
記住我



Precocious puberty refers to the development of secondary sexual characteristics before the age of 8 years in girls and before the age of 9 years in boys (1). Precocious puberty can lead to accelerated skeletal maturation, advanced bone age, and early epiphyseal closure, all of which can impact final adult height. In addition, it may result in psychological issues or abnormal social behavior (2, 3). The long-term health consequences associated with early menarche include increased risk of obesity, type 2 diabetes, estrogen-dependent cancers, and cardiovascular events (3). It has been estimated that approximately one in five thousand children worldwide are affected by precocious puberty, with a markedly higher occurrence in girls than in boys, ranging from five to ten times more (2). Generally, girls are more likely to experience idiopathic precocious puberty, whereas approximately half of boys with precocious puberty have an identifiable cause. Thus, we focus on only girls in the present study (4). Currently, the primary recommended clinical treatment for precocious puberty is the utilization of gonadotropin-releasing hormone analog (GnRHa) (2). Although this treatment is considered safe and generally well -tolerated in children and adolescents (5), the GnRHa treatment cycle usually lasts longer than two years. Furthermore, the higher doses and considerable cost often impose significant psychological and financial burdens on the children’s family (6). Therefore, the prevention of precocious puberty has become an urgent public health goal.
Despite the etiology being incompletely understood, precocious puberty is thought to be associated with obesity, genes, lifestyle habits, environmental endocrine disruptors, and other factors (7, 8). Nutritional status and body fat mass are particularly vital factors contributing to precocious puberty (8, 9). Several research has indicated that obesity is closely linked to precocious puberty (8, 10). Therefore, preventing the occurrence of obesity has become a novel treatment strategy for preventing precocious puberty in girls. (-)-Epigallocatechin-3-gallate (EGCG) is the most abundant, bioactive, and extensively researched catechin in tea, with numerous studies verifying its efficacy in the prevention of obesity (11–13). Our research group found that EGCG has preventive effects on obesity-related precocious puberty (14), but the specific molecular mechanism remains to be elucidated.
Metabolomics is a comprehensive technique for monitoring the dynamics of endogenous small molecule metabolites and reflecting changes in metabolic pathways throughout the metabolic network in an organism (15). Network pharmacology is a widely utilized approach for determining the mechanism of drugs. High-throughput screening and analysis are employed to predict the complexity and integrality of the interactions between drugs and their targets and diseases and the related pharmacological mechanisms (16). In this study, the mechanism of action by which EGCG affected obesity-related precocious puberty was systematically investigated at the molecular level by integrated metabolomics and network pharmacology approach. The present research attempted to offer a theoretical foundation for further investigation into the treatment of obesity-related precocious puberty.
2 Materials and methods2.1 Metabolomics analysisSerum samples were derived from our prior randomized control trial (NCT03628937). Briefly, researchers assigned six to ten-year-old obese girls to two groups: placebo and EGCG. Twelve weeks of EGCG capsules (200mg, 50% EGCG) were given to girls in the EGCG group (n=18). The placebo group (n=16) received placebo capsules that appeared the same for twelve weeks. Detailed descriptions of the research design have been provided previously (14).
Serum metabolomics were analyzed using a high-performance liquid chromatography-electrospray ionization ion-trap tandem mass spectrometry (LC-ESI-MS/MS) in both positive and negative ion modes. Chromatographic conditions were based on the prior description (17).
Data analysis was carried out utilizing Majorbio Cloud (https://cloud.majorbio.com), based on the description given previously (17). In brief, the overall differences between the EGCG and Placebo groups were determined using orthogonal partial least squares discriminant analysis (OPLS-DA) after data preprocessing and annotation. Variable importance in the projection (VIP) values (>1.0) and statistical analysis (P < 0.05) were used to identify differential metabolites. The Kyoto Encyclopedia of Genes and Genomes (KEGG) and Human Metabolome Database (HMDB) were employed to identify and analyze implicated pathways associated with the metabolites.
2.2 Identification of EGCG targetsThe PubChem database was employed to obtain simplified molecular-input line-entry system information and the 2D structure of EGCG (18), which were uploaded respectively to SwissTargetPrediction and PharmMapper Server for potential EGCG targets prediction (19). DrugBank was also used to identify EGCG targets (20). The UniProt database was utilized to correct all retrieved target names (21).
2.3 Identification of obesity and precocious puberty targetsTargets related to obesity and precocious puberty were found in three databases, including GeneCards (22), DisGeNET (23) and the Comparative Toxicogenomic Database (CTD) (24), using the key words “precocious puberty”, “early puberty” and “obesity”. To increase the credibility of analysis, this study included targets with a relevance score >1 in GeneCards, a gene-disease score >0.1 in DisGeNET, and an inference score >50 in CTD. We generated a Venn diagram through a VENNY 2.1 tool to identify common targets of EGCG, precocious puberty, and obesity.
2.4 The protein–protein interaction (PPI) network constructionPPI network was generated via uploading the intersecting targets of EGCG, precocious puberty, and obesity to the STRING database (25). Network visualization was conducted utilizing Cytoscape 3.7.2. software. To evaluate network topology, the CytoNCA plug-in was used, and the hub target genes were screened based on degree scores.
2.5 Gene Ontology (GO) and KEGG enrichment analysesGO and KEGG pathway enrichment analyses were completed using the ClusterProfiler package of R 4.0.3 to elucidate the role of the common targets in gene function and signaling pathways (26). The GO analysis comprised of three primary components, including biological process (BP), cellular component (CC), and molecular function (MF). In order to guarantee the accuracy of the enrichment outcomes, the Benjamin-Hochberg method was employed for multiple testing correction. The adjusted P value < 0.05 was utilized as a threshold of significance for the enriched GO and KEGG terms for target genes. The GO enrichment analysis results were visualized through the online mapping platform Bioinformatics, and significant KEGG pathways (P<0.05) were visualized with OmicShare Tools.
2.6 Molecular dockingTo validate the aforementioned results, molecular docking analysis was conducted to calculate the affinity of hub target gene products in the network. We selected hub target gene products and EGCG for molecular docking. From the PubChem database, three-dimensional (3D) structures of ligands were obtained (18). The RCSB database was utilized to acquire the 3D structures of receptors (27). Using mgltools_win32_1.5.6 software, ligands and receptors were repaired and saved as PDBQT files. The affinity of docking between EGCG and hub target proteins was determined utilizing AutoDock Vina 1.1.2 software. The molecular docking data were visualized utilizing PyMOL 2.3.
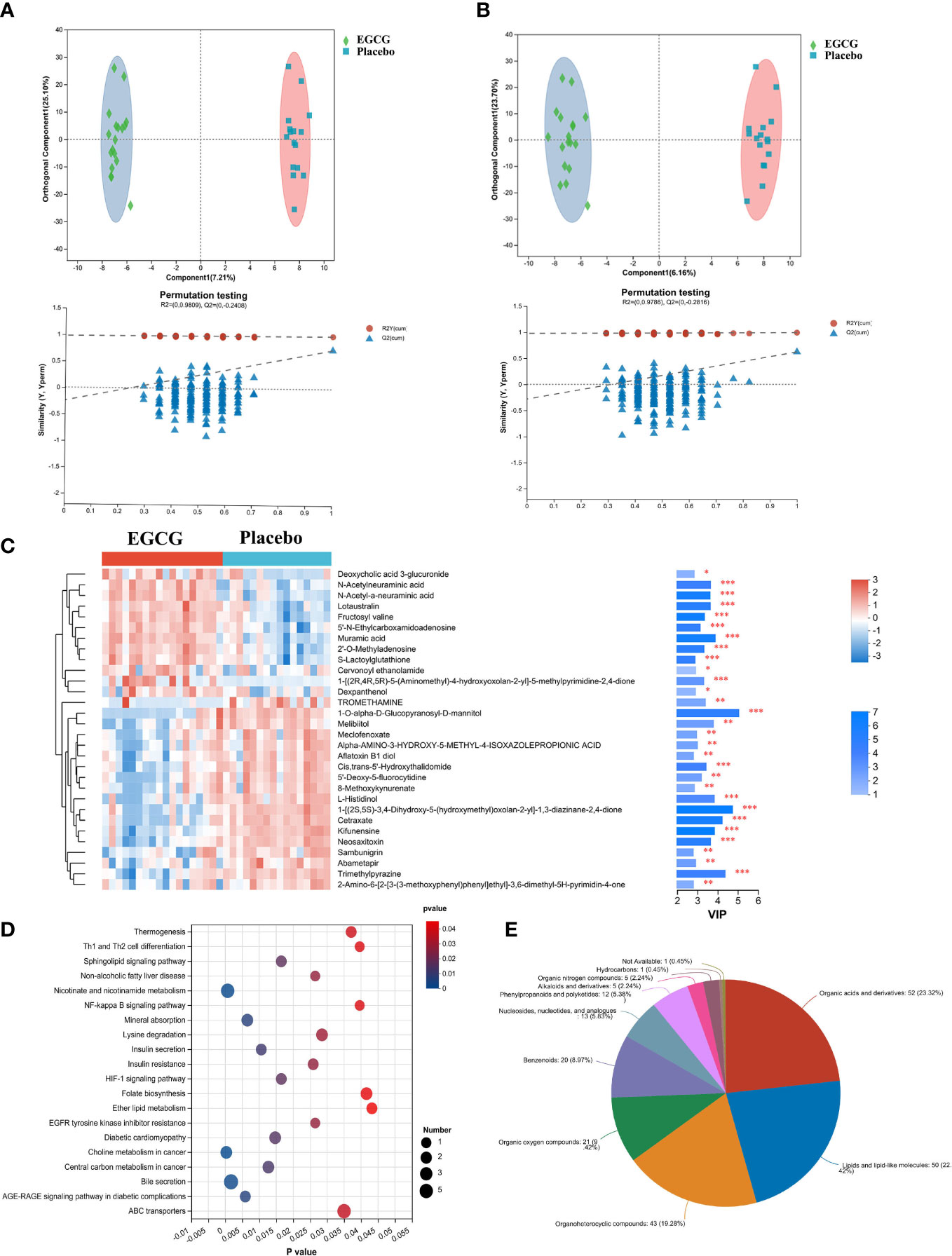
3 Results3.1 EGCG alters the serum metabolomeThis study applied metabolomic analysis to identify endogenous differential metabolites in serum of obese girls following intervention with EGCG. The OPLS-DA analysis revealed that serum samples from the EGCG and placebo groups were clearly separated in both positive and negative modes (Figures 1A, B), indicating significant differences between the two groups. The results of 200 permutations showed that there was no overfitting in the OPLS-DA model. In total, 234 endogenous differential metabolites were screened (Supplementary Table 1), and the top 30 metabolites were depicted in Figure 1C. Figure 1D illustrated endogenous differential metabolites enrichment pathways, mainly involving endocrine pathway (Insulin resistance, Insulin secretion, Non-alcoholic fatty liver disease and AGE-RAGE signaling pathway in diabetic complications), lipid metabolism (Sphingolipid metabolism and Ether lipid metabolism) and signal transduction (NF-kappa B and HIF-1 signaling pathway). With the help of the HMDB database, the differential metabolites between the EGCG and placebo groups were identified and classified according to the HMDB superclass. The majority of these endogenous differential metabolites were organic acids and derivatives (23.32%), and lipids and lipid-like molecules (22.42%) (Figure 1E). Based on these results, EGCG might have a vital regulatory effect on serum lipid metabolism, especially hormonal disorders, in obese girls.
Figure 1 Multivariate statistical analysis of metabolite profiles in serum in obese girls OPLS-DA score plots of the EGCG group and Placebo group in positive mode (A) and negative mode (B). (C) Heatmap of top 30 endogenous differential metabolites between two groups, *P < 0.05, **P < 0.01, ***P < 0.001. (D) KEGG signaling pathway enrichment analysis (P < 0.05). (E) Categories of endogenous differential metabolites.
3.2 Identification of potential targets of EGCG, obesity and precocious pubertyA total of 357 EGCG targets were obtained following the removal of duplicates from the Swiss Target Prediction, PharmMapper, and DrugBank databases (Supplementary Table 2). A total of 3447 targets of obesity were obtained after the merging of results from each database and the removal of duplicates (Supplementary Table 3). Similarly, 3994 targets of precocious puberty were collected by utilizing the key words “precocious puberty” and “early puberty” to search the databases (Supplementary Table 4).
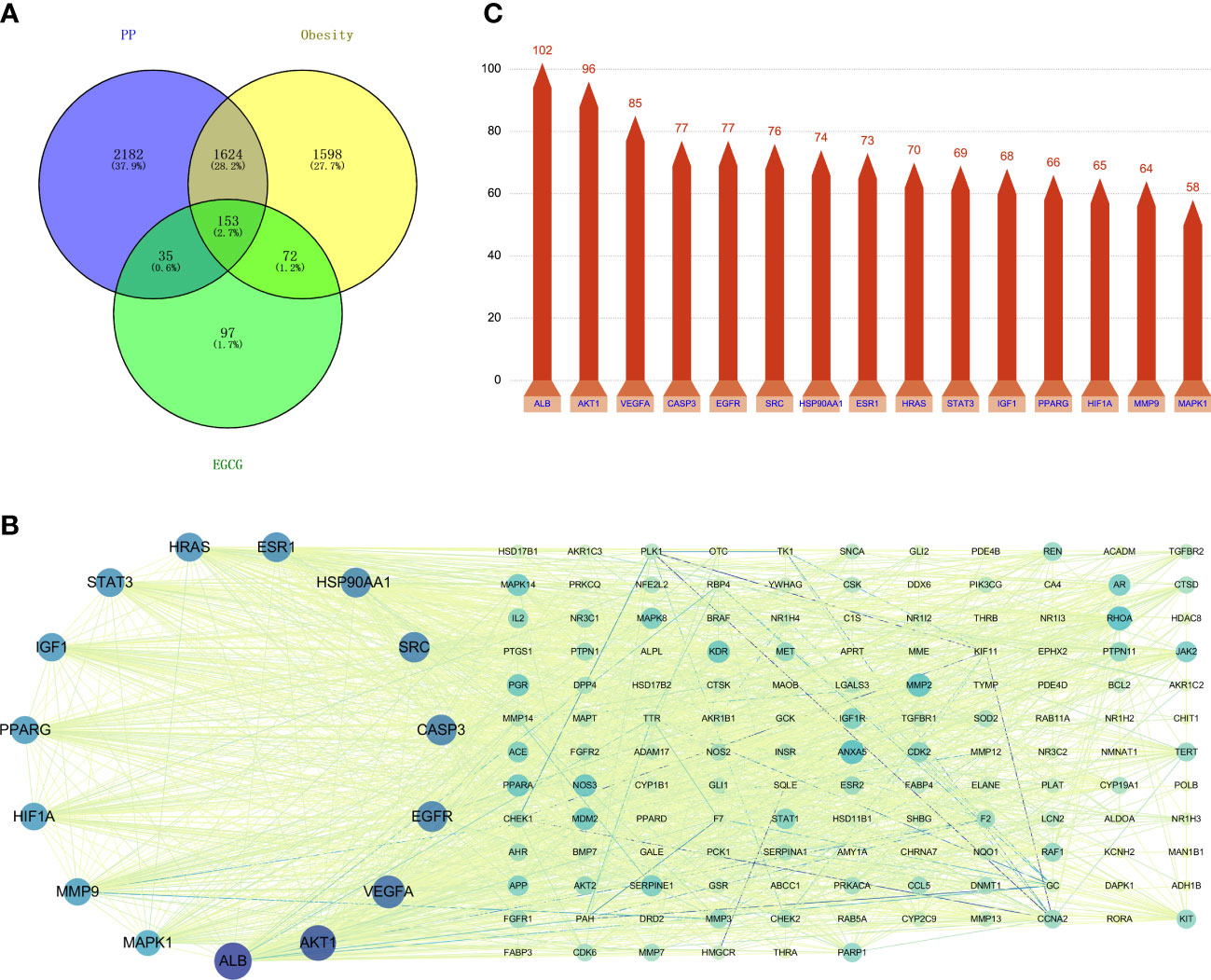
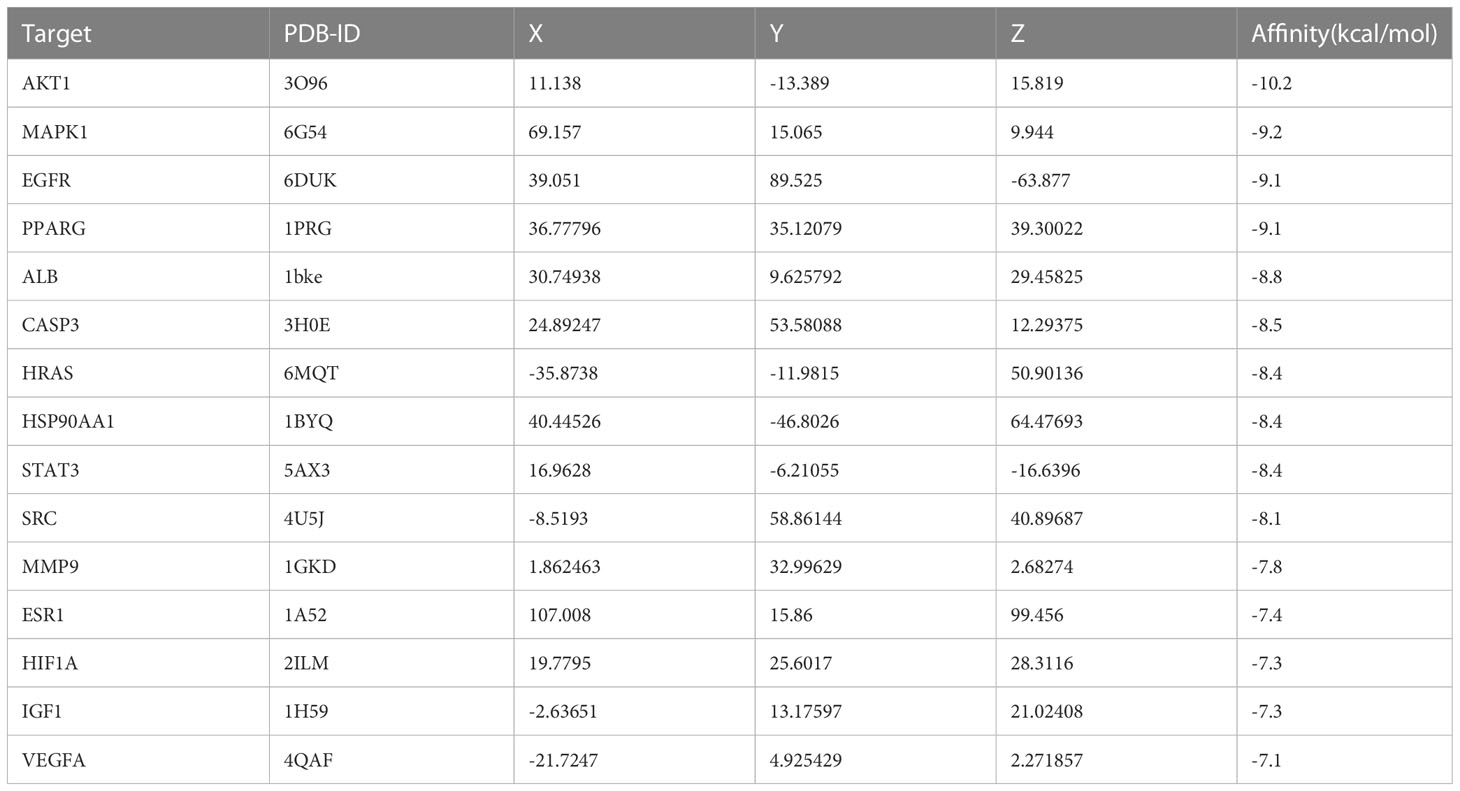
3.3 PPI network of the potential therapeutic targetsAt the intersection of the 357 EGCG targets, 3447 obesity targets and 3994 precocious puberty targets, we identified 153 common targets (Figure 2A, Supplementary Table 4). Subsequently, a PPI network was constructed for these 153 common targets (Figure 2B). A topological analysis was performed to identify hub target genes in this intricate biological network. We determined the top 15 hub target genes according to degree score: ALB, AKT1, VEGFA, CASP3, EGFR, SRC, HSP90AA1, ESR1, HRAS, STAT3, IGF1, PPARG, HIF1A, MMP9, and MAPK1 (Figure 2C). In light of these data, we speculated that EGCG might serve as a vital role in the treatment of obesity and precocious puberty through these 15 hub target genes and their products, which were utilized for the subsequent molecular docking analysis.
Figure 2 An analysis of potential therapeutic targets based on a Venn diagram and a PPI network (A) Venn diagram of the common targets of EGCG, obesity and precocious puberty. (B) Potential therapeutic targets in the PPI network. (C) The bar plot of the protein–protein interaction network.
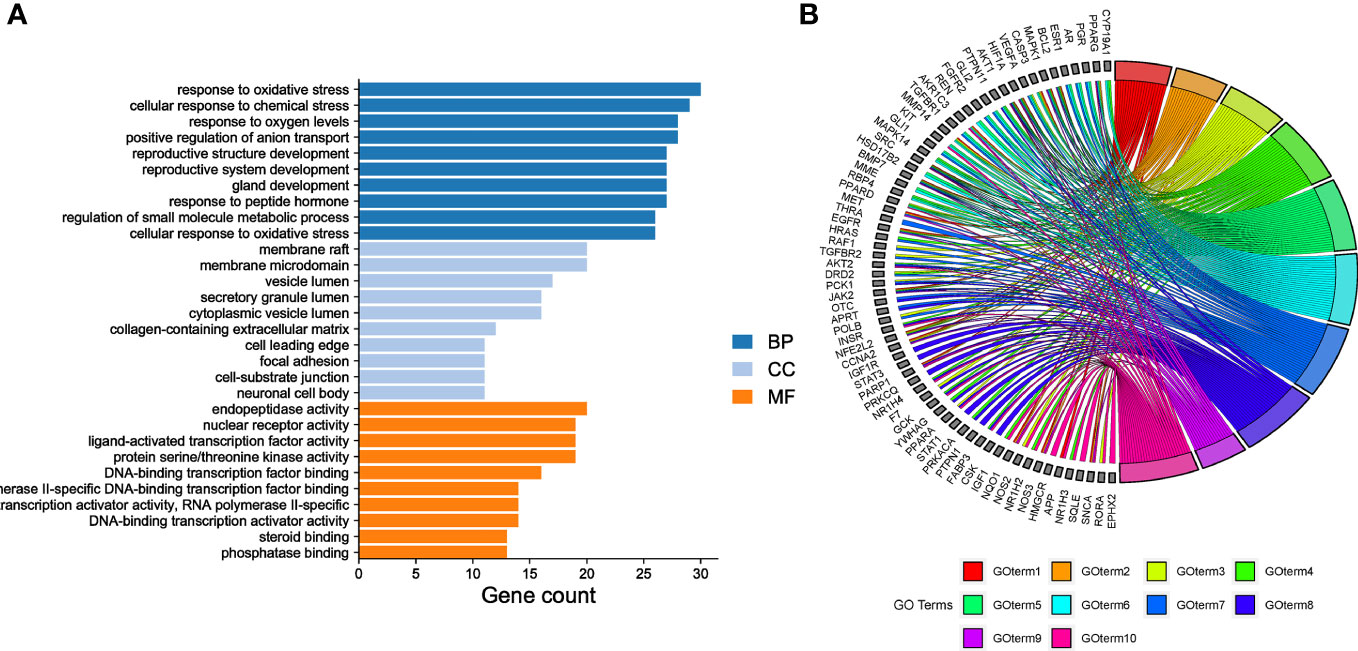
3.3 GO enrichment analysisGO enrichment analysis was conducted to determine the relationships between the 153 common targets and diseases. A total of 2289 BP terms, 51 CC terms, and 163 MF terms were identified (P<0.05). The top ten significantly enriched terms from the three categories were output based on gene count (Figure 3A). The number of genes enriched in the BP category was the highest, indicating that EGCG mainly exerted its anti-obesity and anti-precocious puberty effects by regulating the BP of cells. The mechanisms indicated by the identified terms included the response to oxidative stress, cellular response to chemical stress, response to oxygen levels, positive regulation of anion transport, reproductive structure development, reproductive system development, gland development, response to peptide hormones, regulation of small molecule metabolic processes, and cellular response to oxidative stress. Thirteen of the fifteen hub genes were enriched in the top five enriched BPs, including AKT1, VEGFA, CASP3, EGFR, SRC, ESR1, HRAS, STAT3, IGF1, PPARG, HIF1A, and MAPK1 (Figure 3B). Based on the GO analysis, Figure 3B depicted the top 10 enriched BP terms.
Figure 3 Gene Ontology (GO) enrichment analysis (A) GO enrichment analysis. The top 10 evidently enriched terms in each category. BP, biological process; CC, cellular component; MF, molecular function. (B) The top 10 enriched BP terms.
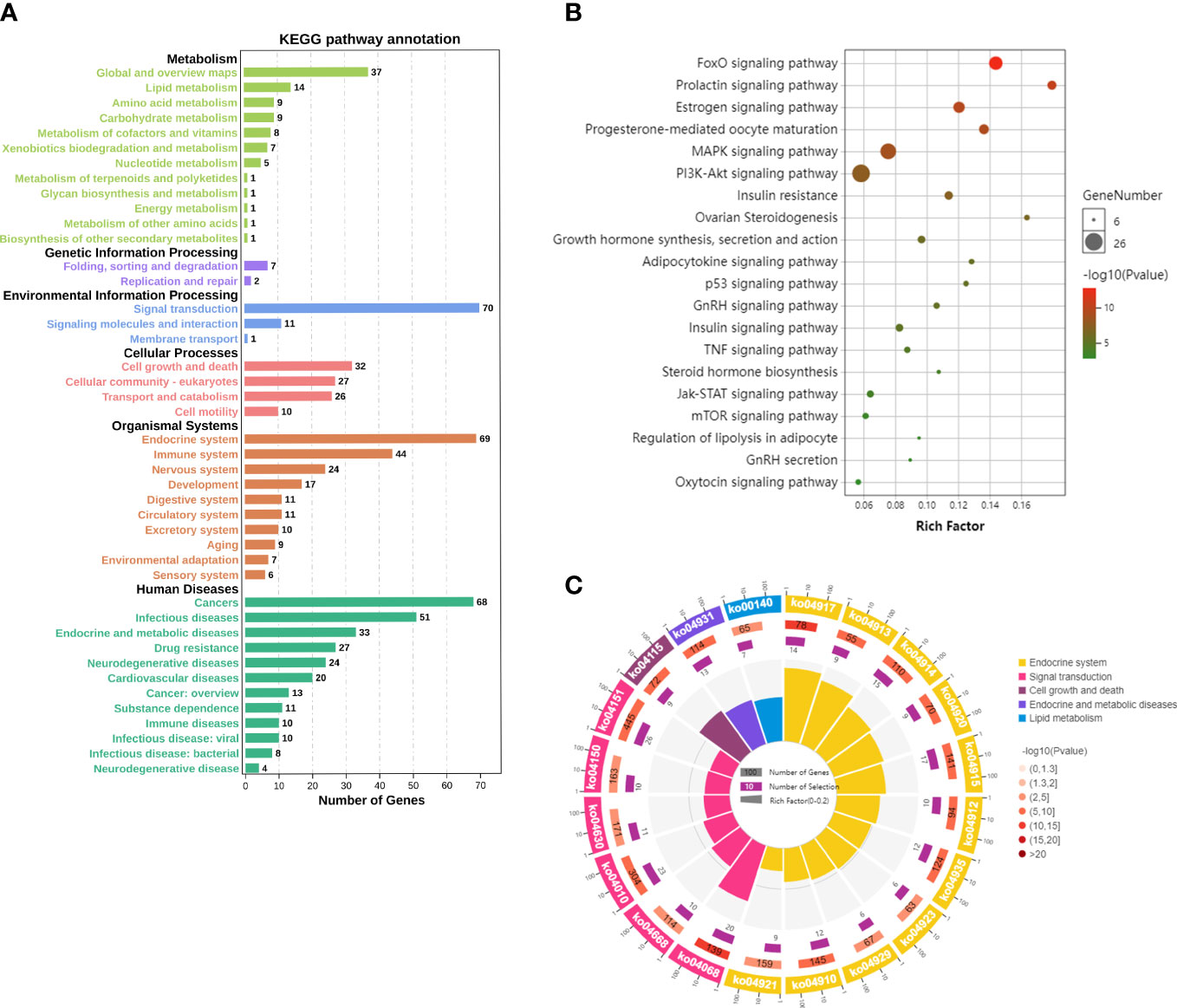
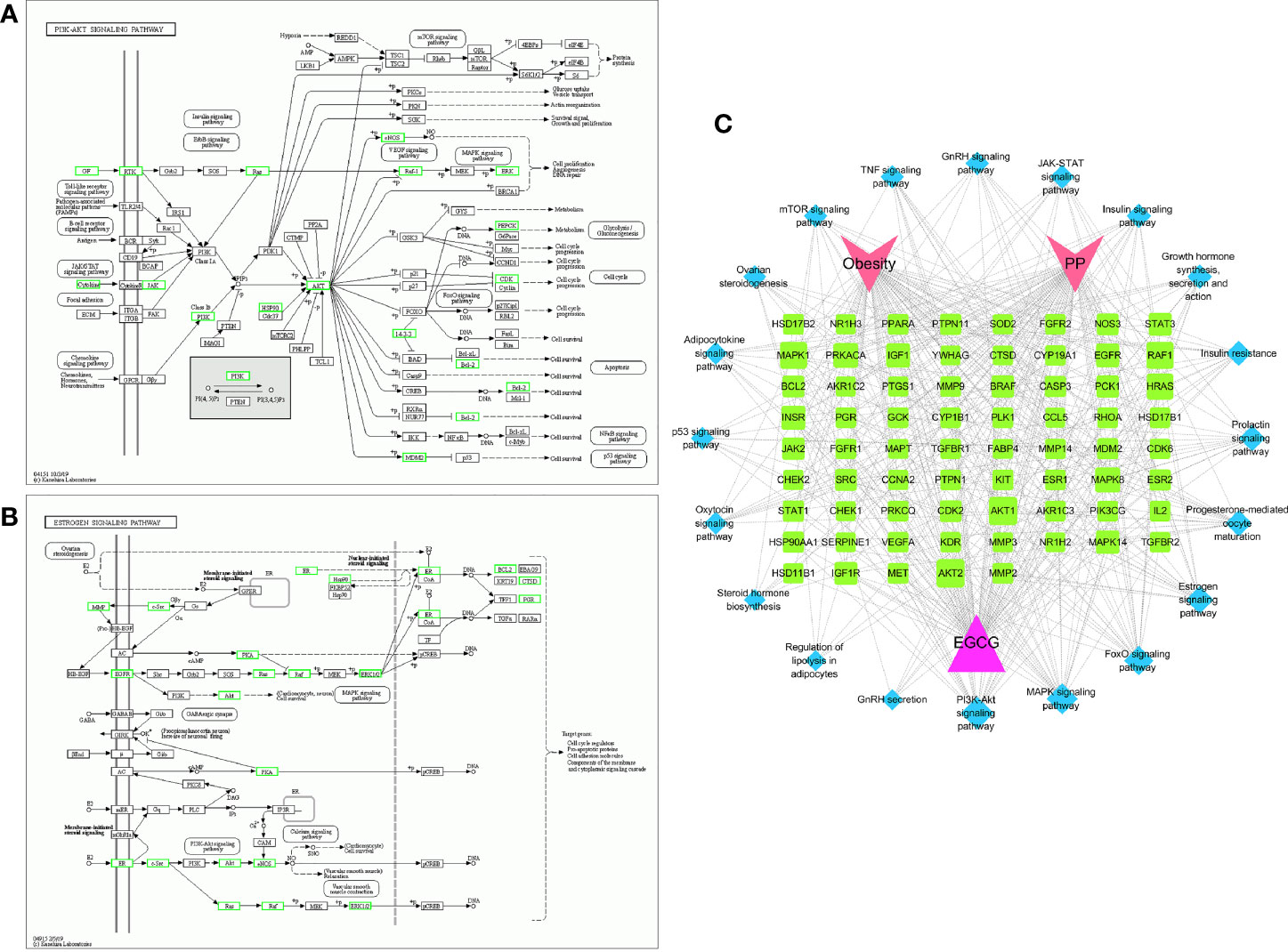
3.4 KEGG enrichment analysisThe mechanism of EGCG in obesity and precocious puberty was further elucidated through KEGG pathway enrichment analysis. A total of 119 pathways with P<0.05 were obtained. The hub genes were predominantly enriched in metabolic pathways, environmental information processes (signal transduction), cellular processes (cell growth and death), and organismal systems (endocrine system) (Figure 4A). Twenty crucial pathways were identified following data screening (Figures 4B, C). The enriched genes were linked to endocrine-related pathways and processes, such as the estrogen signaling pathway, progesterone-mediated oocyte maturation, insulin resistance, and the prolactin signaling pathway. In addition, several of the identified signaling pathways were related to signal transduction, including the forkhead box O (FoxO) signaling pathway, mitogen-activated protein kinase (MAPK) signaling pathway, phosphatidylinositol 3-kinase (PI3K)-protein kinase B (Akt) signaling pathway, and Janus kinase (Jak)-signal transducer and activator of transcription (STAT) signaling pathway. A significant enrichment of the estrogen signaling pathway and PI3K-Akt signaling pathway was observed in the network (Figures 5A, B). Next, the component-target-pathway network diagram was created via Cytoscape 3.7.2 software, as shown in Figure 5C. This diagram demonstrated that 69 of the 153 possible targets were involved in the enrichment of the 20 significant pathways. Additionally, the pathways were interconnected, with certain targets overlapping and interacting with each other. These findings indicated that EGCG could influence multiple interacting pathways to interfere with obesity and precocious puberty.
Figure 4 Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway results (A) Statistical chart of the B-level classification of each pathway. (B) The top 20 significant pathways. Bubble size from large to small indicates the count of potential targets enriched in the pathway in descending order. Bubble color from red to green indicates the -log (p value) in descending order. (C) The top 20 significantly enriched differential pathway circle maps.
Figure 5 Potential therapeutic targets based on evidently enriched pathways The green nodes represent potential therapeutic targets of EGCG obesity and precocious puberty in PI3K-AKT pathway (A) and estrogen pathway (B). (C) EGCG-target-pathway multiple interactive network.
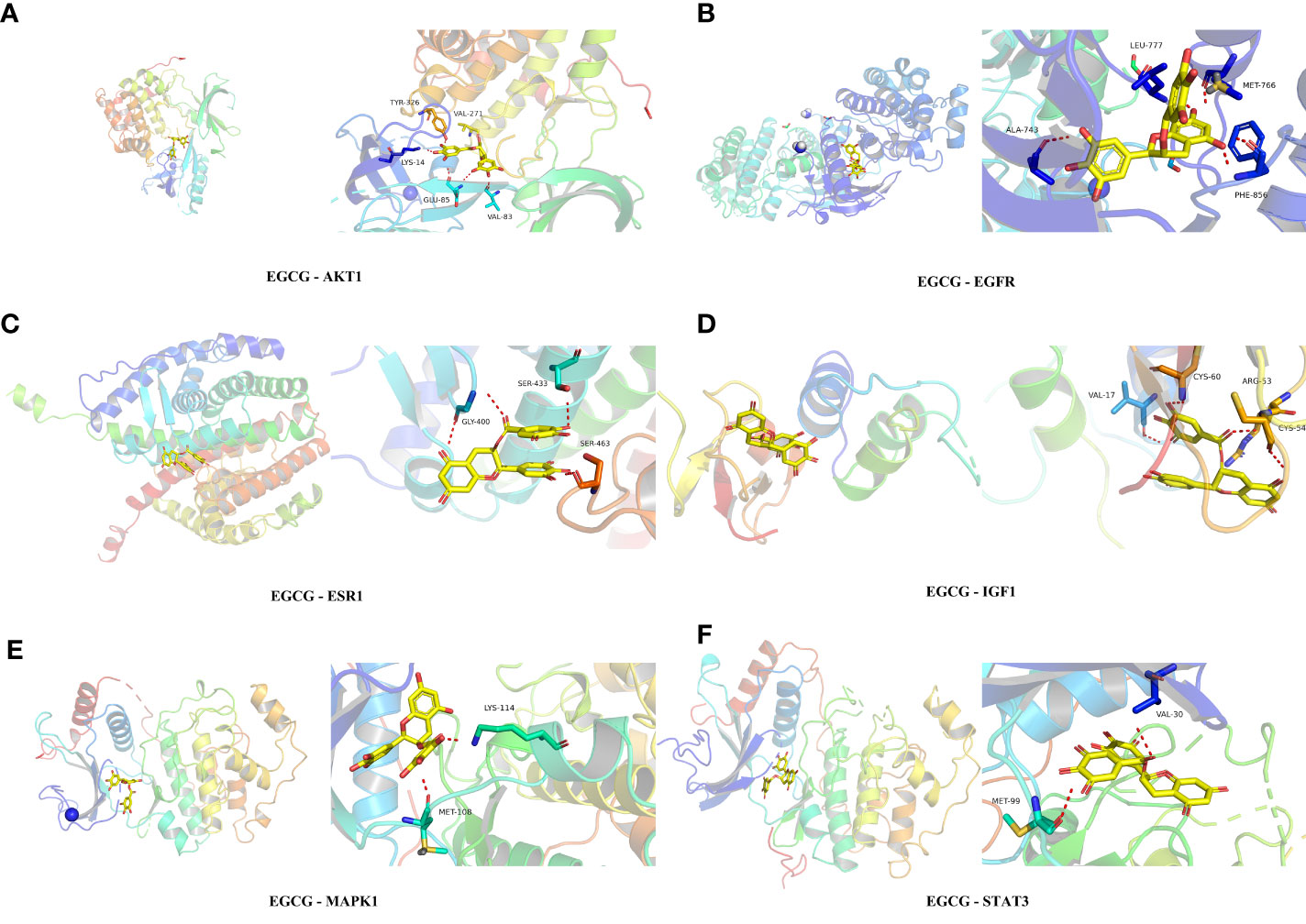
3.5 Molecular docking resultsTable 1 presented the molecular docking outcomes of EGCG to the 15 hub target genes. The binding energies of all the simulations were <−7 kcal/mol, indicating strong binding affinity. Figure 6 demonstrated the binding mode of EGCG with the hub targets, including AKT1, MAPK1, ESR1, EGFR, STAT3, and IGF1. These results suggested that EGCG had strong affinity for the 15 hub target gene products, which may be the primary targets of EGCG in preventing obesity-related precocious puberty.
Table 1 The molecular docking results of EGCG to the 15 hub target gene.
Figure 6 Molecular docking analysis (A) EGCG with AKT1. (B) EGCG with EGFR. (C) EGCG with ESR1. (D) EGCG with IGF1. (E) EGCG with MAPK1. (F) EGCG with STAT3.
4 DiscussionPrecocious puberty is a common endocrine disease that is usually considered to be caused by the premature initiation of the hypothalamic-pituitary-ovarian axis (HPGA), resulting in the premature development of internal and external genitalia (28). Girls with precocious puberty are significantly more inclined to develop breast cancer, cardiovascular disease, and diabetes in adulthood (29). Our previous randomized controlled trial and animal experiments confirmed the significant ability of EGCG to prevent obesity-related precocious puberty (14, 30), but the underlying mechanism remained elusive. Therefore, this study applied integrated metabolomics and network pharmacology approach for the first time to elucidate the mechanism by which EGCG prevented obesity-related precocious puberty and to offer a theoretical foundation for forthcoming research. According to the results of integrated metabolomics and network pharmacology, 6 hub targets, including AKT1, EGFR, ESR1, STAT3, IGF1, and MAPK1, may play significant roles in the effects of EGCG on obesity-related precocious puberty.
The pathogenesis of precocious puberty is related to early initiation of the HPGA, and the pivotal factor in the HPGA initiation is the pulsatile secretion of gonadotropin-releasing hormone (GnRH) (31). Among the top 15 selected hub target genes was AKT1, one of three closely related serine/threonine protein kinases; AKT1 mainly regulates processes such as proliferation, metabolism, and cell survival (32). The phosphatase-binding function of AKT1 is required for IGF-1 and PI3K-mediated adipocyte differentiation, and IGF1 is crucial for preadipocyte proliferation, survival, and differentiation (33). IGF1 is a polypeptide comprised of 70 amino acids, whose function is analogous to that of insulin. IGF1 binds its receptor IGF-1R to activate the downstream PI3K/Akt signaling pathway. Mammalian target of rapamycin (mTOR) is a member of the PI3K-related kinase (PIKK family) and a downstream substrate of PI3K/Akt (34). Researchers have discovered that the hypothalamic IGF1/PI3K/Akt/mTOR signaling pathway regulates reproduction and development during puberty. By activating Kiss-1/GPR54, this pathway promotes GnRH release in the hypothalamus, thereby modulating adolescence (35).
Additionally, the estrogen receptor (ER) has been demonstrated to be strongly correlated with the pubertal development in children (36). ERα and ERβ are two isoforms encoded by ESR1 and ESR2, respectively, and are implicated in luteinization (37). A variety of cellular processes are regulated by SRC and HRAS, which are downstream of ESR1 in the estrogen pathway (38). MAPK1, also known as extracellular signal-regulated kinase 2 (ERK2), is a downstream effector of the epidermal growth factor receptor (EGFR) pathway. Activated ERK promotes follicle growth and ovulation via modulating the expression of gonadotropins LHβ and FSHβ (39). According to the KEGG pathway analysis, EGCG may affect estrogen signaling; ovarian steroidogenesis; insulin resistance; and other signaling pathways. These results supported the findings of our previous clinical study, which demonstrated that a 3-month intervention with 400 mg/d EGCG significantly reduced body fat percentage, ovarian volume and the number of follicles >4 mm in the ovaries of obese girls (14). This evidence suggested that EGCG might have considerable potential for preventing precocious puberty and improving obesity in girls. In addition, our prior experimental animal studies found that EGCG intervention remarkably decreased serum sex hormone levels in rats fed a high-fat diet (30). Moreover, evidence from in vivo studies suggests that the JAK/STAT pathway plays a crucial role in modulating GnRH neurons during puberty. Female GnRH neuron-specific JAK2 conditional knockout mice (JAK2 G-/-) shows reduced GnRH expression and neuronal activity. Furthermore, JAK2 G-/- female mice exhibits delayed puberty and reduced fertility (40). Therefore, EGCG may prevent obesity-related precocious puberty via the JAK2-STAT3 pathway. The present findings offered a novel foundation for the clinical application of EGCG and further related research.
Despite the findings of this study, the current study has certain limitations which should not be overlooked. Firstly, despite the advancements being made, there is still a lack of metabolomics data available for the precise identification of metabolites. Secondly, the sample size was too small. To improve this, it is essential to conduct future studies with a greater sample size of patients. Thirdly, due to the restrictions imposed by certain databases, it is not possible to retrieve all the active targets of EGCG, and the targets and pathways are interrelated and modulate each other. Further research is still required to gain a more comprehensive understanding of EGCG’s pharmacological effects and mechanisms in the prevention of obesity-related precocious puberty, which can be explored and confirmed through both in vivo and in vitro experiments. In the future, it will be necessary to thoroughly validate the details of omics data, including species markers that are distinct, predicted metabolic pathways, and potential functional interactions. Additionally, we can continue to explore the effects of EGCG on genes associated with estrogen, lipid metabolism, and energy metabolism, and further investigate the potential role of EGCG in regulating the endogenous metabolites of obesity-related precocious puberty. Furthermore, considering that the physiological association between EGCG and obesity-related precocious puberty is multifactorial, we will also use transcriptomic and proteomic technologies to reveal the correlation between EGCG and obesity-related precocious puberty at the gene and protein levels with the technical advantages of their high sensitivity and quantitative accuracy in the future research.
In summary, EGCG may serve a role in preventing obesity-related precocious puberty through targets such as AKT1, EGFR, ESR1, STAT3, IGF1, and MAPK1 that acted on multiple signaling pathways, including the estrogen, PI3K-Akt, MAPK, and Jak-STAT pathways. In this study, we investigated the mechanism by which EGCG prevented obesity-related precocious puberty through a network pharmacology approach and found that multiple targets and pathways were involved in the underlying mechanism. This study provided a theoretical foundation for subsequent research.
Data availability statementThe datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Ethics statementThe studies involving human participants were reviewed and approved by the Ethics Committee of Xinhua Hospital. Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin. The animal study was reviewed and approved by the Ethics Committee of Xinhua Hospital.
Author contributionsXS and LL conceived and designed the study. QG, LX, QD, YS, and JH collected data. QG drafted the manuscript. PW provided major comments to the manuscript. All authors contributed to the article and approved the submitted version.
FundingThis work is funded by the Science and Technology Research Project (Medical and Health Field) of Songjiang District of Shanghai (No. 22SJKGGG10), National Natural Science Foundation of China (No. 82173496) and the first National Nutrition Research Fund (No.CNS-NNSRG2019-39).
AcknowledgmentsThe authors gratefully acknowledge everyone who helped us in this study.
Conflict of interestThe authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s noteAll claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary materialThe Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fendo.2023.1159657/full#supplementary-material
Supplementary Table 1 | A total of 357 EGCG targets.
Supplementary Table 2 | A total of 153 common targets in "Precocious puberty", "Obesity" and "EGCG".
Supplementary Table 3 | Common targets in "Precocious puberty" and "Obesity".
Supplementary Data Sheet 1 | Endogenous differential metabolites between two groups.
AbbreviationsEGCG, (-)-Epigallocatechin-3-gallate; LC-ESI-MS/MS, liquid chromatography-electrospray ionization ion-trap tandem mass spectrometry; GnRHa, gonadotropin-releasing hormone analog; OPLS-DA, orthogonal partial least squares discriminant analysis; VIP, Variable importance in the projection; HMDB, Human Metabolome Database; SMILES, simplified molecular-input line-entry system; CTD, Comparative Toxicogenomics Database; KEGG, Kyoto Encyclopedia of Genes and Genomes; PPI, protein–protein interaction; 3D, three-dimensional; HPGA, hypothalamic-pituitary-ovarian axis; GnRH, gonadotropin-releasing hormone; IGF-1, insulin-like growth factor 1; mTOR, Mammalian target of rapamycin; ER, estrogen receptor; ERK2, extracellular signal-regulated kinase 2.
References2. Gu Q, Luo Y, Ye J, Shen X. comparative efficacy and safety of three current clinical treatments for girls with central precocious puberty: a network meta-analysis. Endocr Pract (2019) 25:717–28. doi: 10.4158/EP-2019-0008
PubMed Abstract | CrossRef Full Text | Google Scholar
3. Wu Y, Gu Q, Cui X, Zhu Z, Zang J, Wang Z, et al. Higher poultry consumption was associated with an earlier age at menarche. Acta Paediatr (2021) 110:889–95. doi: 10.1111/apa.15554
PubMed Abstract | CrossRef Full Text | Google Scholar
5. De Sanctis V, Soliman AT, Di Maio S, Soliman N, Elsedfy H. Long-term effects and significant adverse drug reactions (ADRs) associated with the use of gonadotropin-releasing hormone analogs (GnRHa) for central precocious puberty: a brief review of literature. Acta BioMed (2019) 90:345–59. doi: 10.23750/abm.v90i3.8736
PubMed Abstract | CrossRef Full Text | Google Scholar
7. Roberts SA, Kaiser UB. GENETICS IN ENDOCRINOLOGY: genetic etiologies of central precocious puberty and the role of imprinted genes. Eur J Endocrinol (2020) 183:R107–17. doi: 10.1530/EJE-20-0103
PubMed Abstract | CrossRef Full Text | Google Scholar
8. Heras V, Castellano JM, Fernandois D, Velasco I, Rodríguez-Vazquez E, Roa J, et al. Central ceramide signaling mediates obesity-induced precocious puberty. Cell Metab (2020) 32:951–66. doi: 10.1016/j.cmet.2020.10.001
PubMed Abstract | CrossRef Full Text | Google Scholar
9. Hong YH, Woo YJ, Lee JH, Shin Y-L, Lim H-S. Association between dietary habits and parental health with obesity among children with precocious puberty. Children (Basel) (2020) 7:220. doi: 10.3390/children7110220
PubMed Abstract | CrossRef Full Text | Google Scholar
12. Casanova E, Salvadó J, Crescenti A, Gibert-Ramos A. Epigallocatechin gallate modulates muscle homeostasis in type 2 diabetes and obesity by targeting energetic and redox pathways: a narrative review. Int J Mol Sci (2019) 20:532. doi: 10.3390/ijms20030532
PubMed Abstract | CrossRef Full Text | Google Scholar
13. Chen IJ, Liu C-Y, Chiu J-P, Hsu C-H. Therapeutic effect of high-dose green tea extract on weight reduction: a randomized, double-blind, placebo-controlled clinical trial. Clin Nutr (2016) 35:592–9. doi: 10.1016/j.clnu.2015.05.003
PubMed Abstract | CrossRef Full Text | Google Scholar
14. Xie L, Tang Q, Yao D, Gu Q, Zheng H, Wang X, et al. Effect of decaffeinated green tea polyphenols on body fat and precocious puberty in obese girls: a randomized controlled trial. Front Endocrinol (Lausanne) (2021) 12:736724. doi: 10.3389/fendo.2021.736724
PubMed Abstract | CrossRef Full Text | Google Scholar
15. Yang Q-J, Zhao J-R, Hao J, Li B, Huo Y, Han Y-L, et al. Serum and urine metabolomics study reveals a distinct diagnostic model for cancer cachexia. J Cachexia Sarcopenia Muscle (2018) 9:71–85. doi: 10.1002/jcsm.12246
PubMed Abstract | CrossRef Full Text | Google Scholar
16. Nogales C, Mamdouh ZM, List M, Kiel C, Casas AI, Schmidt HHHW. Network pharmacology: curing causal mechanisms instead of treating symptoms. Trends Pharmacol Sci (2022) 43:136–50. doi: 10.1016/j.tips.2021.11.004
PubMed Abstract | CrossRef Full Text | Google Scholar
17. Yue Z, Chen Y, Dong Q, Li D, Guo M, Zhang L, et al. Acrylamide induced glucose metabolism disorder in rats involves gut microbiota dysbiosis and changed bile acids metabolism. Food Res Int (2022) 157:111405. doi: 10.1016/j.foodres.2022.111405
PubMed Abstract | CrossRef Full Text | Google Scholar
19. Daina A, Michielin O, Zoete V. SwissTargetPrediction: updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res (2019) 47:W357–64. doi: 10.1093/nar/gkz382
PubMed Abstract | CrossRef Full Text | Google Scholar
20. Wishart DS, Feunang YD, Guo AC, Lo EJ, Marcu A, Grant JR, et al. DrugBank 5.0: a major update to the DrugBank database for 2018. Nucleic Acids Res (2018) 46:D1074–82. doi: 10.1093/nar/gkx1037
PubMed Abstract | CrossRef Full Text | Google Scholar
22. Stelzer G, Rosen N, Plaschkes I, Zimmerman S, Twik M, Fishilevich S, et al. The GeneCards suite: from gene data mining to disease genome sequence analyses. Curr Protoc Bioinf (2016) 54:1.30.1–33. doi: 10.1002/cpbi.5
CrossRef Full Text | Google Scholar
23. Piñero J, Ramírez-Anguita JM, Saüch-Pitarch J, Ronzano F, Centeno E, Sanz F, et al. The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res (2020) 48:D845–55. doi: 10.1093/nar/gkz1021
PubMed Abstract | CrossRef Full Text | Google Scholar
24. Davis AP, Grondin CJ, Johnson RJ, Sciaky D, Wiegers J, Wiegers TC, et al. Comparative toxicogenomics database (CTD): update 2021. Nucleic Acids Res (2021) 49:D1138–43. doi: 10.1093/nar/gkaa891
PubMed Abstract | CrossRef Full Text | Google Scholar
25. Szklarczyk D, Gable AL, Nastou KC, Lyon D, Kirsch R, Pyysalo S, et al. The STRING database in 2021: customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res (2021) 49:D605–12. doi: 10.1093/nar/gkaa1074
PubMed Abstract | CrossRef Full Text | Google Scholar
27. Burley SK, Berman HM, Bhikadiya C, Bi C, Chen L, Di Costanzo L, et al. RCSB protein data bank: biological macromolecular structures enabling research and education in fundamental biology, biomedicine, biotechnology and energy. Nucleic Acids Res (2019) 47:D464–74. doi: 10.1093/nar/gky1004
PubMed Abstract | CrossRef Full Text | Google Scholar
28. Latronico AC, Brito VN, Carel J-C. Causes, diagnosis, and treatment of central precocious puberty. Lancet Diabetes Endocrinol (2016) 4:265–74. doi: 10.1016/S2213-8587(15)00380-0
PubMed Abstract | CrossRef Full Text | Google Scholar
29. Klein DA, Emerick JE, Sylvester JE, Vogt KS. Disorders of puberty: an approach to diagnosis and management. Am Fam Physician (2017) 96:590–9.
PubMed Abstract | Google Scholar
30. Gu Q, Wang X, Xie L, Yao X, Qian L, Yu Z, et al. Green tea catechin EGCG could prevent obesity-related precocious puberty through NKB/NK3R signaling pathway. J Nutr Biochem (2022) 108:109085. doi: 10.1016/j.jnutbio.2022.109085
PubMed Abstract | CrossRef Full Text | Google Scholar
33. Liu P, Kong F, Wang J, Lu Q, Xu H, Qi T, et al. Involvement of IGF-1 and MEOX2 in PI3K/Akt1/2 and ERK1/2 pathways mediated proliferation and differentiation of perivascular adipocytes. Exp Cell Res (2015) 331:82–96. doi: 10.1016/j.yexcr.2014.09.011
PubMed Abstract | CrossRef Full Text | Google Scholar
34. Ingaramo P, Alarcón R, Muñoz-de-Toro M, Luque EH. Are glyphosate and glyphosate-based herbicides endocrine disruptors that alter female fertility? Mol Cell Endocrinol (2020) 518:110934. doi: 10.1016/j.mce.2020.110934
PubMed Abstract | CrossRef Full Text | Google Scholar
35. Shao P, Wang Y, Zhang M, Wen X, Zhang J, Xu Z, et al. The interference of DEHP in precocious puberty of females mediated by the hypothalamic IGF-1/PI3K/Akt/mTOR signaling pathway. Ecotoxicol Environ Saf (2019) 181:362–9. doi: 10.1016/j.ecoenv.2019.06.017
PubMed Abstract | CrossRef Full Text | Google Scholar
36. Luo Y, Liu Q, Lei X, Wen Y, Yang Y-L, Zhang R, et al. Association of estrogen receptor gene polymorphisms with human precocious puberty: a systematic review and meta-analysis. Gynecol Endocrinol (2015) 31:516–21. doi: 10.3109/09513590.2015.1031102
PubMed Abstract | CrossRef Full Text | Google Scholar
37. Feldman RD, Limbird LE. GPER (GPR30): a nongenomic receptor (GPCR) for steroid hormones with implications for cardiovascular disease and cancer. Annu Rev Pharmacol Toxicol (2017) 57:567–84. doi: 10.1146/annurev-pharmtox-010716-104651
PubMed Abstract | CrossRef Full Text | Google Scholar
38. Ożegowska K, Brązert M, Ciesiółka S, Nawrocki MJ, Kranc W, Celichowski P, et al. Kempisty, genes involved in the processes of cell proliferation, migration, adhesion, and tissue development as new potential markers of porcine granulosa cellular processes : a microarray approach. DNA Cell Biol (2019) 38:549–60. doi: 10.1089/dna.2018.4467
PubMed Abstract | CrossRef Full Text | Google Scholar
39. Yang L, Xu H, Chen Y, Miao C, Zhao Y, Xing Y, et al. Melatonin: multi-target mechanism against diminished ovarian reserve based on network pharmacology. Front Endocrinol (Lausanne) (2021) 12:630504. doi: 10.3389/fendo.2021.630504
PubMed Abstract | CrossRef Full Text | Google Scholar
40. Wu S, Divall S, Hoffman GE, Le WW, Wagner K-U, Wolfe A. Jak2 is necessary for neuroendocrine control of female reproduction. J Neurosci (2011) 31:184–92. doi: 10.1523/JNEUROSCI.2974-10.2011
留言 (0)