



記住我


Dexterous, effortless and reproducible hand movements represent a determinant feature of human behavior: they allow interaction with the surrounding environment, manipulate and craft objects and tools, generate complex non-verbal forms of communication and satisfy indispensable needs according to external contingencies. Since the first experiments in the early twentieth century on direct cortical stimulation, investigations on the anatomical and functional substrates of pure motor responses first and complex, meaningful hand motor actions later have shown a growing trend with an intensification in the last three decades, fostered by the expanding number of available invasive and non-invasive study modalities. The former, mostly in non-human primates, clarified that motor behavior control lies on the cortico-subcortical sensory and motor input integration throughout reciprocal modulations of primary motor, parieto-premotor and cortico-thalamic loops (Alexander and Crutcher, 1990; Rizzolatti et al., 1998).
In monkeys, previous intracortical micro-stimulation (ICMS), cytoarchitectonic and functional imaging studies clarified the involvement of a frontoparietal system connecting areas of the inferior parietal lobule (IPL) and the ventral premotor (PMv) cortex in the selection process and online control of purposeful goal-oriented hand actions (Belmalih et al., 2009; Gerbella et al., 2011; Maeda et al., 2015; Borra et al., 2017) This large-scale network, centered on vPM and extended to prefrontal and temporal areas, has been defined as lateral grasping network and is supposed to shape the motor output integrating sensorimotor information with higher order inputs as action goals and object’s features derived from the context or retrieved from memory (Borra et al., 2017; Borra and Luppino, 2019). Similarly, consistent data from healthy human subjects showed a homolog topography comprehending areas within the supramarginal gyrus (SMG), the angular gyrus (AG), intraparietal sulcus and the ventral premotor cortex (vPM) at the core of a vast network encoding objects properties, contextual information and behavioral adaptation schemes for generating complex hand-limb motor actions (Jeannerod et al., 1995; Rosenzopf et al., 2022; Sartin et al., 2022). Converging findings from different study modalities agree that non-human and human primates might share a common functional architecture subserving goal-directed actions comprehending prehension and manipulation of objects and tools (Orban and Caruana, 2014). In both species, an analog duality in dorsal frontoparietal connectivity (i.e., a dorsomedial pathway specialized in the visuo-motor integration for reaching and limb lifting and dorso-ventral pathway encoding sensorimotor integration and more direct access to motor output through parieto-premotor and premotor-motor projections) has been identified, segregating the control of specific and complementary features of hand motor schemes (Geschwind, 1975; Goldenberg, 2009; Grafton, 2010; Nelissen and Vanduffel, 2011; Glover et al., 2012; Caminiti et al., 2015). The evolutionary gain in humans led to an expansion of multimodal areas in the frontal, temporal and parietal lobes, probably due to an evolutionary-guided alteration of the macaque’s pre-existing dual-stream frontoparietal network areas. In this view, this modification allowed more complex behavioral responses—which are not affordable for non-human primates—to be encoded (i.e., praxis abilities, complex communicative limb gestures and abstract manipulative tasks; de Waal and Ferrari, 2010; Van Essen et al., 2016; Dressing et al., 2018). Accordingly, fMRI studies in humans described a wider fronto-temporo-parietal network, defined as “the praxis representation network” (PRN), consistently involved in elaborating conceptual and sensorial knowledge into goal-directed and specialized hand motor actions (Frey, 2008; Króliczak and Frey, 2009).
The extensive knowledge about neural correlates of motor functioning has been successfully translated from animal studies to intraoperative brain mapping protocols developed to guide oncological and epilepsy neurosurgery. This intimate relation has been indissoluble since Penfield and colleagues’ first experiments on direct electrical stimulation (DES) of the human cortex eliciting motor responses, influenced by previous pioneering animal experiments Penfield witnessed and collaborated on under Sherrington’s guide in the early 20th century. Since then, animal studies have inspired and provided a solid background to many brain mapping studies in intraoperative scenarios.
Awake surgery employing DES is routinely performed in specialized centers to preserve cortical and subcortical essential components of the motor network to maximally extend the resection, preserving patient’s motor abilities (so-called “onco-functional balance”; Duffau and Mandonnet, 2013). Despite several advancements, a comprehensive exploration of the mechanisms underlying object-oriented dexterous hand movements in pre- and clinical scenarios remains a relevant challenge. The old-world monkeys, like macaques, represent the closest ancestor of Homo Sapiens in which, through invasive neurophysiological and anatomical studies, detailed anatomical and physiological notions on the neural bases of sensorimotor and high-domain cognitive functioning can be carried out and generalized to human models. However, despite undoubted similarities between species, the about 30 million years of independent evolution drove significant differences in brain architecture and function, contributing to the peculiar cognitive capabilities of humans but also entangling complete transpositions of evidence from one species to the other. In addition, invasive methods with high temporal and spatial resolutions constituting the gold standard for formulating causal inference in neural mechanisms are not reproducible—for obvious reasons – in humans.
FMRI applications have been developed to fill this gap in humans; to map specific areas, activated voxels scans and analyses of time-dependent regional activities can be obtained first, then the relation of such areas to the specific function elicited can be indirectly derived with reduced spatial and temporal resolutions compared to the methods above developed in monkeys (for additional details see Hillman, 2014).
Although the translational impact of these neural substrates on the neurosurgical practice is indisputable, the anatomo-functional consistency between preclinical data gathered on primates and the results obtained in neurosurgical settings with brain mapping protocols has not yet been fully analyzed and discussed. A systematic review of this topic is a critical step in shedding light on the influence of cross-species basic science on neurosurgical practice, directly impacting patients’ quality of life and survival expectations. The current study aims to systematically collect evidence on the neural substrates of object-oriented hand manipulation movements in healthy non- and human primates and from intraoperative studies investigating responses after direct electrical stimulation of hand movement-related cortical areas and subcortical structures.
2 Methods 2.1 Information sources and search strategyA systematic literature review was conducted according to the Cochrane Handbook for Systematic Reviews and was reported based on the PRISMA statement for reporting systematic reviews and meta-analyses (Page et al., 2021). The aim of the current study was clarified through the definition of the following questions:
i. Query 1 (Non-human primate and human grasping investigations; from here referred to as Q1): What is the evidence on the anatomical-functional substrate of objects/tools grasping and hand manipulation skills in non-human primates and healthy humans? What are the homologies and differences between species?
ii. Query 2 (Intraoperative awake grasping and hand manipulation tasks investigations: from here referred to as Q2): What implication have the evidence collected from non-human primates and healthy humans in designing and implementing intraoperative advanced brain mapping paradigms to preserve grasping capacity, hand dexterity and fine motor abilities during awake surgeries? What is the state of the art on the intraoperative mapping and monitoring of grasping and/or any additional fine hand motor task during awake brain surgery?
A systematic literature search was conducted in three biomedical databases: (i) PubMed, (ii) EMBASE, (iii) Web of Science. The search was updated to 28 February 2023 and further updated on 15 September 2023). To the best of our knowledge, no additional study published after this date and available through a literature search in the databases reported above could be included in our study.
2.2 Inclusion criteria 2.2.1 Query 1For Query 1 (Q1), the following PICO terms were used: “(Grasping) AND ((fMRI) OR (MRI) OR (functional MRI) OR (PET)).” All studies had to respect the following inclusion criteria to be considered in our systematic review (Figure 1):
- To report an experimental investigation on non-human primates or healthy adult human candidates with a sample size equal to or greater than two participants (we excluded single case reports given the poor level of evidence provided);
- To test a hand grasping, reach-and-grasp or hand manipulation task reporting a contrast showing more significant activation levels for the execution task than a control condition. Control conditions include passive view, reach, simple finger movements, object detection and object discrimination);
- To declare a measurement of brain activity during the active execution of the task mentioned above as the study’s primary outcome. Comparative assessment of brain activity during planning or passive tasks was reported as secondary findings if the study’s primary goal fit all inclusion criteria. Studies focused on brain activity during resting or planning phases were excluded;
- To use fMRI or PET to measure neural activity indirectly;
- To have conducted a ROI-base or whole-brain analysis;
- To have performed a univariate, a multivoxel pattern (MVPA), or a functional connectivity analysis (aiming to qualitatively summarize the body of literature irrespectively of the nature of data provided, a consensus among authors was achieved for interpreting the impact of results expressed through different types of analyses; L.T., L.M., L.V., M.R.);
- To report activation areas in Montreal Neurologic Institute (MNI) or Talairach coordinate spaces (TAL); studies reporting findings in native space were excluded.
