
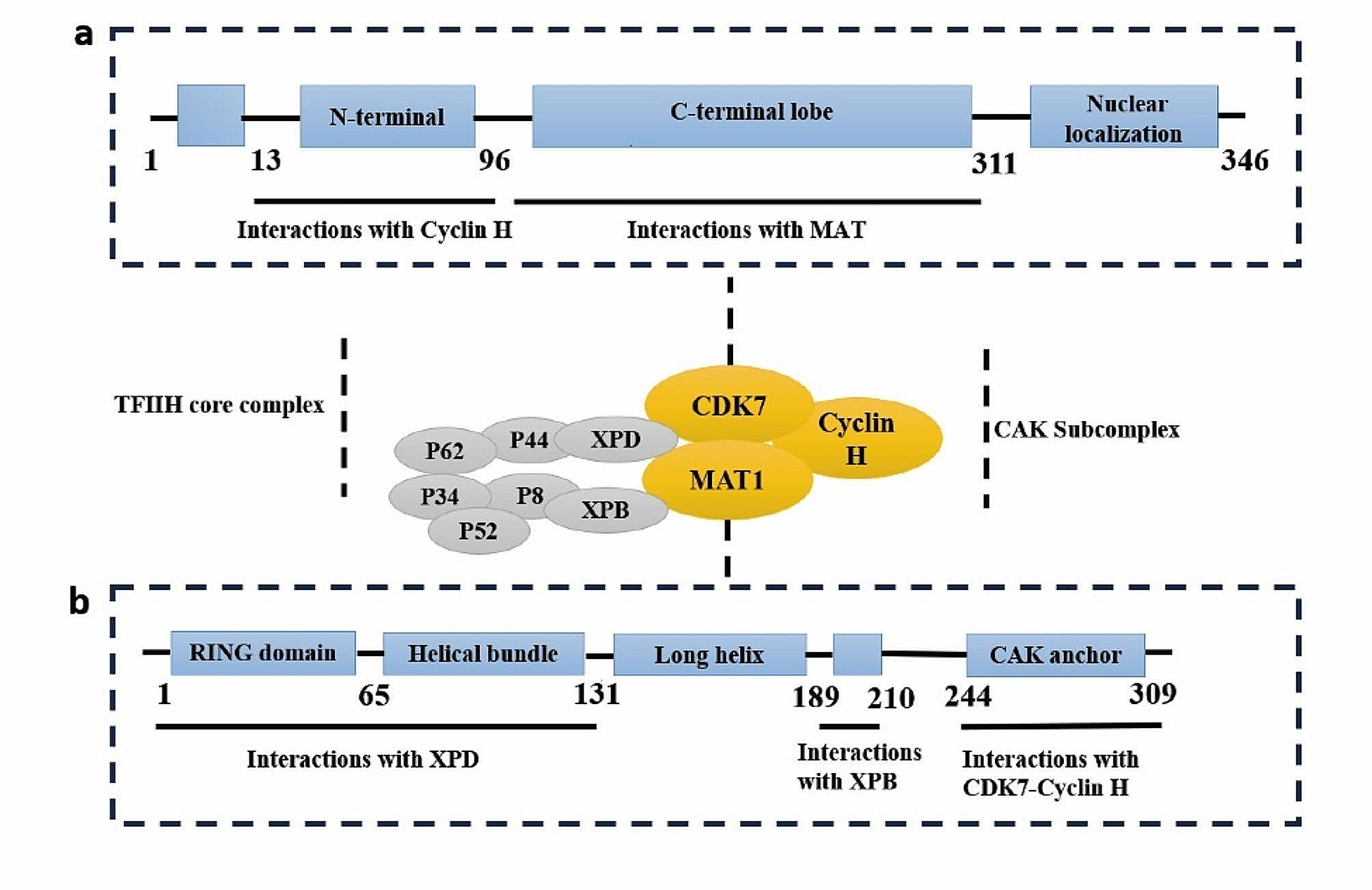
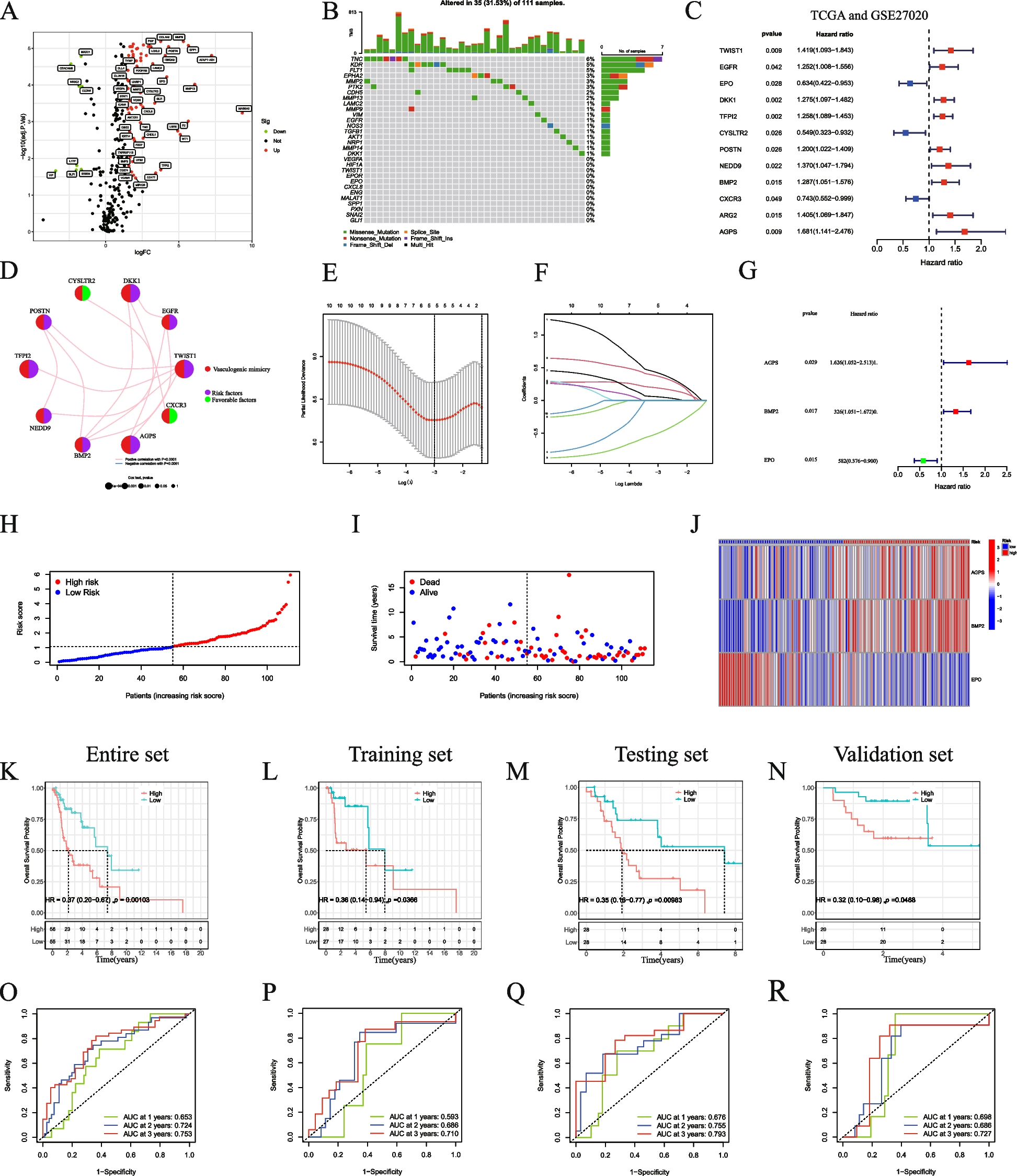
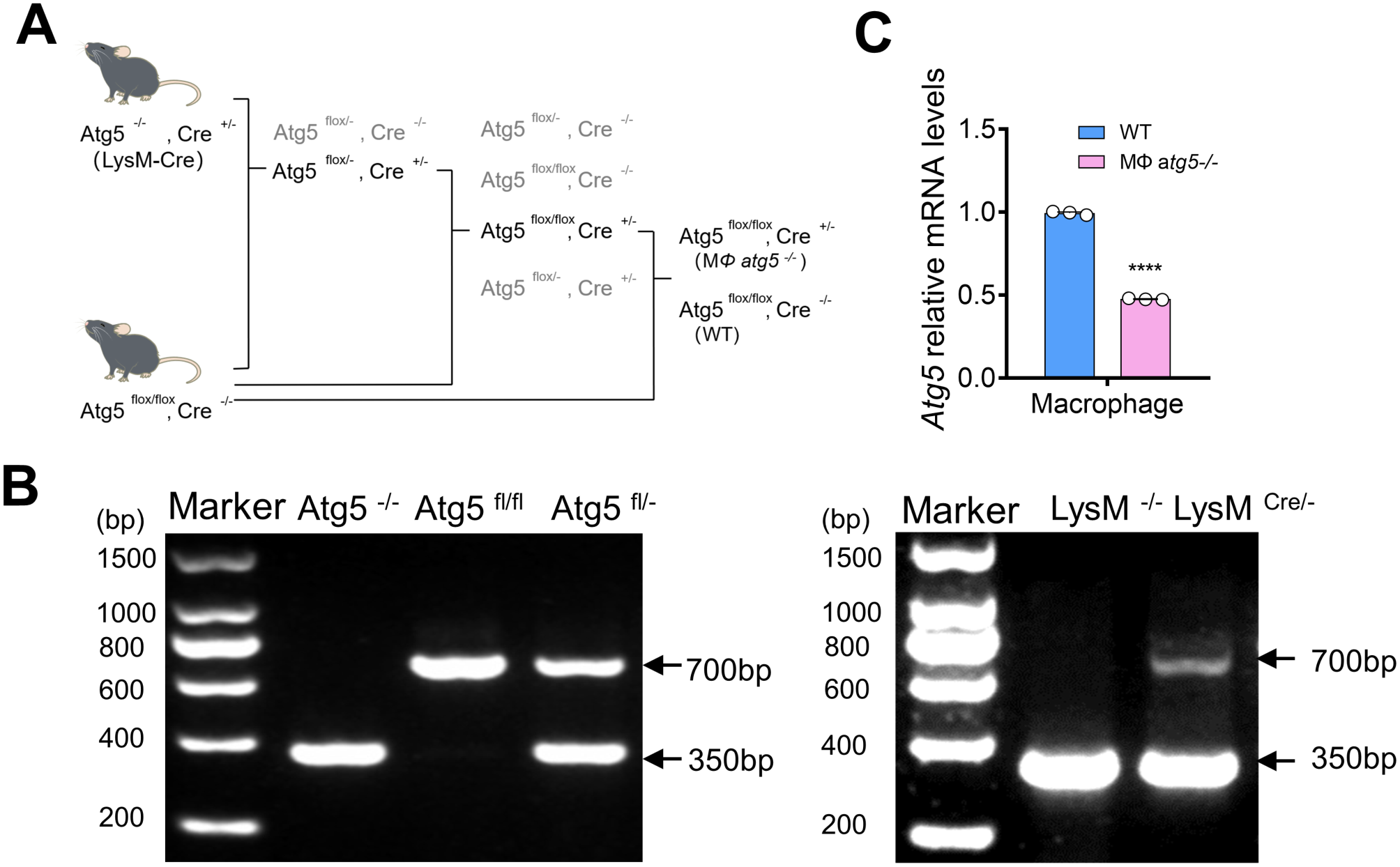
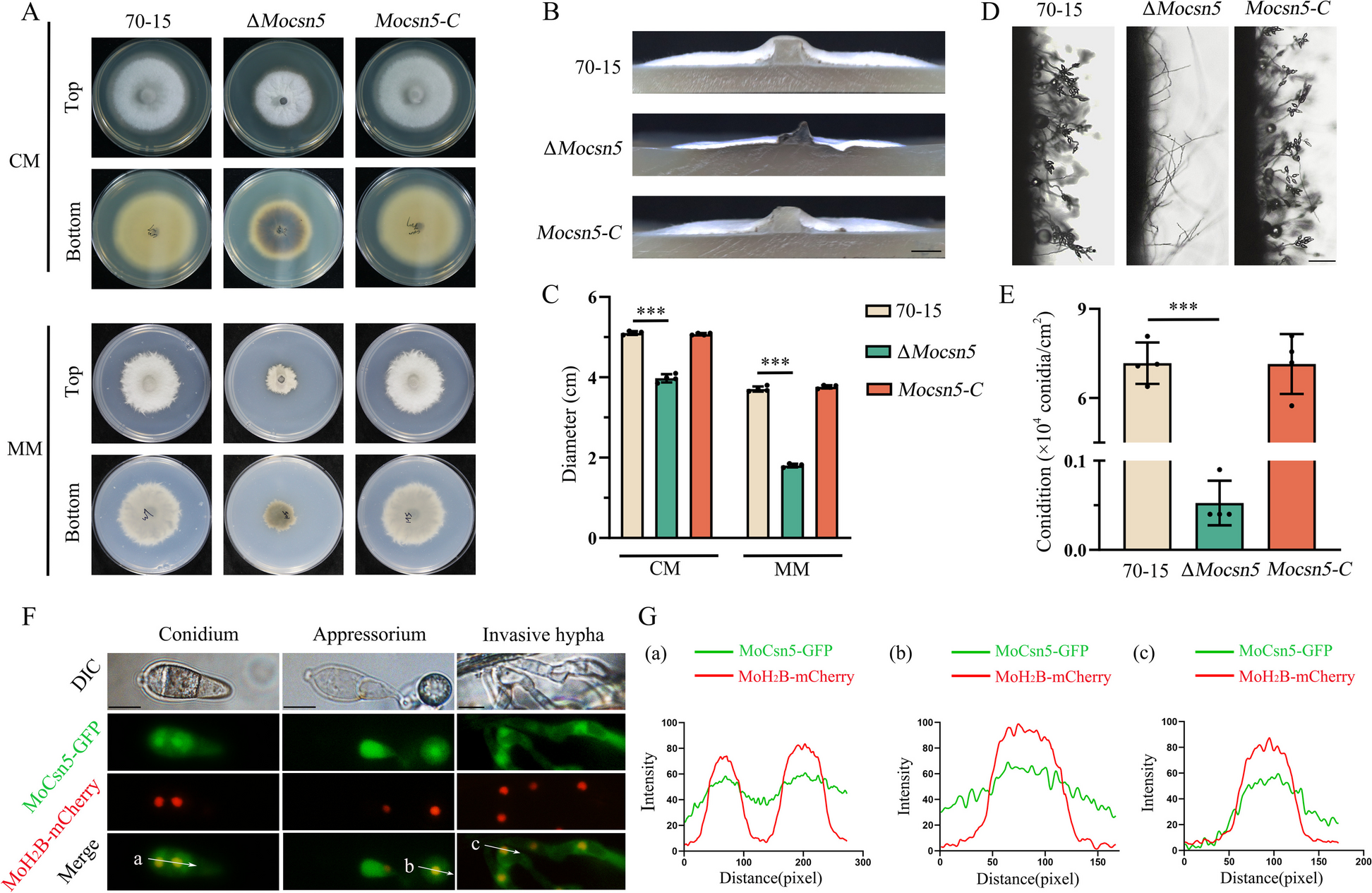
記住我
To study the interplay between secreted FGFs and galectins we produced representative members from all five FGF subgroups that are secreted in a conventional manner and contain N-glycosylation motifs as fusions with Fc in CHO cells: FGF4-Fc, FGF9-Fc, FGF10-Fc, FGF18-Fc and FGF23-Fc (Fig. 1B, lanes 1, 3, 5, 7 and 9). These FGFs contain either one (FGF4 and FGF9) or two (FGF10 and FGF18) putative N-glycosylation sites, which are located within the N-terminal extensions or in the FGF β-trefoil domain, in an area that based on the structures of the FGF/FGFR complexes is likely to be distant from the FGFR binding sites (Fig. 1A). Unlike the aforementioned FGFs, FGF23 lacks an N-glycosylation motif, but is O-glycosylated in the linker region between the FGFR and α-Klotho binding sites, and this modification regulates FGF23 activity (Fig. 1A) [14]. Since the Fc fragment of IgG1 can be N-glycosylated, we used FGF23-Fc as a control and demonstrated that after PNGase F treatment there is only a very slight change in the migration of FGF23-Fc in SDS-PAGE (which reflects de-glycosylation of the Fc) (Fig. 1B, lane 9 and 10). In contrast, PNGase F treatment largely altered the migration of FGF4-Fc, FGF9-Fc, FGF10-Fc and FGF18-Fc, confirming the N-glycosylation of these proteins (Fig. 1B, lanes 1–8).
