potential or generate subthreshold membrane potential fluctuations. These two activities are important because they form membrane potential dynamics of single neurons, which, in relationship with neuronal population dynamics and computational properties emerging from the neuronal networks per se, contribute to neuronal circuit activity (Johnson et al., 2019), and subsequently neuronal functions (Glasgow et al., 2019). This multi modal and spatial–temporal communication demands the same degree of complexity of neural interfaces to be used in therapies, capable of modulating such communication mechanisms.
Chemical and electrical synapses allow neurons to communicate. Neuronal contacts can be made through a gap separating the presynaptic and postsynaptic cell and/or through gap junctions, forming a channel made of connexin proteins, which mechanically link both the presynaptic and postsynaptic cell. The former contact corresponds to the chemical synapse, whereas the latter is known electrical synapse. As the name suggests, chemical synapses use chemical messenger molecules, called neurotransmitters, which are stored in presynaptic vesicles and are released to the synaptic cleft to bind receptors in the postsynaptic neuron. This is followed by the activation of downstream signaling processes and the opening of gates, known as ion channels, to allow ions to flow into the postsynaptic cell. In contrast, electrical synapses use ions and messenger proteins that can pass through gap junctions to transmit electrical impulses and molecules.
A fundamental step in chemical neurotransmission is the release of neurotransmitters to the synaptic cleft, which is dependent of intracellular Ca2+, known from the earliest work of Dr. Ricardo Miledi and recognized as the “Ca2+ hypothesis” (Miledi and Slater, 1966; Katz and Miledi, 1968; Jeng, 2002; Eusebi, 2007). Neurotransmitters can be classified in excitatory and inhibitory, excitatory neurotransmitters allow cation influxes, and anion effluxes to increase the likelihood of an action potential firing in the postsynaptic neuron, and inhibitory neurotransmitters allow anion influxes, and cation effluxes do the opposite (IFC, n.d.). It should be noted that in addition, neuromodulators released at synapses further shape synaptic transmission (O’Brien, 2014, 2019; Alcedo and Prahlad, 2020; McCormick et al., 2020; Wu et al., 2022). The development of neuromodulation therapies is closely linked to knowledge of precise changes in synaptic transmission, such as exact thresholds of stimulation, as well as the frequency and duration of pulses needed to control desired changes in targeted neural circuits. These aspects will be discussed later.
Electrical properties of the neuronal membrane are critical to understand developing neuromodulation therapies, and in the last decades these are being incorporated in the design of in-silico models from a single axon to the whole body, which is being used to design electrical requirements for neuromodulation (Davids et al., 2019, 2020; Stefano et al., 2021). For any synapse to function, as for any cell to live, an electrochemical gradient across the cell membrane must exist (Hammond, 2015; Levitan and Kaczmarek, 2015). It is essential for the neuron to maintain a negative transmembrane electrical potential at rest, the specific value of which depends on the type of cell. For example, in the dark, photoreceptors hold a relatively depolarized membrane potential (~ −40 mV) compared to typical neurons, which have a resting membrane potential of −70 mV (Purves et al., 2001a; Henley, 2021a). Just as crucial to neuronal responsiveness to inputs are ion channels (Hille, 1987). The neuronal plasma membrane is characterized by a large density (Beaulieu-Laroche et al., 2021) and diversity of these transmembrane proteins that allow the passive movement of ions across membranes and thus, electrical signaling in the nervous system (Hammond, 2015). The properties of ion conductance’s through neuronal cell membranes are the topics of numerous book chapters and reviews (to cite a few: Levitan and Kaczmarek, 2015; Hammond, 2015; Dolphin et al., 2020). As stated in the previous section, voltage-gated Ca2+ channels are crucial for neurotransmitter release, while ligand-gated channels shape the post-synaptic response.
I would like to take the opportunity to emphasize that there is more to synaptic transmission than the classical neurophysiology of Na+, K+, and Ca2+ channels shaping action potentials (Levitan and Kaczmarek, 2015), as well as post-synaptic potentials (Burke and Bender, 2019; Henley, 2021b). As previously mentioned, synaptic transmission relies on the single neuron transmembrane potential, which is influenced by local field potentials that initiate from regulated ion channel activity (Nolan, 2016; Sinha and Narayanan, 2022), and if chemical synaptic activity is considered to be the main contributor to local field potentials (Herreras, 2016), intrinsic ionic conductances that support the existence of intrinsic oscillation generators (Whittington et al., 2018), electrical synapses, and slow fluctuations of the membrane potential of glial cells are equally important factors in the local variations of electric fields. All the above put the synaptic transmission and its modulation at the heart of neuronal communication, which demands the same level of complexity of neural interfaces to be able to decode and modulate the neuronal function. Examples of how neuromodulation underlies the flexibility of neural circuit operation and behavior by modulating synapses, synaptic strength and dynamics, and neuronal excitability have been previously detailed [for reviews, see Deer and Constant (2014), Nadim and Bucher (2014), Bazzari and Parri (2019), Pacholko et al. (2020)]. Our understanding of the modulation of the activity of neural circuits should also include the study of local variations in electric fields at the level of tissues or organisms, with a particular focus on the role of magnetic fields generated or not generated by neuronal electrical activity.
4 Neurotransmitter switching: electrical activity regulates neurotransmitter identity in health and disease (M. Pratelli, N. C. Spitzer)Before artificially modifying the activity of neuronal circuitry as occurs in Bioelectronic Medicine, we need to understand its function during natural conditions. Understanding brain development, as well as adult plasticity, may help us to decipher the highly complex self-modulation of neuronal circuitry. Beyond millisecond signaling, electrical activity acts on slower time scales to regulate the assembly and maintenance of neurons throughout the lifespan. It achieves this through coupling to cell metabolism (e.g., gene expression, protein synthesis). Spontaneous activity is essential during development to regulate neurogenesis, neuronal differentiation, programmed cell death, migration, myelination, establishment of proper connectivity, and synaptic pruning (Luhmann et al., 2016; Faust et al., 2021; Pumo et al., 2022). In the adult brain, electrical activity maintains and regulates the number, shape, and receptor composition of synapses, as well as neurotransmitter identity in behaviorally relevant circuits (Spitzer, 2017; Pan and Monje, 2020). Our lab has long been interested in activity-dependent regulation of neurotransmitter identity in different animal models. Electrical activity affects neurotransmitter identity during development and in adults.
During early development, spontaneous electrical activity generates Ca2+ transients that guide neuronal differentiation. Using the frog Xenopus laevis we showed that increasing the frequency of Ca2+ transients in neurons of the developing spinal cord increases the number of GABAergic inhibitory neurons as well as the expression of the GABA synthetic enzyme, GAD67 (Spitzer et al., 1993; Gu and Spitzer 1995). Suppressing or enhancing Ca2+ transient frequency in the neural tube stage Xenopus embryos increases the number of neurons expressing excitatory or inhibitory neurotransmitters, respectively (Borodinsky et al., 2004). Remarkably, changes in neurotransmitter identity are accompanied by matching changes in the type of receptors expressed postsynaptically, allowing the formation and maintenance of functional synapses (Borodinsky and Spitzer, 2007; Hammond-Weinberger et al., 2020). It is therefore no surprise that changes in the type of transmitter expressed influence animal behavior (Dulcis and Spitzer, 2008; Demarque and Spitzer, 2010).
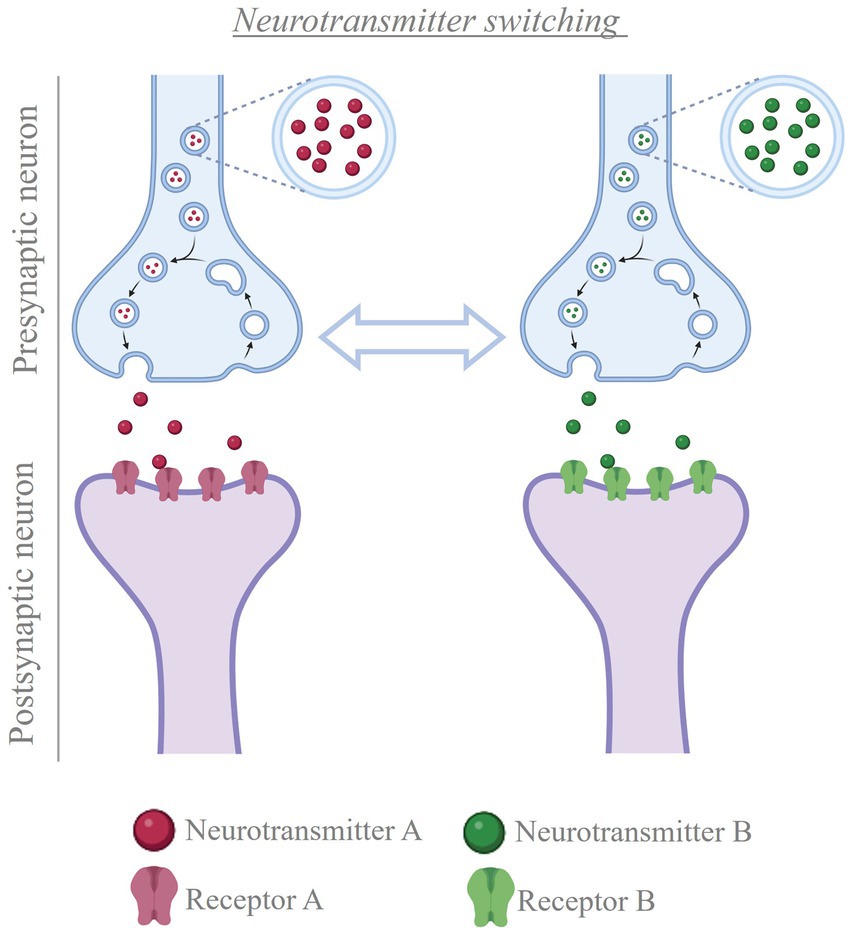
Interest in exploring the plasticity of neurotransmitter in the adult mammalian brain led to recent studies focused on showing how electrical activity regulates neurotransmitter plasticity. Multiple external stimuli that increase neuronal activity in specific brain regions cause changes in the type of transmitter expressed by local neurons in mice and rats (Gutiérrez, 2000; Gutiérrez, 2002; Dulcis et al., 2013; Spitzer, 2017; Meng et al., 2018; Romoli et al., 2019; Li and Spitzer, 2020; Porcu et al., 2022; Pratelli et al., 2022), with corresponding changes in postsynaptic receptors (Dulcis et al., 2013; Figure 5). However, when exposure to the inducing stimulus is coupled with experimentally-driven suppression of neuronal hyperactivity, the change in transmitter identity is prevented (Gutiérrez, 2000; Gutiérrez, 2002; Dulcis et al., 2013; Spitzer, 2017; Meng et al., 2018; Romoli et al., 2019; Li and Spitzer, 2020; Porcu et al., 2022; Pratelli et al., 2022). Once the change in transmitter has occurred, neuronal activity helps stabilizing the newly acquired phenotype. Experimentally-driven manipulation of electrical activity can successfully reverse the change in transmitter identity after it has occurred, rescuing the linked behavioral alterations (Pratelli et al., 2022). These findings open new opportunities when designing therapies where modulation of neurotransmission is required.
Figure 5. Neurotransmitter switching. Cartoon depicting the process of neurotransmitter switching. In the presence of a stimulus, neurons can change the transmitter expressed, from the transmitter they were expressing before to a new transmitter (e.g. from neurotransmitter A to neurotransmitter B, or vice versa). This change in neurotransmitter is accompanied by matching changes in the population of postsynaptic receptors, thus altering the signal that the presynaptic neuron conveys to the post-synaptic cells Created with BioRender.com.
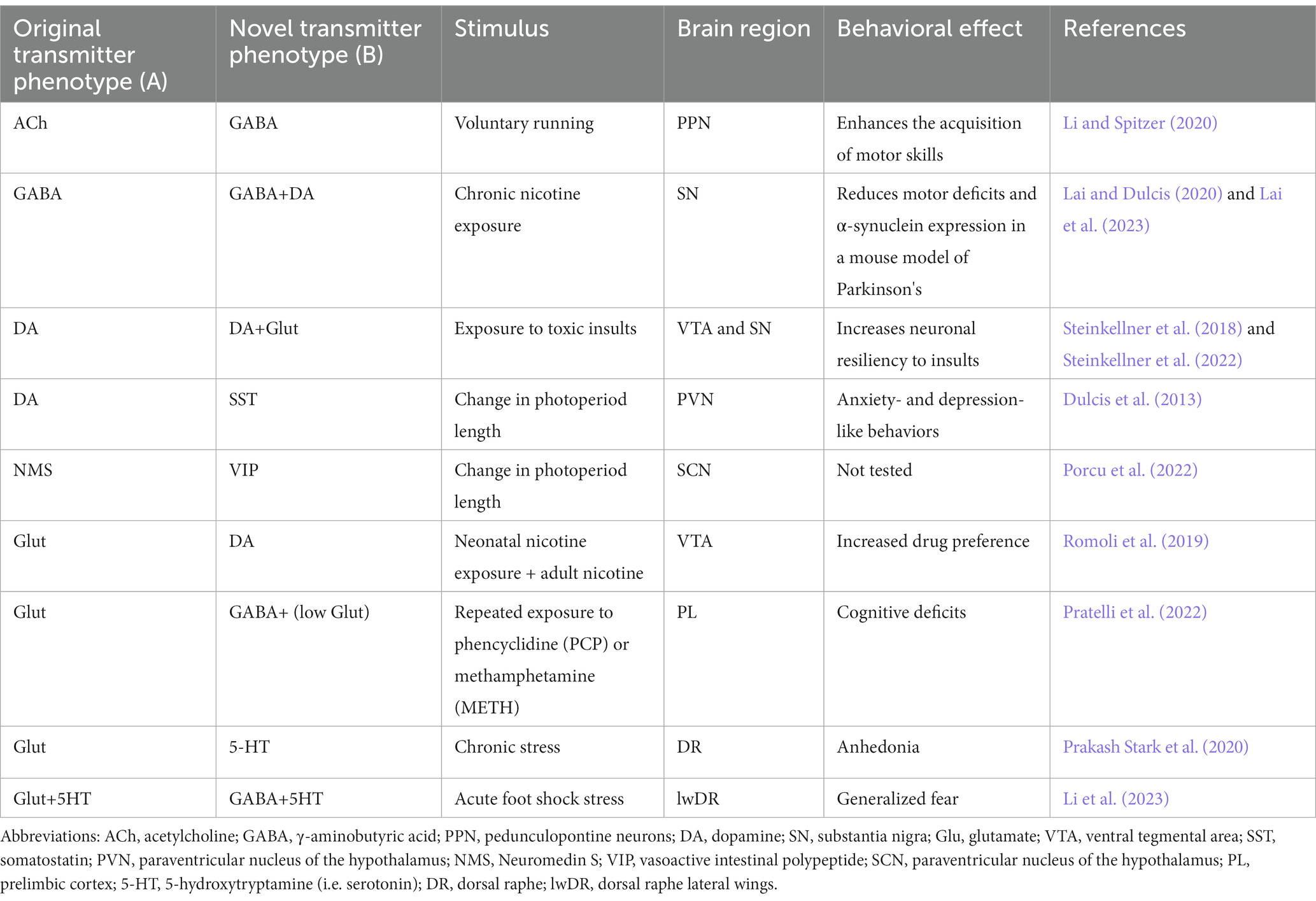
4.1 Changes in neurotransmitter identity can be beneficial or detrimentalA main question in neuromodulation is whether treatment will achieve beneficial or detrimental results. Activity induced-changes in neurotransmitter identity can substantially impact the ability of the individual to deal with the environment, by promoting either advantageous or disadvantageous changes in behavior. Beneficial effects have been linked to changes in the transmitter identity of midbrain neurons and could pave the way for novel treatment opportunities in motor and neurodegenerative disorders (Table 1; Steinkellner et al., 2018, 2022; Li and Spitzer, 2020; Lai et al., 2023). In contrast, changes in the neurotransmitter phenotype of hypothalamic neurons cause stress-induced behaviors in response to seasonal changes in photoperiod length (Dulcis et al., 2013; Porcu et al., 2022). Examples of maladaptive changes in neuronal transmitter identity are also found after prolonged exposure to drugs of abuse. In multiple brain regions, different addictive drugs cause alterations in the number of neurons expressing specific neurotransmitters (Grieder et al., 2014; Kesby et al., 2017; Ladrón de Guevara-Miranda et al., 2017; James et al., 2019; Romoli et al., 2019), some of which have been linked to drug-induced changes in behavior (James et al., 2019; Romoli et al., 2019; Pratelli et al., 2022). Activity-dependent changes in neurotransmitter phenotype linked to generalized fear and anhedonia have also been observed in the dorsal raphe in response to acute and chronic stress, respectively (Li et al., 2023).
Table 1. Neurotransmitter switching and its behavioral effects.
4.2 Opportunities for non-invasive manipulation of electrical activity to achieve health benefitsAppreciation of the extent to which activity-dependent changes in neurotransmitter phenotype influence behavior raises enthusiasm for the possibility of achieving therapeutic outcomes by leveraging neuronal activity via non-invasive stimulation methods. DBS and transcranial magnetic stimulation (TMS) allow modulation of electrical activity in the human brain and are already showing promising results in clinical settings. However, the underlying mechanisms of action remain unclear. Further spatiotemporal therapies are being developed. Changes in transmitter expression have been observed after both DBS and TMS (Trippe et al., 2009; Volz et al., 2013; Makowiecki et al., 2018), enhancing the potential use of these approaches for targeted manipulations of neurotransmitter expression. However, we are still far from being able to selectively modulate the activity of specific neuronal subtypes using these approaches. Development of non-invasive stimulation methods with higher cell-type specificity, as well as an increased understanding of the circuits and mechanisms driving activity-dependent changes in transmitter expression, are needed for optimized therapeutic intervention. The use of focused ultrasound for transcranial neuromodulation appears promising, as this approach can achieve high spatiotemporal precision and reach neurons located in deep regions of the brain (Rabut et al., 2020). This calls for a collaborative and multidisciplinary effort to promote the development of novel and optimized methods of non-invasive electrical manipulation, as well as the individuation of suitable neurobiological targets for therapeutic interventions.
In conclusion, fostering the development of effective Bioelectronic Medicine requires, among other efforts, a better understanding of how electrical activity affects behavior by causing changes in neurotransmitter identity. Ideally, experimental efforts should proceed on two levels. On one hand, it is important to understand under which circumstances experience-induced changes in neuronal activity affect neurotransmitter identity, causing behavioral alterations. This can provide valuable targets for alleviating specific symptoms via Bioelectronic Medicine. On the other hand, research should also aim to unveil the effects that different brain stimulation approaches have on neurotransmitter identity. Such knowledge can be useful not only to optimize therapeutic interventions, but also to identify potential off-target effects of brain stimulation that could lead to side effects.
5 Glial cells and neuromodulation (A. Verkhratsky, M. E. Tremblay)Neuroglia play a fundamental role in neuromodulation through comprehensive control over brain homeostasis and multiple functions, including neuronal synapses formation, activity and plasticity (synaptic pruning, stripping, neurotransmitter production and recycling), neuronal metabolism (supplying neurons with metabolic substrates), blood–brain and brain-meninges barriers (astroglial and microglial endfeet forming glia limitans perivascularis and superficialis), and immune responses (phagocytosis, release of inflammatory mediators). Neuromodulation therapies increase the capillary permeability, which likely modifies astrocytic and microglial endfeet, but could also have consequences at sites of contact with synapses and other parenchymal elements. The implant of neural interfaces triggers inflammation and instigates the formation of astrocytic barrier (erroneously referred in the literature as ‘glial scar’) and fibrotic scar around the implant, which directly interfere with the performance of both recording and stimulation neural interfaces due to the increase of resistance between electrode-neurons (see Section 7). In the neuromodulation field, the concepts of astrocytes and glia or neuroglia are often used interchangeably, which deserves clarification. It is worth mentioning that not all neuroglia are astrocytes, and astrocytes are not the only glial elements interacting with an implanted electrode as part of the foreign body response. Microglia as well as cells of the oligodendroglial lineage also need to be taken into consideration.
Neuroglia are non-neuronal and (mostly) non-excitable cells of the nervous system, which are the ultimate supportive elements responsible for maintaining homeostasis of the nervous system, for providing myelination, and for mounting defense and neuroprotection. Through multiple homeostatic pathways neuroglia play an important role in regulation of neuronal excitability, synaptic plasticity, ion homeostasis, and energy metabolism. Moreover, neuroglia are key players in the neuronal immune system having a direct impact on a pathogenesis of all neurological disorders, including neuropsychiatric and neurodegenerative diseases, such as amyotrophic lateral sclerosis (ALS), epilepsy, Parkinson’s disease (PD), Alzheimer’s disease (AD), and frontotemporal dementia (FTD) (Bazzari and Parri, 2019; Pacholko et al., 2020; Šimončičová et al., 2022; Vecchiarelli and Tremblay, 2023). Thus, in neuromodulation therapies neuroglial cells are modified and also respond to the neuronal modifications. At continuation (In this section), we present main insights of neuroglia and define concepts with relevance in the field of neuromodulation that is so crucial for Bioelectronic Medicine.
5.1 Neuroglia: definition and classificationNeuroglia are an extended class of cells populating peripheral and central nervous systems (PNS and CNS respectively) responsible for homeostatic support and defense of the nervous tissue (Verkhratsky and Butt, 2023). Neuroglia of the PNS are represented by Schwann cells (myelinating, non-myelinating, perisynaptic and subcutaneous), satellite glia residing in sensory and autonomic nervous system ganglia, and enteric glia that provide physiological support for the enteric nervous system (Verkhratsky and Butt, 2023). Neuroglia of the CNS are classified into macroglia (astroglia, wrongly referred as astrocytes in the neuromodulation field (see Section 5.3), oligodendroglia, and microglia. Regarding the embryonic/developmental origin, neuroglia of the PNS and macroglia of the CNS are of neuroectodermal origin (PNS neuroglia originate from the neural crest, whereas macroglia from neuroepithelial cells of the neural tube); microglia are of mesodermal origin, being the scions of fetal macrophages that enter and populate the neural tube in the early embryonic development (Verkhratsky and Butt, 2023). Neuroglia emerge early in the evolution (Verkhratsky and Nedergaard, 2016); the very first neuroglial cells shape and support peripheral sensory organs (known as sensilla) and provide homeostatic support to neuronal ganglia and primitive CNS in invertebrates (Verkhratsky et al., 2022). Microglia cells show remarkable evolutionary conservation of their morphology, function and genetic signatures from leech to human (Geirsdottir et al., 2020). All neuroglial cells constantly communicate with other glia, with neurons and the vasculature; such tightly coordinated communications are central for the integration of all cellular elements into the active milieu of the nervous tissue, and underlie its functional activity (Escartin et al., 2021; Lim et al., 2021).
The homeostatic function of neuroglia is executed at all levels of organization from organ to organism. Radial glia are the universal neural progenitors, while in developing brain nascent glial cells (microglia, astroglia and oligodendroglia) provide the migratory pathways for neural cells, shape the nervous system cyto-architecture, regulate synaptogenesis and synaptic remodeling through synaptic pruning, and provide for myelination of axons thus securing connectome. Astroglia and oligodendroglia regulate ionostasis and homeostasis and turnover of many essential neurotransmitters and neurohormones; some astroglial cells act as a central chemosensors thus being involved in systemic homeostatic control. Astrocytes, which are a subtype of astroglia (see Section 5.3), form the parenchymal part of blood–brain barrier an
留言 (0)