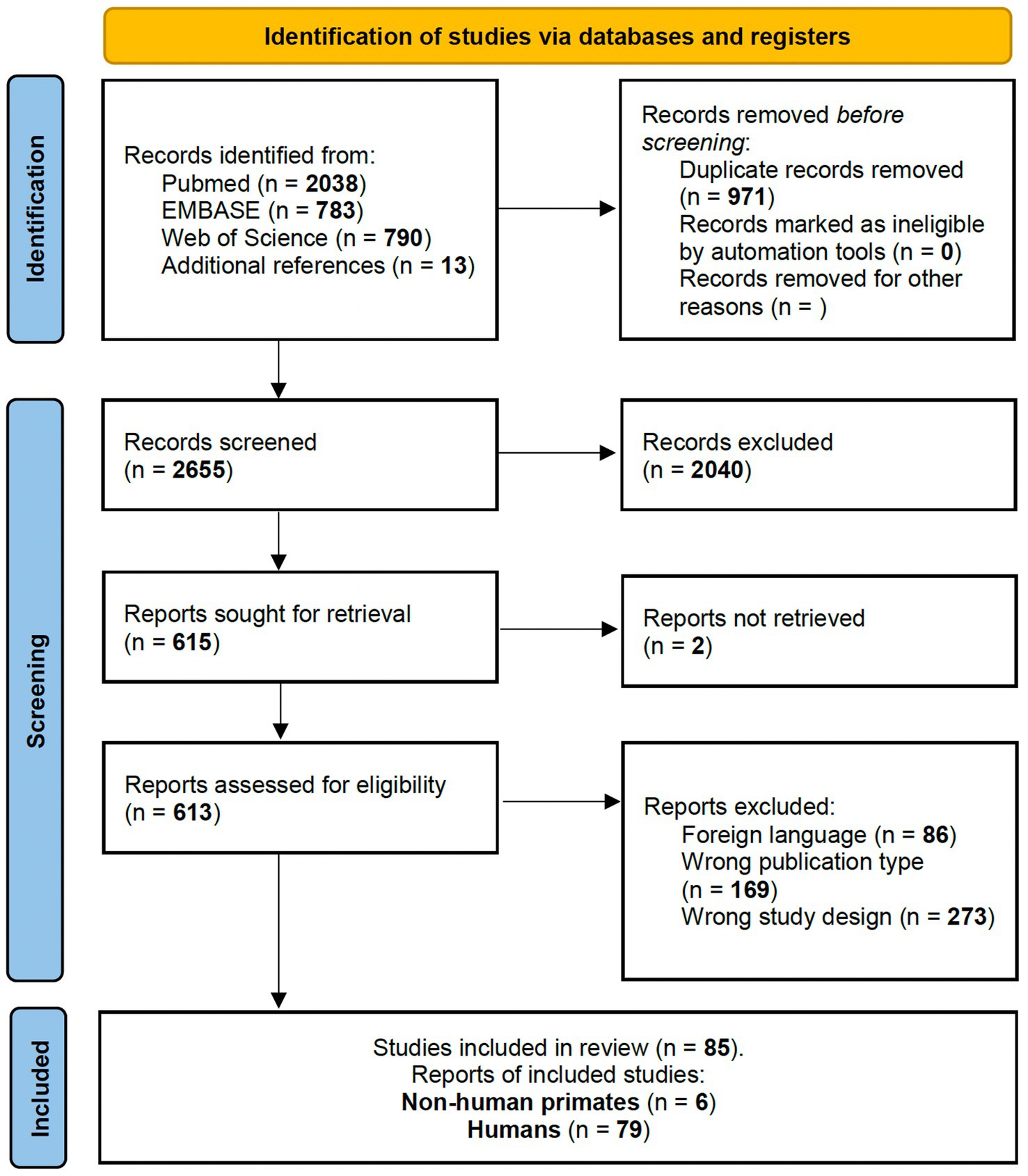
Figure 1. PRISMA 2020 flow diagram for studies involving non-human primates and healthy human participants investigating brain activated region during grasp-related fine hand gestures.
2.2.2 Query 2For Query 2 (Q2), the following PICO terms were used: “((Dexterity) OR (grasping) OR (Fine motor) OR (Grip) OR (Haptic) OR (Hand manipulation) OR (manipulation) OR (Hand movement) OR (praxis) OR (apraxia) OR (sensorimotor network) OR (motor network)) AND ((Intraoperative monitoring) OR (IOM) OR (Direct Electrical Stimulation) OR (DES) OR (awake) OR (Intraoperative Mapping)).” All studies had to respect the following inclusion criteria to be considered in our systematic review (Figure 2):
- To report an experimental study investigating any intraoperative assessment of grasping, reach-and-grasp ability or other fine hand movement tasks (i.e., dexterity, haptic-related fine finger movements, precision grip, pinching and whole-hand power grip).
- To test the functions mentioned earlier in human patients undergoing awake surgery for any condition (i.e., intra-axial tumors, vascular lesions in so-called “eloquent areas” or epilepsy surgery) employing brain mapping with or without intraoperative neurophysiological monitoring (IOM).
- To report surgical outcomes regarding function preservation, functional independence after surgery, quality of life and/or extent of resection.
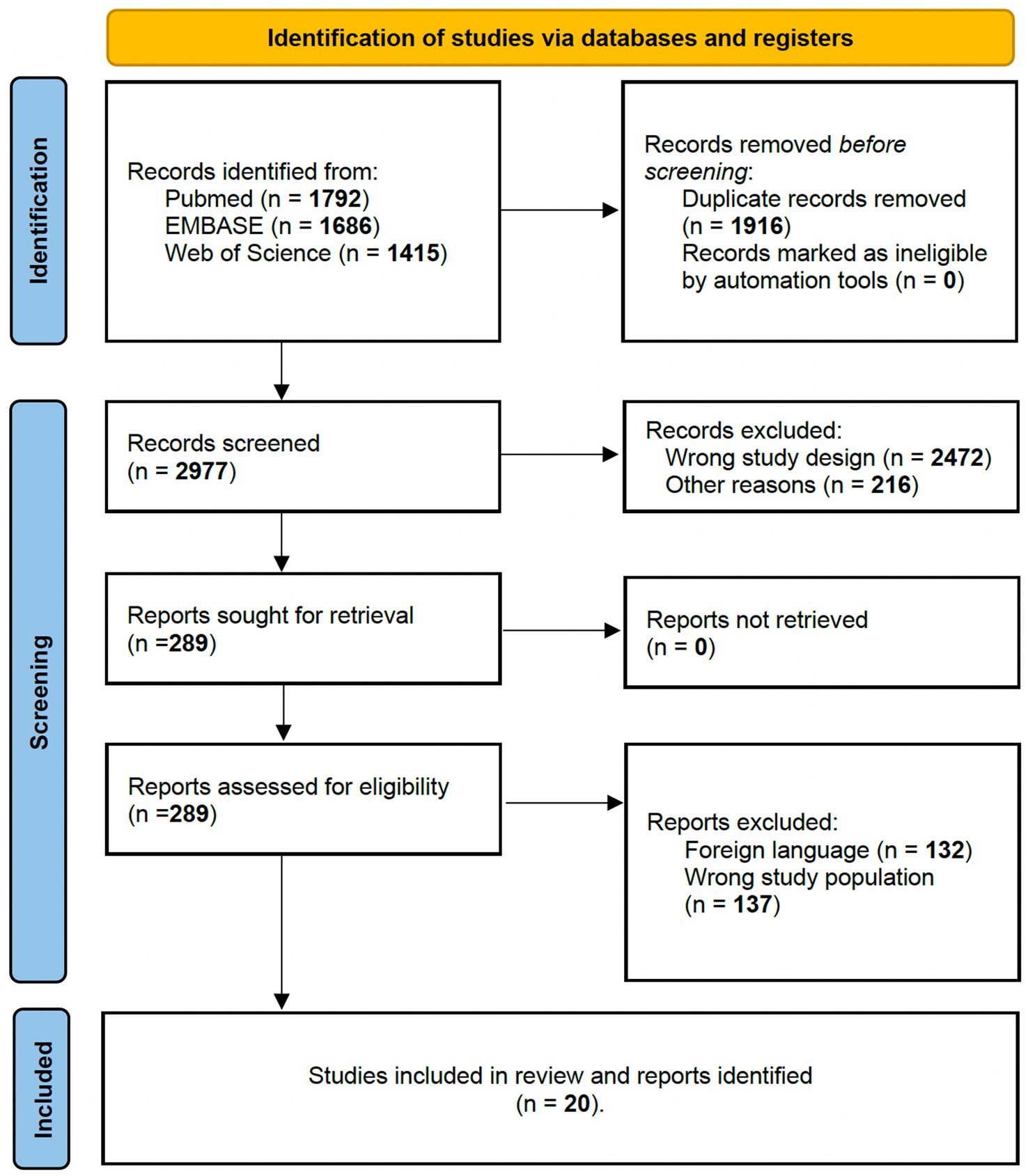
Figure 2. PRISMA 2020 flow diagram for intraoperative awake surgery brain mapping and monitoring of grasping and fine hand manipulation tasks.
2.3 Exclusion criteriaThe following exclusion criteria were applied (both Q1 and Q2):
- Publication design: Case reports, abstracts, commentaries, editorial papers, conference papers and publications written in any language but English were excluded
- Contents: all studies not fulfilling the aforementioned inclusion criteria were excluded.
2.4 Selection processTwo authors (L.T. and L.M.) independently reviewed the titles and abstracts of the retrieved articles, classifying them as included, excluded and maybe. During this stage, the articles that did not meet the inclusion criteria were excluded (such as reports written not in English, studies on non-primate animals, in vitro studies, abstracts, reviews, commentaries and case reports). In case of disagreement between the authors, the consensus was reached by full-text jointly-conducted examination. Afterwards, the full texts of the articles classified as “included” and “maybe” were independently assessed by the same authors (L.T. and L.M.). Again, in case of disagreement, the consensus was reached by broad discussion with the senior author (M.R.). No automatic tools were employed during the selection process.
2.5 Review of reportsDue to the heterogeneity in the body of evidence collected for Q1 and Q2, quantitative synthesis was deemed inappropriate. For Q1, a qualitative synthesis of results was provided: results collected from the reviewed articles were compiled through a narrative approach, and an updated description of the anatomical and functional bases of grasping and fine hand motor tasks characteristics and functional implications in non-human primates and healthy humans was provided. Similarly, for Q2, a synthesis of findings will be provided: the state-of-the-art intraoperative neurophysiological monitoring and brain mapping paradigms for hand fine movements/grasping-related tasks preservation during awake surgeries were presented in a narrative form.
Overall, to improve the readability of our qualitative review, major evidence from selected studies will be organized according to their anatomical location and connectivity, following cortical and cortico-subcortical topographical segregation. We are aware this approach will penalize the characterization and discussion of specific studies and their methodological peculiarities; however, we aimed to provide a physician-oriented comprehensive review of the most relevant cortical and subcortical nodes involved in the mechanisms under investigation with a potential impact on the neurosurgical research and clinical practice.
A detailed methodological revision of study designs and their implications was beyond the scope of the current study.
2.6 Graphical contentsThe relevant sectors of the lateral grasping network in monkeys and object-oriented hand dexterity network / praxis representative network in humans were collected and reproduced in tridimensional standardized left-hemisphere brain maps. In Monkeys, lateral grasping network sectors, comprehending the exploratory oculomotor network, were drawn on the Mount Sinai cohort data on the INIA-19 template, as previously done by Howells and colleagues (for additional methodological insights, see Rohlfing et al., 2012; Howells et al., 2020). Similarly, the relevant areas in humans were extracted by the Human Connectome Project atlas, multi-modal cortical parcellation (HCP-MMP1.0; Glasser et al., 2016). Intraoperative Data shown were extracted from Vigano et al. and Fornia et al. and rendered within the left hemisphere only (Fornia et al., 2020a, 2023). The figures in the article were designed and produced within the open-source surface render software “SurfIce” (SurfIce, 2015).
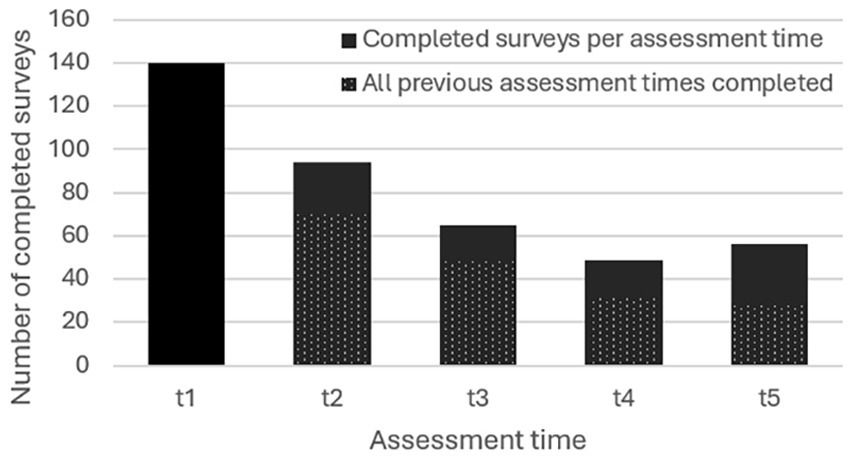
3 Results 3.1 Imaging-based experimental findingsFor Q1, bibliographic searches on literature databases yielded 3,611 records (PubMed: 2,038; Embase:783; Web of Science: 790). After removing duplicates (971 records) and unrelated manuscripts, 598 were selected for full-text evaluation. Among these, 528 were further excluded, as they did not meet the predefined inclusion criteria. Additional 13 studies were extracted from appropriate references during the screening. Overall, 85 records were included in our systematic review: six on non-human primates and 79 on healthy human candidates. Figure 1 shows the flow diagram of the literature search and study selection.
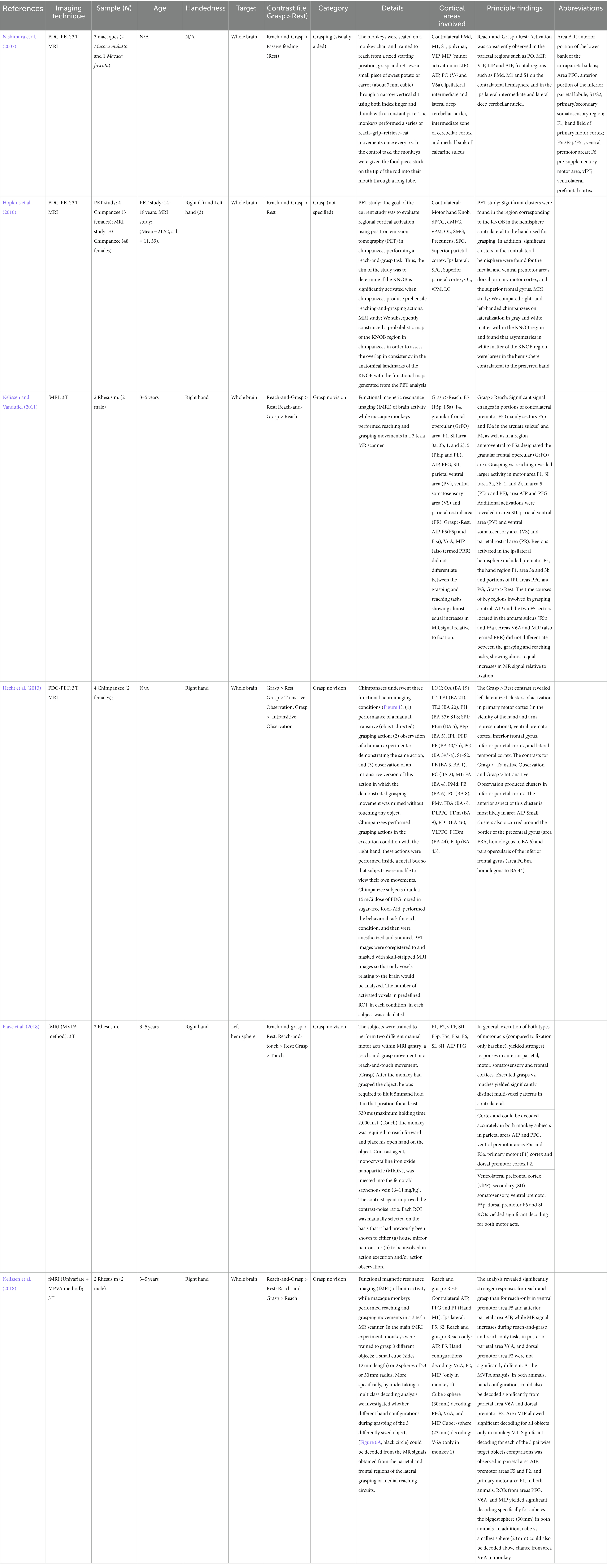
3.1.1 Non-human primatesSix studies (Nishimura et al., 2007; Hopkins et al., 2010; Nelissen and Vanduffel, 2011; Hecht et al., 2013; Fiave et al., 2018; Nelissen et al., 2018) on non-human primates were completed between 2007 and 2018 and included overall nine macaques and 78 chimpanzees [in Hopkins et al., 2010, only four chimpanzees underwent the behavioral task during PET scanning; 70 chimpanzees were recruited for baseline MRI scan only and further segmentation of hand knob region]. Five studies explored activation areas on the whole brain surface, while Fiave et al. focused only on the left hemisphere. Four studies conducted grasp and reach-and-grasp experiments with (Nishimura et al., 2007; Hopkins et al., 2010) or without visual aid (i.e., “grasping in the dark”; Nelissen and Vanduffel, 2011; Hecht et al., 2013; Fiave et al., 2018; Nelissen et al., 2018). All studies but Hopkins et al. allowed right hand movements only.
Three investigations used an FDG-PET imaging acquisition and further co-registration in MRI-normalized coordinates (Nishimura et al., 2007; Hopkins et al., 2010; Hecht et al., 2013), while the other three studies performed the behavioral task during fMRI (Nelissen and Vanduffel, 2011; Fiave et al., 2018; Nelissen et al., 2018).
The behavioral contrasts reported by the authors are the following: Grasp > Rest (Hecht et al., 2013); Reach-and-Grasp > Rest (Nishimura et al., 2007; Hopkins et al., 2010; Nelissen and Vanduffel, 2011; Fiave et al., 2018; Nelissen et al., 2018); Reach-and-Grasp > Reach (Nelissen and Vanduffel, 2011; Nelissen et al., 2018); Grasp > transitive and intransitive passive observation (Hecht et al., 2013); Reach-and-Touch > Rest (Fiave et al., 2018) and Grasp > Touch (Fiave et al., 2018). Additional information on the study design and findings are reported in Table 1. The results of the individual studies will be summarized in the narrative discussion and in Figures 3, 4.
Table 1. Main findings (non-human primates): Abbreviations are reported in the main table.
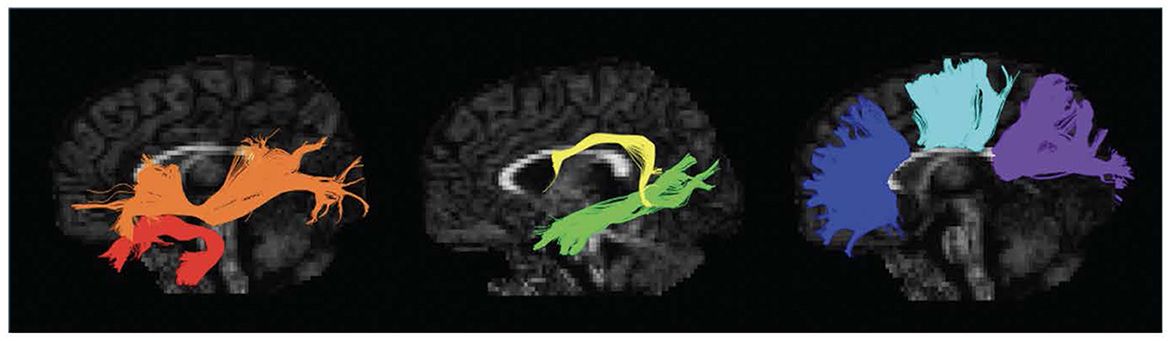
Figure 3. Grasping Network (non-human primates): Graphical representation of the lateral grasping network in non-human primates (macaques). INIA19 template. Neuromaps Atlas (Macaca mulatta). 46, lateral prefrontal cortex, area 46; AIP, Anterior IntraParietal area; F1, frontal motor area F1, macaque homolog of human M1 area; F5, frontal motor area F5, macaque homolog of human vPM area; anterior (F5a), posterior (F5p) and convexity (F5c); PFG, posterior parietal area PFG; SII, Secondary Somatosensory cortex; Insula, insular cortex (anterior); Interm12r, intermediate segment of rostral frontomesial area 12.
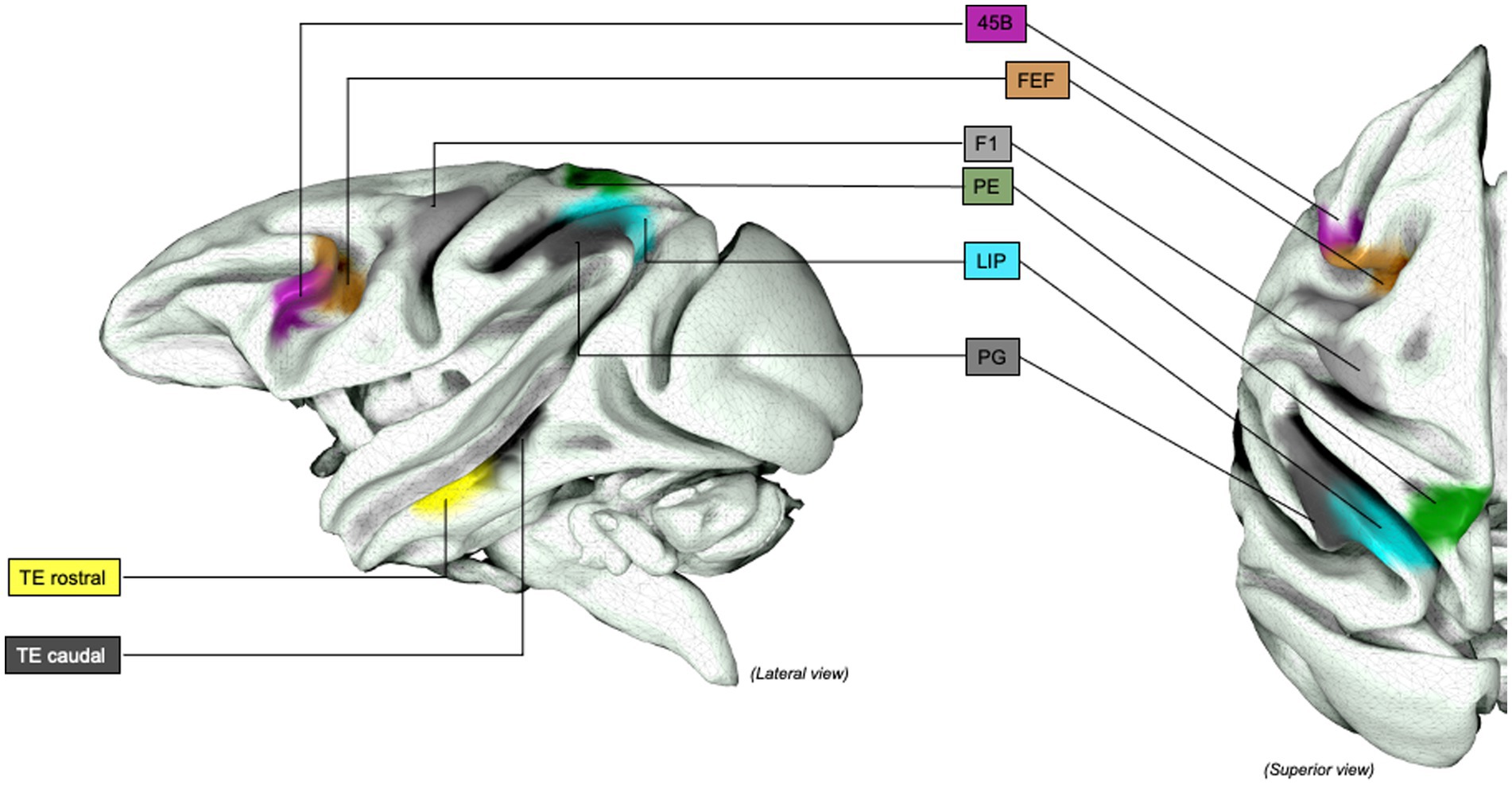
Figure 4. Grasping Network (non-human primates), extended representation: Graphical representation of the extended lateral grasping network in non-human primates (macaques). INIA19 template. Neuromaps Atlas (Macaca mulatta). 45B, part of the macaque homolog of Broca’s area; AIP, Anterior INtraParietal area; F1, frontal motor area F1, macaque homolog of human M1 area; FEF, Frontal eye Field; LIP, lateral IntraParietal Sulcus; PE, posterior parietal are PE; PG, posterior parietal area PG; TE, inferior temporal area TE, rostral and caudal.
3.1.2 Healthy human candidatesSeventy-nine studies were completed between 1996 and 2022, including 1,412 healthy patients (560 females, 9 studies did not specify gender heterogeneity; Grafton et al., 1996; Matsumura et al., 1996; Binkofski et al., 1999a,b; Ehrsson et al., 2000, 2001, 2007; Kuhtz-Buschbeck et al., 2001; Chapman et al., 2002, 2007, 2011; Culham et al., 2003; Grèzes et al., 2003; Ward and Frackowiak, 2003; Frey et al., 2005, 2015; Talati et al., 2005; Shmuelof and Zohary, 2006; Cavina-Pratesi et al., 2007; Grol et al., 2007; Króliczak et al., 2007; Milner et al., 2007; Vaillancourt et al., 2007; Begliomini et al., 2007a,b, 2008, 2014, 2015; Park et al., 2008; Stark and Zohary, 2008; Verhagen et al., 2008; Gallivan et al., 2009, 2011a; Hinkley et al., 2009; Matsuda et al., 2009; Spraker et al., 2009; Cabinio et al., 2010; Kurniawan et al., 2010; Fiehler et al., 2011; Holmström et al., 2011; Hong and Jang, 2011; Kim et al., 2011, 2021; Martin et al., 2011; Monaco et al., 2011, 2014, 2015, 2017; Neely et al., 2011; Glover et al., 2012; Makuuchi et al., 2012; Nathan et al., 2012a; Vingerhoets et al., 2012a; Renzi et al., 2013; Rossit et al., 2013; Fabbri et al., 2014, 2016; Plata et al., 2014; Gutteling et al., 2015; Pavlova et al., 2015; Hamzei et al., 2016; Leo et al., 2016; Marangon et al., 2016; di Bono et al., 2017; Gatti et al., 2017; Przybylski and Króliczak, 2017; Ariani et al., 2018; Cavina-Pratesi et al., 2018; Styrkowiec et al., 2019; Marneweck and Grafton, 2020; Sulpizio et al., 2020; Turella et al., 2020; Bencivenga et al., 2021; Knights et al., 2021, 2022; Errante et al., 2021a; Livne et al., 2022; Michalowski et al., 2022; Ras et al., 2022).
All 79 studies investigated cortical sites of activation in the left and right hemispheres. Seventy-two studies included right-handed patients, and seven included left-handed participants (Begliomini et al., 2008; Cabinio et al., 2010; Martin et al., 2011; Gallivan et al., 2011a; Vingerhoets et al., 2012b; Fabbri et al., 2014; Gutteling et al., 2015); two studies did not declare the participants’ handedness (Knights et al., 2021, 2022). Seven studies on right-handed patients investigated grasping task of the non-dominant hand (Binkofski et al., 1999b; Ward and Frackowiak, 2003; Shmuelof and Zohary, 2006; Hong and Jang, 2011; Kim et al., 2011; Vingerhoets et al., 2012a; Begliomini et al., 2015).
Twenty-two studies performed experiments with no direct view of the target of the grasping task (“i.e. grasping in the dark; Ehrsson et al., 2000, 2001, 2007; Kuhtz-Buschbeck et al., 2001; Shmuelof and Zohary, 2006; Milner et al., 2007; Spraker et al., 2009; Kurniawan et al., 2010; Fiehler et al., 2011; Holmström et al., 2011; Hong and Jang, 2011; Kim et al., 2011, 2021; Neely et al., 2011; Renzi et al., 2013; Fabbri et al., 2014; Marangon et al., 2016; Gatti et al., 2017; Ariani et al., 2018; Cavina-Pratesi et al., 2018; Styrkowiec et al., 2019; Turella et al., 2020). Moreover, eight studies reported specific hand manipulation tasks with (Binkofski et al., 1999a,b; Pavlova et al., 2015; Michalowski et al., 2022; Ras et al., 2022) and without visual aid (Talati et al., 2005; Marangon et al., 2016; Styrkowiec et al., 2019), and two studies acquired functional imaging during a pointing behavioral task (Frey et al., 2005; Cavina-Pratesi et al., 2018).
Additional information, including behavioral contrasts implemented, is available in Supplementry Tables 1–3. The results of individual studies will be summarized in the narrative discussion and reported in Figures 5, 6.
Figure 5. Object-oriented hand manipulation network in humans: Graphical representation of the object-oriented hand manipulation network (within the praxis-representative network) in humans. Atlas, Human Connectome Project-MMP1.0; 3a-4, Brodmann areas 3a-4; 5 m, Brodmann area 5 medial; 6ma, Brodmann area 6ma (preSMA, mesial SFG); 6mp, Brodmann area 6mp (SMA, mesial SFG); 6d, Brodmann area 6d, premotor area dorsal; 6v, Broadmann area 6v, premotor area ventral; AIP, Anterior IntraParietal area; PF, Inferior Parietal Lobule area PF; PGp, posterior portion of human Angular Gyrus.
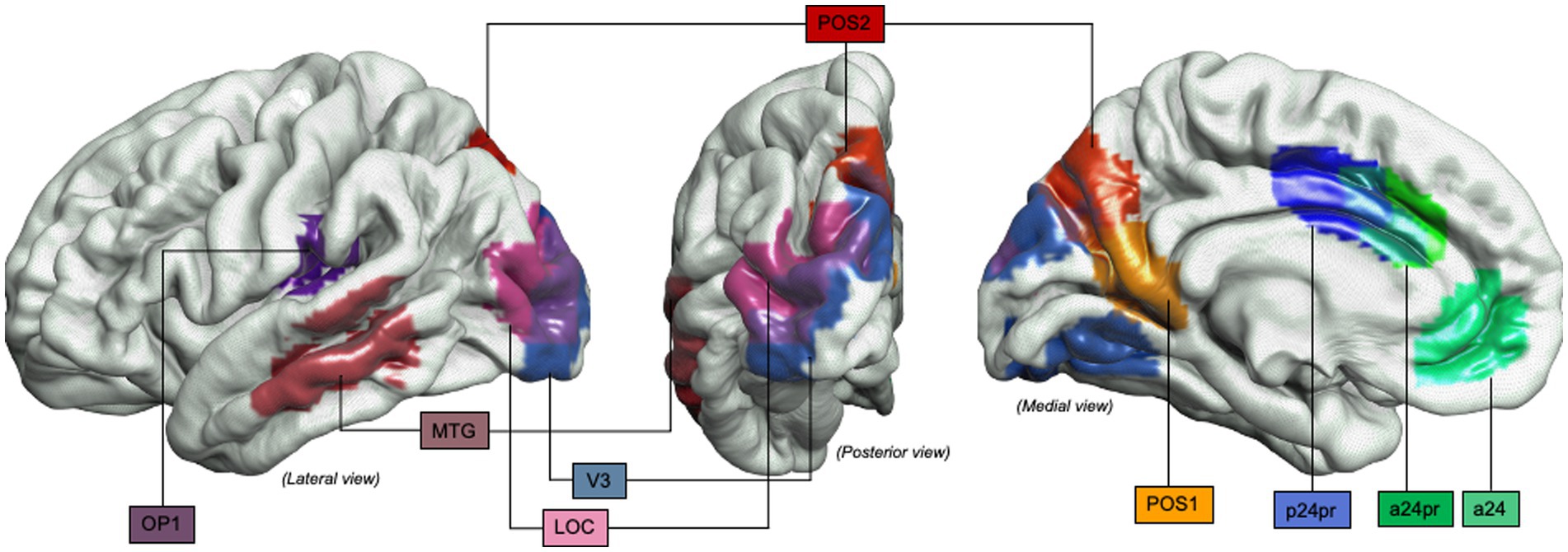
Figure 6. Object-oriented hand manipulation network in humans (extended view): Graphical representation of the extended object-oriented hand manipulation network (within the praxis-representative network) in humans. Atlas, Human Connectome Project-MMP1.0; a24, subdivision of Anterior Cingulate Cortex; a24pr, subdivision of the anterior part of Middle Cingulate Cortex; LOC, Lateral Occipital Cortex; MTG, Middle Temporal Gyrus; OP1, Operculum Parietal 1 area; p24pr, subdivision of the posterior part of Middle Cingulate Cortex; POS, Parieto-Occipital Sulcus; V3, Visual Area 3.
3.2 Intraoperative awake surgery findingsFor Q2, bibliographic searches on literature databases yielded 4,892 records (PubMed: 1,790; Embase:1,686; Web of Science: 1,415). After removing duplicates (1,916 records) and unrelated manuscripts, 287 records were selected for full-text evaluation. Among these, 269 were further excluded, as they did not meet the predefined inclusion criteria. Overall, 20 records were included in our systematic review. Figure 2 shows the flow diagram of the literature search and study selection. The results of individual studies will be summarized in the narrative discussion (see also Table 2 and Figure 7).
Table 2. Main intraoperative findings during awake brain mapping procedures (human primates).
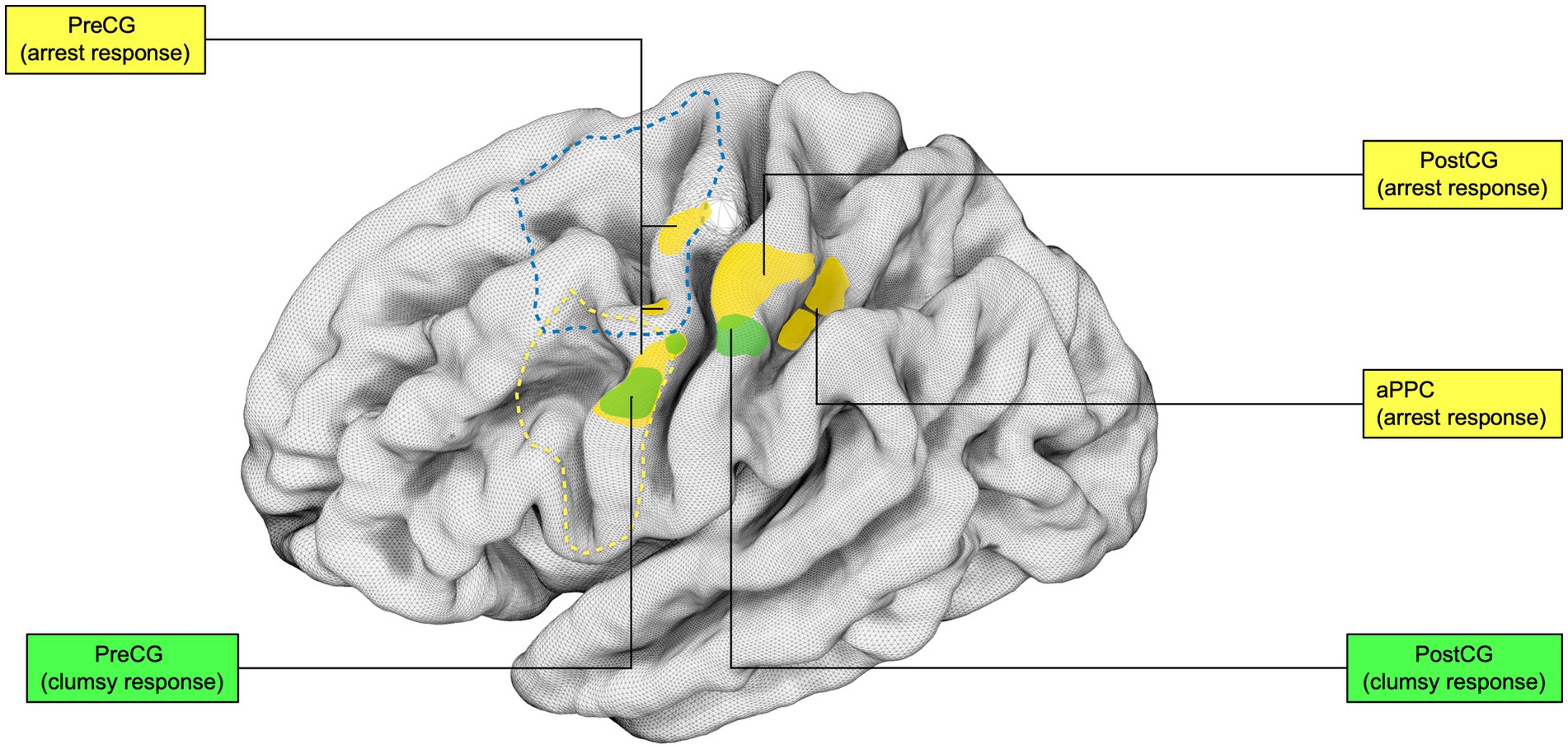
Figure 7. Intraoperative brain mapping stimulation sites distribution during DES-induced hand manipulation task (hMT) disruption: Distribution of stimulation sites in the standardized left hemisphere. The sectors are segregated according to the EMG and behavioral response during DES administration and online execution of the hand-manipulation task (hMT). The “arrest response sites” were defined as those anatomical sectors containing effective sites associated with deficient muscle performance, characterized by complete task arrest during DES. The “clumsy response sites” were defined as those anatomical sectors for which effective sites were associated with high variability among muscles, possibly reflecting poorer muscle coordination. The arrest response pattern is clustered in the ventral and dorsal sectors of the precentral (PreCG, PMd), the postcentral gyrus (postCG) and the anterior sector of the posterior parietal cortex (aPPC). In contrast, the clumsy pattern is identified in the dorsal vPM and the PostCG’s mid-portion. Not represented in the figure: Within the arrest behavioral responses, a “suppression response,” determined as a hand movement arrest and a general decrease in muscle recruitment, was identified in the precentral gyrus in correspondence with a sector within the dorsal vPM and in within arrest sites within aPPC, PostCG and anterior supramarginal gyrus. Similarly, a “recruitment response,” characterized by an involuntary movement and a general unspecific increase of muscles activity within an arrest behavioral phenomenon, was localized in a dorsal sector of PMd and a caudal PostCG sector. Mixed muscle effects were also identified within the caudal PostCG sector. Yellow, arrest response; Green, clumsy response.
4 Discussion 4.1 Query 1: Non-human primates 4.1.1 Anatomical substrates of lateral grasping network in monkeysThe classical paradigm for monkey prehension network (namely the “lateral grasping network”) was initially framed on parallel cortico-cortical pathways connecting neurons of the posterior parietal (PPC) and frontal cortex exhibiting similar architecture and functional responses. The ventral premotor area F5 is one of the most relevant areas in object motor grasping and receives inputs from the inferior parietal lobule (IPL) areas, namely AIP, PF, PFG and SII, projecting to the primary motor cortex (F1) and spinal cord (especially F5p; di Bono et al., 2017). On the medial frontolateral convexity, the rostral part of area F2 (F2vr) directly integrates grasping control, specifically during wrist rotation and orientation toward a target. However, it is also involved in reaching, although weaker evidence is available (Dum and Strick, 2002; Gamberini et al., 2009; Hecht et al., 2013). Imaging-based studies investigating the role of F3 (proper-SMA in monkeys) and F6 (pre-SMA area) in primates are less reported: primary evidence from single-neuron studies advoked the involvement of F6 in learning, execution of sequential motor behaviors and the initiation of conscious action. Accordingly, F6 neurons were reported preferentially activated while learning a sequence of actions compared to subsequent execution of the learned scheme in a behavioral study on monkeys (Nakamura et al., 1998). Based on these findings, it is widely accepted that area F6 controls action time and appropriateness.
Several prefrontal cortex areas are connected to the premotor and parietal cortex and areas 46 (46dr and 46d, 46vr and 46v) and area 12r are the most represented on dorsal and ventral banks of the principal sulcus (Gerbella et al., 2013; Saleem et al., 2014; Bufacchi et al., 2023). In a recent study, Luppino et al. reported that prefrontal areas might be subdivided into three caudo-rostral strips according to their primary output connectivity. From rostral to caudal, the first strip carries intrinsic lobar connections within the prefrontal cortex, connecting the frontal pole and orbital prefrontal areas (Borra et al., 2011; Saleem et al., 2014). The caudal part shows significant connections with the lateral intraparietal cortex (LIP), frontal eye field (FEF), supplementary eye field (SEF) and other subcortical nodes involved in gaze control, while the intermediate cluster is mostly interconnected with premotor, parietal areas and subcortical structures involved in grasping and reaching movements (Gerbella et al., 2013; Saleem et al., 2014; Borra et al., 2017). The ventral prefrontal areas (VLPF) share a common connectivity with the temporal lobe, suggesting a major role in processing object semantic features, while the dorsal prefrontal cortex (DLPF) is strongly connected to the parietal and premotor areas, as area 46 is involved in motor and behavioral control of grasping movements (Goldman-Rakic, 1987; Hecht et al., 2013).
The most reported parietal nodes of the grasping network are the anterior infraparietal cortex (AIP) and the adjacent area, PFG, both dense in neurons encoding grasping movements execution (Hyva¨rinen, 1981; Taira et al., 1990). Dorsally, MIP and the dorsal part of area V6A (V6Ad) were later attributed to reaching control (Andersen et al., 1997). The former have dense interconnections, except PFG, which only shows selective connectivity with AIP. The latter also receive input from higher-order associative visual areas in the temporo-occipital cortex (Rozzi et al., 2006). Overall, this sector mediates visuomotor transformation for reaching and grasping, processing target features (size, orientation, position and shape) and activating appropriate potential motor actions. This process, called “affordance extraction, “elicits objects’ visual properties transformation and projection through the dorsal stream (Sakata et al., 1997).
Finally, the inferior temporooccipital and the posterior occipital regions define the “ventral visual stream” involved in visual information processing, object identification and semantic recognition (Tanaka et al., 1996). Nelissen et al. identified cortical activations during action observation at the level of the upper (superior temporal polysensory area or STP) and lower banks (Tea/m sector) of the superior temporal sulcus (STS). STP is a high-order multisensory sector integrating information encoded in multiple sensory modalities, also populated by visual neurons encoding self-produced and external motion features connected to the PFG area in parietal lobe (Baylis et al., 1987). Tea/m sector was identified as a ventral visual node specialized in three-dimensional object and action processing (Orban and Caruana, 2014). Information encoded in STS is projected through Tea/m to AIP and the mirror system in the parietal lobe, providing input of action goal performed by others, intrinsic information to identity the target object as a substrate for affordance extraction, but also along STP-PFG pathway to elaborate the intention and the goal of the observed action (Jellema et al., 2000). An additional area in the parietal operculum, SII area, hosts visual-responsive neurons firing during external action observation, suggesting that it might interact with temporoparietal projections as a multisensory integrating node for motor control and action recognition (Hihara et al., 2015).
4.1.2 Functional modeling of the grasping network in monkeysThe dorsolateral and dorsomedial pathways of the lateral grasping network are essential for sensorimotor integration (i.e., planning and online control) during reaching, grasping, and gaze control (Rizzolatti et al., 1998; Andersen and Buneo, 2002). According to this classical model, a dorsolateral pathway encodes grasping and different grip features, while a dorsomedial pathway encodes reaching and control of the transport/lifting phase (Jeannerod et al., 1995; Caminiti et al., 1998; Culham et al., 2003).
The dorsolateral pathway comprehends AIP (Murata et al., 2000) and subareas F5p/F5c of the ventral premotor cortex (PMv; Murata et al., 1997; Rizzolatti et al., 1998; Nishimura et al., 2007; Nelissen and Vanduffel, 2011; Nelissen et al., 2018). Similarly to AIP, PFG area responded to hand movements observed but even without a target or the presentation of 3D objects, implying a pivotal role in grasp motor scheme planning. Of note, an additional neural category responding to passive viewing of actions performed by others and peri-personal space awareness in PFG (I.e. “mirror neurons”) was characterized (Rozzi et al., 2008; Hopkins et al., 2010; Hecht et al., 2013; Fiave et al., 2018; Nelissen et al., 2018). A core description of the extended mirror network is beyond the aim of the current review (for a review, see Rizzolatti and Craighero, 2004).
AIP was proposed as primer of grasping response by activating visual-dominant neurons, which extract 3D object characteristics and propagate them to F5 visuomotor neurons encoding congruent motor representations for the affordable object. To integrate the modulation of the prefrontal cortex to the intended behavior, Arbib and co-workers proposed the “Fagg–Arbib–Rizzolatti–Sakata (FARS) model” (Fagg and Arbib, 1998; Arbib and Mundhenk, 2005). AIP produces multiple motor representations of object affordances to F5, while modulated by prefrontal inputs, encoding the goal of the individual in affording the target object; this whole process selects the most appropriate motor execution program. The robustness of this model relies on the fact that F5 is not directly interconnected with the inferotemporal cortex but receives dense connections from the ventral prefrontal cortex, which in turn receives input from inferior and posterior temporal areas, as previously described.
The dorsolateral pathway encoded the transformation of intrinsic target properties into appropriate behavioral and motor commands comprehending hand pre-shaping, force adjustment and type of grip during visually-aided grasping (Jeannerod et al., 1995) through visuomotor neurons in AIP, PFG and F5, with significant activation while observing a graspable object or performing a grasping task (Bonini et al., 2014). Lesion studies within this pathway clarified how lesions within AIP and F5p affected hand pre-shaping and wrist orientation, leaving object-reaching ability mostly unaffected. These findings were consistent only during precision grip of small objects, while whole hand prehension showed no deficit, confirming the crucial role of AIP and F5p during complex sensorimotor control or pinching of small objects (Ehrsson et al., 2001; Cavina-Pratesi et al., 2007; Grol et al., 2007; Begliomini et al., 2007a, 2014; Renzi et al., 2013; Monaco et al., 2015). Neighbor area F5c (F5 subarea) lesion, despite having the same visuomotor properties as other F5 subareas, was not responsible for any grasping impairment (Fogassi, 2001; Bonini et al., 2014).
The selection of the object’s meaning also relies on connections from prefrontal area 12r, while the behavioral response based on the overarching goal could be appointed to prefrontal area 46v, which is densely connected with posterior parietal and premotor cortex. Eventually, the affordance selection elicits F5 motor representation, activating F1(the primary motor area). Once affordance is selected and hand shaping programmed, additional features modulate arm-hand movement and grip characteristics. SII region, for example, was activated during movement, especially object grasping and different hand configurations, object orientation and passive view: it is plausible to consider this area as a fundamental sensorimotor integration node for object’s physical and visual properties during reaching and grasping, receiving feedback information used in F5, AIP and PFG for online monitoring and update grasping motor scheme (Hihara et al., 2015).
The dorsomedial pathway connects the PPC, among all V6a (Bosco et al., 2010), VIP and MIP (Johnson et al., 1996) with F2 anteriorly, within the primate dorsal premotor cortex (PMd), and associative visual areas posteriorly (Nishimura et al., 2007; Nelissen and Vanduffel, 2011; Fiave et al., 2018; Nelissen et al., 2018). MIP and AIP are part of both ventrolateral and dorsomedial pathways, confirming the integration and overlap of the two pathways. Visual areas V1–V4 are strongly connected to V6A and, in particular V1 and V2 subareas, are involved in the representation of peripheral visual field showed the most substantial convergence onto V6A, with similar attributes toward peripheral vision and stimulus position in craniotopic coordinates encoding (Daniel and Whitteridge, 1961; Galletti et al., 1999). Despite previous modeling including the precuneus in the dorsomedial stream only, a substantial dorsomedial and dorsolateral pathways overlap was later identified, whose direct function in reaching and grasping remains only partially understood. Tracer studies in monkeys revealed projections to both area F5 and V6A, suggesting a role in coordinating reaching and grasping, in addition to the well documented activation during bimanual performance, complex tasks and (Johnson et al., 1996) as a functional hub in the default mode network (DMN) in monkeys (and humans); however, this area of grasping-reaching overlapping does not seem to correspond with the DMN activation sites in fMRI (Johnson et al., 1996; Fransson and Marrelec, 2008; Mantini et al., 2011). Of note, the frontal eye field (FEF, area 8 m) area is also strongly connected to the precuneus, suggesting a role during sensory guidance of limb and hand movements as much as a hand-eye coordination node (Thompson et al., 1996).
Overall, an integration gradient on the functional level between the dorsomedial and ventrolateral streams seems consistent. Although the ventrolateral stream is primarily integrated with somatosensory areas and the dorsomedial stream the has more robust interconnections to the visual cortex, several areas within each pathway are strongly interconnected. Grasp planning and execution revealed significant activation in the IPL convexity areas posterior to AIP, comprehending area PF, PG, and particularly area PFG, having direct connections to F5 (Bonini et al., 2011, 2014). Similarly, neural activity during grasping was recorded in V6A and PMd subareas (i.e., area F2vr; Fattori et al., 2010, 2012). Conversely, subpopulations of F5 and AIP showed reaching-related coding in additional studies (Lehmann and Scherberger, 2013), supporting an overlapping architecture of the lateral grasping network (Battaglini et al., 2002).
4.1.3 Cortico-subcortical loops integrated with the lateral grasping network in monkeysThe cerebellum and the basal ganglia play a role in motor cognitive tasks execution and control ranging from sensorimotor to complex behavioral integration (Strick et al., 2009; Bostan et al., 2013; Caligiore et al., 2013). They fulfill a functional architecture of input projections from a wide range of cortical areas, despite earlier findings supported the idea that these connections were anatomically and functionally segregated according to the function being modulated (Middleton, 2000; Baldassarre et al., 2013). Hoshi et al. reported a strong bisynaptic connection between the striatum and the thalamus, while Bostan, Dum and colleagues showed that the subthalamic nucleus has similar connections to the cerebellar cortex through the pontine nuclei (Hoshi et al., 2005; Bostan et al., 2010). Tracer studies in monkeys clarified that efferent connections from the cerebellar nuclei are projected to M1 but also on premotor, prefrontal and parietal areas (Percheron et al., 1996). The output streams to M1 and premotor areas within the dorsal dentate nucleus are clustered in a circumscribed “motor” domain while a ventral region of the nucleus showed connections to the prefrontal (non-motor) areas (Shinoda et al., 1992; Dum and Strick, 2003). This functional segregation in the dentate nucleus has a counterpart in the cerebellar cortex. The dorsal “motor” dentate is majorly connected to the anterior cerebellar lobe (lobules III–VI) and paramedian lobule (HVIIB and HVIII), while the ventral dentate is connected with the posterior vermis and the cortex are not involved in motor control, as consistently documented also in imaging studies (Kelly and Strick, 2003). The cerebellar cortex elicits activation of the dentate nucleus, and this functional loop allows cortical areas (i.e., prefrontal, premotor and F1) to interconnect with the dentate nucleus via the pontine nuclei while areas not projecting to the cerebellum showed no major connections from the cerebellum (i.e., area 46v, 12 and TE, for example). This series of closed-loop circuits might represent the anatomical substrate of a multi-level modulation system involving the cerebellum as a significant player. If so, cerebellar output directly influences multiple non-motor cortical areas (i.e., premotor and parietal cortex), suggesting an essential role in various cognitive tasks (Koziol et al., 2014). Similarly, motor and non-motor subregions of STN showed a similar projection onto the motor and non-motor cerebellar cortex (Bostan et al., 2010), while Chen et al. demonstrated that the cerebellum is responsible for a short-latency direct modulation of the striatum (Chen et al., 2014). Although the growing evidence hereby reported, the physiology of cerebellar-striatal interconnections is still poorly understood in contexts other than motor functioning, and further research is prompted.
4.2 Query 1: Human primates 4.2.1 Anatomical and functional substrates of the object-oriented hand manipulation networkThe anatomical and functional substrates of dexterous motor behavioral responses, complex non-verbal communicative gestures and abstract manipulative tasks in humans—comprehending those abilities defined as “praxis”—represents the translational evolution of the pre-existing dual streams frontoparietal network (i.e., lateral grasping network) defined in the macaques. With a certain amount of approximation, we can refer to it as “praxis representation network” (PRN) in the human model.
The premotor and prefrontal areas play a significant role in the dorsoventral and dorsomedial pathways for reaching and grasping performance within the network. The human ventral premotor area (PMv or hPMv) has been proposed as the homolog of the rostral part of the nonhuman primate F5 area, with which it shares direct or indirect control of hand movements and mirror-like properties (Cerri et al., 2003, 2015; Ehrsson et al., 2007; Maranesi et al., 2012; Fornia et al., 2018). A recent rTMS study highlighted PMv features by reproducing an impairment of current finger position during precision grip, confirming its involvement in such a task (Davare et al., 2009). As mentioned above about area F2 in monkeys, the dorsal premotor area (PMd) is involved in grasping and reaching tasks, confirming that the dorsomedial frontoparietal circuit serves both grasping and reaching encoding (Raos et al., 2006; Errante and Fogassi, 2019). Indeed, the inactivation of PMv impaired hand pre-shaping, while that of PMd corresponded to interference in object lifting with disruption of grasping-lifting coupling, conforming a different but convergent contribution of these areas in controlling hand/arm performance during grasping, as previously demonstrated in monkeys (Davare et al., 2006; Fornia et al., 2020b). Comparable with animal findings, additional areas involved in hand motor control are the supplementary motor areas (proper- and pre-SMA) and ventrolateral and dorsal prefrontal cortex (VLPFC and DLPFC, respectively). Several imaging studies investigating planning vs. online control of hand motion toward a target attributed a relevant inhibition and behavioral switching control to pre-SMA, DLPFC (Spraker et al., 2009) and VLPFC (Mostofsky et al., 2003; Aron, 2009; Glover et al., 2012; Tabu et al., 2012), comprehending force strength adaptation and dynamic grip modulation (Neely et al., 2011) to grasped objects (Kuhtz-Buschbeck et al., 2001; Holmström et al., 2011). Most of the studies we reviewed on healthy humans collected evidence on dominant hand grasping execution in right-handed subjects with a skew lateralization of cortical activity measured during handgrip only for the involvement of contralateral sensorimotor cortex (and ipsilateral superior cerebellum). Nevertheless, most studies consistently reported bilateral activation of PMv, PMd, SMA, cingulate motor cortex, IPL, insular, vermis and both superior/inferior cerebellar hemispheres, despite previous reports left-lateralized cortical activations in right-handers humans but a widespread bi-hemispheric sensorimotor, premotor and SMA activations in left-handed candidates performing grasping tasks with the dominant and—even more evident—non-dominant hand (Grafton et al., 1996; Ehrsson et al., 2000, 2001; Kuhtz-Buschbeck et al., 2001; Ward and Frackowiak, 2003; Cavina-Pratesi et al., 2007; Vaillancourt et al., 2007; Matsuda et al., 2009; Kurniawan et al., 2010; Fiehler et al., 2011; Hong and Jang, 2011; Martin et al., 2011; Neely et al., 2011; Glover et al., 2012; Makuuchi et al., 2012; Nathan et al., 2012b; Begliomini et al., 2014; Fabbri et al., 2014, 2016; Plata et al., 2014; Monaco et al., 2015; Leo et al., 2016; Przybylski and Króliczak, 2017; Ariani et al., 2018; Cavina-Pratesi et al., 2018; Styrkowiec et al., 2019; Marneweck and Grafton, 2020; Sulpizio et al., 2020; Bencivenga et al., 2021; Errante et al., 2021b; Michalowski et al., 2022). These results must be interpreted in light of several methodological and biological factors: Milner et al., for example, reported that younger participants showed greater deactivation of ipsilateral M1 during hand grip compared to elderly volunteers in both dominant and non-dominant limbs (Milner et al., 2007).
The parietal lobe comprehends areas specialized in encoding and modulating reach- and grasp-related fine hand movements: human medial parietal sulcus (mIPS) and superior parieto-occipital cortex (SPOC) have been proposed as homologs for V6 area complex found in macaques (Pitzalis et al., 2013). Recent studies confirmed the specific role of SPOC area in visually-aided reaching movements (Gallivan et al., 2009; Cavina-Pratesi et al., 2010): its activation during reaching subserves subject hand preferences and stimulus location in peripersonal space. Indeed, left-handers commonly employ both hands when asked to reach and grasp a target object, while right-handers showed a segregation toward a dominant hand use. This preference is also extended to the peripersonal workspace within range of action: right-handers respond with higher SPOC activation when the object is presented within range of their dominant limb, while left-handers exhibit response to objects within range of both sides. Irrespective of hand preference, bilateral SPOC and left precuneus showed visual field polarization, activating most when the object was presented in the inferior visual field. The anterior precuneus (aPCu) is significantly activated during both visually-aid and reaching in the dark, suggesting it does not act as an associative visual area but modulates both visuomotor and sensorimotor input transformation for reaching execution (Filimon, 2010; Gatti et al., 2015). Within this dorsomedial parietal region, the presentation of affordable objects in different visual field sectors elicit reproducible selective activations, not discordant from what is observed in animals: mIPS—which is strongly interconnected with caudal PMd through the dorsomedial stream, is more activated during object reaching execution within the central visual field, while a wider bilateral perieto-occipital junction (POJ) area is elicited during peripheral vision engagement. The latter showed selective connectivity to the rostral PMd (Prado et al., 2005).
In homology with animal models, ventrolateral parietal areas are involved in grasping encoding and performance, namely the rostral part of the lateral bank of IPS (aIPS) and the supramarginal gyrus (SMG; Frey et al., 2005; Culham and Valyear, 2006; Filimon, 2010). An additional area in the superior parietal lobule (SPL) seems related to grasping encoding but overlaps with SPOC, precuneus and mIPS areas. All these share activation signals with PMd in fMRI studies (Tunik et al., 2008; Gallivan et al., 2011b; Fabbri et al., 2014) and co-activate during complex 3D object haptic manipulation (either with and without proper grasping task), together with the right SPL, aIPS, anterior SMG (aSMG) and area SII in the parietal operculum (Jancke, 2001). The parietal opercular region comprehends peculiar associative somatosensory areas, namely OP1 and OP4, as extensive components of the “human grasping/praxis network” (Eickhoff et al., 2006, 2007). According to fMRI data, OP1 and OP4 are concurrently activated during tactile hand stimulation, motor execution and are related to fine object manipulation for target recognition with and without visual aid (Eickhoff et al., 2007; Burton et al., 2008). Further TMS evidence confirmed their involvement in haptic working memory during object identification in the darkness and grasping motor scheme programming. In light of the previous evidence, it has been proposed that OP1 and OP4 are the human homologs for parietal area SII in macaques (human SII).
A connectivity gradient is present in the parietal areas related to the object manipulation network: more dorsal cortical areas – showing higher connectivity with posterior visual areas – might be involved in special processing of visual input integration for action planning when the target object is presented outside the range of action. On the contrary, parieto-ventral areas activation might imply remapping of motor behaviors encoding peripersonal space affordance depending on other than visual sensorial feedback, confirming the consistency with the lateral grasping network model described in monkeys and its dual streams partial segregation (Gallivan et al., 2011a; Renzi et al., 2013; Rossit et al., 2013; Monaco et al., 2017).
The cortico-cortical connections between frontal nodes and PPC within the network consist of long-range association bundles, namely the superior longitudinal fasciculus (SLF) and the arcuate fasciculus (AF) bilaterally. Both injective tracers in monkeys and non-invasive imaging techniques in both species described the anatomical substrate of this densely interconnected network. Probabilistic DTI tractography, among all, permitted an accurate virtual definition of the dorsal stream bundles anatomy, comprehending the classification of its subdivisions (SLF I, II, III) in the living human brain. According to Martino et al., SLF can be anatomically split into three independent fascicles: SLF I connects the superior frontal gyrus and anterior cingulate cortex to SPL and precuneus posteriorly (Martino et al., 2013). SLF II binds the posterior part of the superior and middle frontal gyri and the caudal part of IPL (aIPS and AG). Finally, SLF III connects the inferior frontal gyrus (IFG-BA44, comprehending vPM) to aIPS and IPL. In the interest of simplification, it might be assumed that SLF I represents the cortico-cortical connections of the so-called dorsomedial frontoparietal pathway.
In
留言 (0)