The human galectin family consists of twelve proteins that differ in their oligomeric state and carbohydrate specificity (Fig. 1C). To screen for galectins that bind FGFs, we developed galectin dot blot arrays with recombinant human galectins [34]. FGFs-Fc, Fc (positive controls), and recombinant galectins were immobilized on PVDF membrane, membranes were blocked and the galectin arrays were incubated with equimolar concentrations of FGFs-Fc or Fc as a negative control. After extensive washing, complexes between individual galectin and FGFs were detected with anti-Fc antibody conjugated to HRP and chemiluminescence. The spots with positive controls: FGFs-Fc and Fc were readily detected with anti-Fc antibody, which confirms the validity of our experimental approach (Fig. 1D, rows 14 and 15). We observed positive signals for the vast majority of distinct galectins and FGF4-Fc, FGF9-Fc, FGF10-FC and FGF18-Fc (Fig. 1D). In contrast, for O-glycosylated FGF23-Fc we detected significantly fewer interactions with galectins, with positive signals identified mainly for galectin-3 and − 9 (Fig. 1D, rows 3 and 7). The presence of lactose in dot blot experiments with galectins greatly reduced the signal detected for most of galectins, while for few of galectins (mainly galectin-4, -13, -14 and − 16) the signal was enhanced, indicating their possible sugar-independent interaction with FGF4-Fc (Fig. S1). These data indicate that different galectins can directly interact with glycosylated FGFs.
To confirm these findings, we used biolayer interferometry (BLI). FGFs-Fc and the Fc fragment were immobilized on Protein-A BLI biosensors in a pairwise manner (FGF-Fc and Fc as a control), incubated with recombinant galectins and the association and dissociation stages of the interaction between the proteins tested were monitored. For FGF4-Fc, FGF9-Fc, FGF10-Fc and FGF18-Fc, the strongest signals were obtained for galectin-1, -3, -7 and − 8 (Fig. 1E). For O-glycosylated FGF23-Fc, we detected virtually no interaction with galectin-1, -3 and − 7 and a weak signal for galectin-8 and − 13 (Fig. S2). The presence of lactose completely inhibited the binding of galectin-1, -3, -7 and − 8 to FGF4-Fc (Fig. S3). We also confirmed with BLI that the interaction with GST-tagged FGF4-Fc was not due to non-specific binding to GST (Fig. S4).
Galectin-8 is a tandem repeat galectin composed of two CRDs with distinct carbohydrate specificities (Fig. 1C) [35]. We produced separate CRDs of galectin-8 and demonstrated that only gal-8CRD1 interacts with FGF4-Fc, indicating that CRD1 is exclusively involved in FGF4-Fc binding (Fig. 1F).
These data indicate that galectins from all three families directly interact with all tested N-glycosylated FGFs, while they display very limited binding to O-glycosylated FGF23. Since galectin-1, -3, -7 and − 8 showed positive signals in both galectin dot-blots and BLI, we decided to focus on these galectins in subsequent studies.
N-glycans of FGFs constitute binding sites for galectinsWe evaluated whether the direct interaction between galectin-1, -3, -7 and − 8 and FGFs occurs via N-glycans of FGFs. BLI experiments with enzymatically de-glycosylated FGFs-Fc revealed that FGF4, FGF10 and FGF18 strictly require N-glycosylation for binding of galectins (Fig. 2A, C and D). However, we observed a weaker dependence on N-glycosylation for the interaction between FGF9-Fc and galectins (Fig. 2B). These data indicate that N-glycans of FGFs are recognized by galectin-1, -3, -7 and − 8.
Fig. 2
N-glycans of FGFs are recognized by galectins. A-D BLI analyses of the interaction between N-glycosylated and de-glycosylated FGFs and galectins. N-glycosylated and enzymatically de-glycosylated FGFs: FGF4-Fc (A), FGF9-Fc (B), FGF10-Fc (C) and FGF18-Fc (D) were immobilized on Protein-A biosensors (blue lines) in a pairwise fashion with equimolar concentrations of PNGase F-treated FGF4-Fc, FGF9-Fc, FGF10-Fc and FGF18-Fc, respectively (red lines), and the association and dissociation phases were recorded with BLI. Representative results from at least three independent experiments are shown
Subsequently, for reasons of ease of isolation and handling, we decided to focus on FGF4-Fc as a representative N-glycosylated FGF and studied in depth its functional interconnection with identified galectins. We measured kinetic parameters of the interaction between FGF4-Fc and galectins. As shown in Table 1 galectins bind FGF4 with affinity in high nanomolar range (KD of about 10− 7 M).
Table 1 Kinetic parameters of the interaction between the studied galectins and FGF4 Multivalency of galectins is strictly required for differential fine-tuning of the trafficking and signaling of N-glycosylated FGF4Since we identified a direct interaction of galectin-1, -3, -7 and − 8 with the N-linked sugars of FGFs, we wondered whether galectins could modulate the trafficking, signaling and function of N-glycosylated FGF4. To investigate whether endogenous galectins adjust the signaling of FGF4-Fc, we washed serum-starved NIH3T3 cells with lactose and treated the cells with FGF4-Fc. Lactose washes significantly reduced FGF4-Fc ability to induce phosphorylation of FGFR1 and ERK1/2, indicating that endogenous extracellular galectins modulate FGF4 (Fig. 3A). We assessed the effect of particular identified galectins on the ability of FGF4-Fc to activate FGFR-dependent signaling by pre-forming FGF4-Fc complexes with the galectins tested, incubating the complexes with cells and evaluating pFGFR and pERK1/2 levels. Interestingly, galectin-1 and − 3 significantly inhibited the activation of FGFR1 and ERK1/2 by FGF4-Fc, while galectin-7 and − 8 increased receptor activation and upregulated downstream signaling cascades (Fig. 3B).
Fig. 3
Multivalency of galectins is crucial for the attraction of glycosylated FGF4-Fc to the cell surface and for the modulation of FGF4-Fc trafficking and signaling. A Effect of endogenous galectins on FGF4-Fc signaling. Serum-starved NIH3T3 cells were washed with 50 mM lactose prior cell supplementation with various concentrations of FGF4-Fc (2–20 ng/mL). Cells were lysed and analyzed with WB using the indicated antibodies. CBB served as a loading control (left panel). Densitometric analyses of the effect of lactose washes on the activation of FGF-Fc-dependent signaling pathways by 20 ng/mL FGF4-Fc (right panel). Mean values from at least three independent experiments +/- SEM are shown. Statistical analyses were performed with Student’s t-test (*p < 0.05; **p < 0.005 and ***p < 0.001). B and C Effects of the wild type galectins (B) and their monovalent variants (C) on FGF4-Fc signaling. Serum-starved NIH3T3 cells were treated with FGF1 (100 ng/mL, control), different concentrations of FGF4-Fc (2 and 20 ng/mL) in the presence or absence of recombinant galectins (1 µg/mL) or their monovalent variants (5 µg/mL). Cells were lysed and analyzed with WB using the indicated antibodies (left panel). CBB and tubulin served as loading controls. