Background
KCNQ2 mutations can lead to a range of heterogeneous early-onset epileptic syndromes, from self-limited familial neonatal epilepsy to severe neonatal-onset developmental and epileptic encephalopathy. Additional features may include dystonia, spasticity, cortical visual impairment, abnormal eye movements, or feeding intolerance (Morrison-Levy et al., 2021). KCNQ2 is one among the genes coding for KCNQ M-current potassium channels that assemble as heteromers, controlling adaptation of neuronal firing and spike frequency across the brain (Delmas and Brown, 2005; Soh et al., 2022). The overall incidence of KCNQ2-related disorders is estimated at 2.93–3.59 per 100 000 births (López-Rivera et al., 2020). Functional outcome is highly variable with different patterns of seizure activity (some subsequently affecting brain maturation) on the one hand, and altered neurodevelopment on the other, independently of epilepsy. Moreover, different mutations of the KCNQ2 gene can cause the same phenotype (Milh et al., 2016). In this context of heterogeneous genotype/phenotype relationships, the spectrum of KCNQ2-related neurodevelopment extends from normal cognition to severe motor, behavioral and social impairment (Symonds et al., 2019). However, although the available literature draws an increasingly clearer picture of the various epileptic syndromes and developmental profiles, there is a need to improve our understanding of the socio-behavioral aspects of the disease to inform and prepare families and caregivers. This is especially important in the long-term as seizure frequency usually declines or recedes after childhood, while a high frequency of autistic features and mild to severe intellectual disability (ID) are reported among adults with KCNQ2 encephalopathy (Boets et al., 2022).
Case presentation
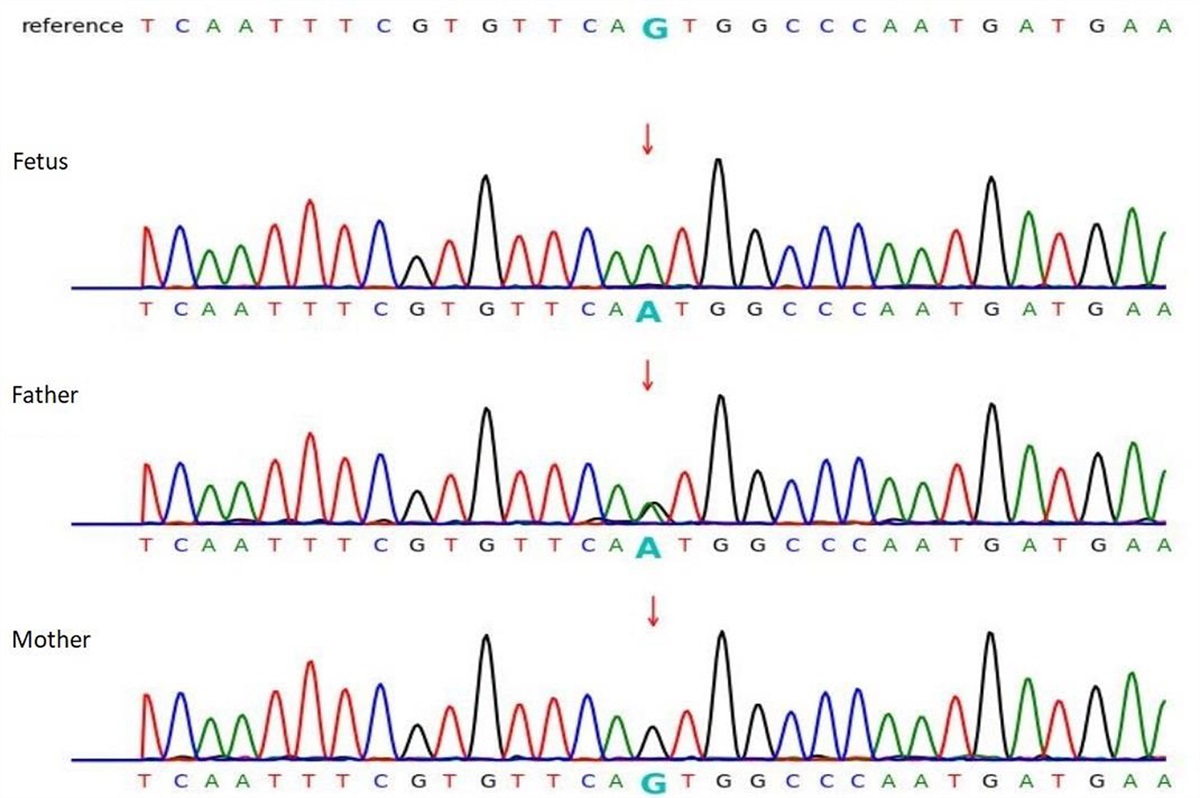
We describe the clinical trajectory of a 5-year-old boy who presented at three months with a cluster of sudden-onset generalized tonic-clonic seizures, over a period of 10 days, efficiently treated with sodium valproate 20 mg/kg for 15 months with full recovery and no further antiepileptic medication required. His father reported similar early-onset generalized tonic-clonic seizures during childhood in the first weeks after birth, efficiently treated with antiepileptics, and followed by a typical neurodevelopment. An investigation in the family revealed that a KCNQ2 mutation was already known on the paternal grandfather’s side. Further contacting the estranged part of the family on this side, the parents learned that many of the family members also presented with various levels of neurodevelopmental delays. At 5 months, a multiplex ligation-dependent probe amplification confirmed the familial deletion c.402delC (p.IIe134Metfs*37, NM_172107.2) in a heterozygous state, in the exon 3 of the KCNQ2 gene. At age 2, psychomotor regression occurred: he became silent, social response could not be elicited anymore, and he developed an important food selectivity to certain colors and textures. While a diagnosis of KCNQ2-related encephalopathy was retained at the time, there had been no formal assessment of the autistic traits. Educational help was provided with a carer dedicated to him in kindergarten but without autism-specific accommodations. At age four, because of persistent behavioral rigidity and disabling sensory particularities, the parents reached out to a KCNQ2 patient association, who subsequently referred them to our specialist autism outpatient unit, where the patient underwent an extensive neuropsychiatric assessment. We received him at age 5. He measured 1.06 m for 17.3 kg, with 52 cm head circumference. General physical and neurological examination was unremarkable. Socially, the patient gave initially no attention to adults, had no joint attention, was avoidant, but allowed a limited interaction after a period of habituation. He demonstrated hetero-aggressivity when frustrated. Our extensive assessment is described in Table 1. It led to a diagnosis of autism spectrum disorder (ASD) and ID with severe delay in cognitive, adaptive abilities, and speech (less than 1 year of equivalent developmental age). The confirmation of the ASD diagnosis provided the patient with an ASD-specific schooling orientation in anticipation of primary school. The assessment of his level of ID helped the various caregivers find the most appropriate rehabilitation or adaptive techniques for his intellectual age.
Table 1 -
Neuropsychiatric and psychological assessment
Domain
Behavior
Sensoriality
Social interaction
Level of development
Adaptive behavior
Scales
ADHD rating scale: 38
Aberrant behavior checklist: 165
Repetitive behavior scale: 63
Dunn sensory profile:
Touch processing: 11
Olfactory procession: 11
Auditory filtering: 15
Visual processing: 16
Energy: 25
Social responsiveness scale: 118, T = 79
Autism diagnostic interview-revised
- Social interaction: 31
- Communication (non-verbal): 14
- Restricted, repetitive behaviors: 16
- Onset before 36 months: 5
Autism diagnostic observation schedule-2
- Social affect: 19
- Restricted, repetitive behaviors: 6
- Total: 25
- Comparison score: 0
Mullen scale (in months)
Visual perception: 14
Fine motor skills: 22
Receptive speech: 2
Expressive speech: 9
Vineland scale (percentiles)
Communication < 1%
Daily life < 1%
Socialization < 1%
Motricity < 1%
Standard result < 1%
ADHD: attention deficit and hyperactivity disorder: cut-off = 28. Social responsiveness scale: cut-off = T > 60. Autism diagnostic interview revised (ADI-R): social interaction cut-off = 10; communication non-verbal cut-off = 7; restricted, repetitive behaviors cut-off = 3, onset before 36 months cut-off = 1. Autism diagnostic observation schedule (ADOS): total cut-off = 7.
Discussion
KCNQ2-related disorders are usually diagnosed in the neonatal period, as the vast majority of patients start with stormy epilepsy, which begins in the first week of life (Weckhuysen et al., 2012; Milh et al., 2013). This mode of onset is quite similar in both self-limited and severe forms. The present report indicates that this specific mutation seems to lead to a relatively late epilepsy onset for a disorder related to KCNQ2, with self-limiting epilepsy that no longer persists. Epilepsy and neurodevelopmental disorders seem thus to develop independently. Despite the deletion of an exon, we do not know whether it resulted in a truncated protein or a complete loss of function, although a single base deletion in exon 3 has been shown to lead to a frameshift and the synthesis of a truncated protein (Lauritano et al., 2019). For missense mutations, the correlation between severity and genotype is less straightforward than for truncating and stop mutations (Soldovieri et al., 2014). Most of the latter lead to self-limited forms with or without slight developmental disorder, generally leading to mild cognitive impairment. Missense mutations are associated with a much wider range of phenotypes (Weckhuysen et al., 2012). This heterogeneity is partly related to the biological effect of the mutation. Indeed, in some cases, there is a relationship between the impact of the mutation on the IM current and the intensity of the neurodevelopmental disorder (Miceli et al., 2013). In other cases, the severity seems to be related to an abnormal channel distribution on the neuronal membrane (Abidi et al., 2015). These functional consequences exert a dominant negative effect. More recently, gain-of-function mutations have been described. Epilepsy either is delayed or absent, and the neurodevelopmental disorder is very variable with a high prevalence of ASDs (Mulkey et al., 2017). Interestingly, phenotypes associated with recurrent mutations have been described in detail. Despite the similarity of the KCNQ2 mutation, the phenotype may be extremely variable, indicating that factors other than those related to the KCNQ2 mutation may be involved in the phenotype of patients (Miceli et al., 2022).
The study of patients and models of KCNQ2 opens fascinating perspectives for the understanding of the link between visible epilepsy and developmental disorders. This link is not causal. In animal models, cognitive impairment is not prevented by treating seizures, but by correcting the mutation or its effects before it becomes symptomatic (Peters et al., 2005; Marguet et al., 2015). In humans, the link between epileptic activity and cognitive profile has not yet been proven (Berg et al., 2022). Recently, the study of an animal model of severe KCNQ2-related disorder showed that the impact of the mutation on the IM current was limited in time. Indeed, the IM current was abnormal until the age of weaning, causing neuronal hyperexcitability. After weaning, the current normalized, the neuronal hyperexcitability disappeared, while the seizures persisted and the cognitive disorders became measurable (Biba-Maazou et al., 2022). Overall, current data tend to show: (1) that the disorder induced by the mutation occurs earlier than the seizures (which are a late reflection of it); (2) that treating the seizures will probably not be sufficient to allow normal development; and (3) that at the stage of cognitive and/or autistic disorders, the target is no longer IM, but late co-consequences of the earlier dysfunction of IM. The study of the effects of the mutation on the early stages of development (establishment of protomaps, first neuronal assemblies, first network activities; impact on critical periods of development…) opens an unexplored field to better understand the emergence of the autistic disorder in this condition and in general (Cossart and Garel, 2022). When mutations are associated with highly variable phenotypes between a parent and child, the possibility that the parent carries a somatic mosaic has been raised (Milh et al., 2015). Here, this possibility is excluded because of the transmission over several generations.
This patient’s history is relevant for psychiatric and social reasons as well. A main issue of care in this case was the diagnostic delay. First, the molecular diagnosis was delayed by the fact that the existence of a familial mutation was initially unknown to the parents. This raises the question of guilt and stigma around genetic diagnoses and the reasons not to disclose them to other members of the family. Thus, it has been reported that people with epilepsy could experience genetic information as harmful for a variety of reasons from fear of health insurance eligibility and loss of coverage for their children, to how this may change one’s view of reproduction, or significant guilt in parents who may have transmitted the genetic risk (Shostak et al., 2011). Thus, felt stigma was found to be increased by genetic causal attribution of epilepsy (Sabatello et al., 2015; Garofalo et al., 2019). A specialist genetic counseling should therefore systematically be offered to families, especially as the diagnosis itself provides many benefits not only in terms of early healthcare but also as a reducer of self-blame and uncertainty. By improving their understanding of the disease, patients can regard it as something they are not responsible for in terms of lifestyle (Shostak et al., 2011). Second, there was a delay in the subsequent diagnosis of ASD. ASD is frequently associated with KCNQ2-related diseases and is directly attributable to the mutation (Cheng et al., 2021; Siracusano et al., 2022). Early screening is an important goal as it opens the way to specialized ASD-oriented care, such as applied behavior analysis therapy, while allowing families to benefit from adapted schooling. In the context of precision medicine, these interventions are crucial to assess and improve the quality of life of patients with developmental and epileptic encephalopathies, which require personalized, individually based assessment tools of clinical outcome and communication abilities (Berg et al., 2022; Cohen et al., 2022).
Acknowledgements
We thank the patient and his parents for their support. We thank the referring physicians for their cooperation. All authors have contributed to this study in data collection and analysis, as well as manuscript preparation.
Patient consent for publication: Parental/guardian(s) consent obtained.
Conflicts of interest
There are no conflicts of interest.
References
Abidi A, Devaux JJ, Molinari F, Alcaraz G, Michon F-X, Sutera-Sardo J, et al. (2015). A recurrent KCNQ2 pore mutation causing early onset epileptic encephalopathy has a moderate effect on M current but alters subcellular localization of Kv7 channels. Neurobiol Dis 80:80–92.
Berg AT, Kaat AJ, Zelko F, Wilkening G (2022). Rare diseases – rare outcomes: Assessing communication abilities for the developmental and epileptic encephalopathies. Epilepsy Behav 128:108586.
Biba-Maazou N, Becq H, Pallesi-Pocachard E, Sarno S, Granjeaud S, Montheil A, et al. (2022). Time-limited alterations in cortical activity of a knock-in mouse model of KCNQ2-related developmental and epileptic encephalopathy. J Physiol 600:2429–2460.
Boets S, Johannesen KM, Destree A, Manti F, Ramantani G, Lesca G, et al. (2022). Adult phenotype of KCNQ2 encephalopathy. J Med Genet 59:528–535.
Cheng P, Qiu Z, Du Y (2021). Potassium channels and autism spectrum disorder: An overview. Int J Dev Neurosci 81:479–491.
Cohen SR, Helbig I, Kaufman MC, Schust Myers L, Conway L, Helbig KL (2022). Caregiver assessment of quality of life in individuals with genetic developmental and epileptic encephalopathies. Develop Med Child Neurol 64:957–964.
Cossart R, Garel S (2022). Step by step: cells with multiple functions in cortical circuit assembly. Nat Rev Neurosci 23:395–410.
Delmas P, Brown DA (2005). Pathways modulating neural KCNQ/M (Kv7) potassium channels. Nat Rev Neurosci 6:850–862.
Garofalo DC, Sorge ST, Hesdorffer DC, Winawer MR, Phelan JC, Chung WK, et al. (2019). Genetic attribution and perceived impact of epilepsy in multiplex epilepsy families. Epilepsia 60:2286–2293.
Lauritano A, Moutton S, Longobardi E, Tran Mau-Them F, Laudati G, Nappi P, et al. (2019). A novel homozygous KCNQ3 loss-of-function variant causes non-syndromic intellectual disability and neonatal-onset pharmacodependent epilepsy. Epilepsia Open 4:464–475.
López-Rivera JA, Pérez-Palma E, Symonds J, Lindy AS, McKnight DA, Leu C, et al. (2020). A catalogue of new incidence estimates of monogenic neurodevelopmental disorders caused by de novo variants. Brain 143:1099–1105.
Marguet SL, Le-Schulte VTQ, Merseburg A, Neu A, Eichler R, Jakovcevski I, et al. (2015). Treatment during a vulnerable developmental period rescues a genetic epilepsy. Nat Med 21:1436–1444.
Miceli F, Soldovieri MV, Ambrosino P, Barrese V, Migliore M, Cilio MR, et al. (2013). Genotype–phenotype correlations in neonatal epilepsies caused by mutations in the voltage sensor of Kv72 potassium channel subunits. Proc Natl Acad Sci USA 110:4386–4391.
Miceli F, Millevert C, Soldovieri MV, Mosca I, Ambrosino P, Carotenuto L, et al. (2022). KCNQ2 R144 variants cause neurodevelopmental disability with language impairment and autistic features without neonatal seizures through a gain-of-function mechanism. eBioMedicine 81:104130.
Milh M, Boutry-Kryza N, Sutera-Sardo J, Mignot C, Auvin S, Lacoste C, et al. (2013). Similar early characteristics but variable neurological outcome of patients with a de novo mutation of KCNQ2. Orphanet J Rare Dis 8:80.
Milh M, Lacoste C, Cacciagli P, Abidi A, Sutera-Sardo J, Tzelepis I, et al. (2015). Variable clinical expression in patients with mosaicism for KCNQ2 mutations. Am J Med Genet A 167A:2314–2318.
Milh M, Cacciagli P, Ravix C, Badens C, Lépine A, Villeneuve N, et al. (2016). Severe neonatal seizures: From molecular diagnosis to precision therapy? Rev Neurol (Paris) 172:171–173.
Morrison-Levy N, Borlot F, Jain P, Whitney R (2021). Early-onset developmental and epileptic encephalopathies of infancy: an overview of the genetic basis and clinical features. Pediatr Neurol 116:85–94.
Mulkey SB, Ben-Zeev B, Nicolai J, Carroll JL, Grønborg S, Jiang Y-H, et al. (2017). Neonatal nonepileptic myoclonus is a prominent clinical feature of KCNQ2 gain-of-function variants R201C and R201H. Epilepsia 58:436–445.
Peters HC, Hu H, Pongs O, Storm JF, Isbrandt D (2005). Conditional transgenic suppression of M channels in mouse brain reveals functions in neuronal excitability, resonance and behavior. Nat Neurosci 8:51–60.
Sabatello M, Phelan JC, Hesdorffer DC, Shostak S, Goldsmith J, Sorge ST, et al. (2015). Genetic causal attribution of epilepsy and its implications for felt stigma. Epilepsia 56:1542–1550.
Shostak S, Zarhin D, Ottman R (2011). What’s at stake? Genetic information from the perspective of people with epilepsy and their family members. Soc Sci Med 73:645–654. doi:
10.1016/j.socscimed.2011.06.043.
Siracusano M, Marcovecchio C, Riccioni A, Dante C, Mazzone L (2022). Autism spectrum disorder and a De Novo Kcnq2 gene mutation: a case report. Pediatr Rep 14:200–206.
Soh H, Springer K, Doci K, Balsbaugh JL, Tzingounis AV (2022). KCNQ2 and KCNQ5 form heteromeric channels independent of KCNQ3. Proc Natl Acad Sci USA 119:e2117640119.
Soldovieri MV, Boutry-Kryza N, Milh M, Doummar D, Heron B, Bourel E, et al. (2014). Novel KCNQ2 and KCNQ3 mutations in a large cohort of families with benign neonatal epilepsy: first evidence for an altered channel regulation by syntaxin-1A. Hum Mutat 35:356–367.
Symonds JD, Zuberi SM, Stewart K, McLellan A, O'Regan M, MacLeod S, et al. (2019). Incidence and phenotypes of childhood-onset genetic epilepsies: a prospective population-based national cohort. Brain 142:2303–2318.
Weckhuysen S, Mandelstam S, Suls A, Audenaert D, Deconinck T, Claes LRF, et al. (2012). KCNQ2 encephalopathy: emerging phenotype of a neonatal epileptic encephalopathy. Ann Neurol 71:15–25.






留言 (0)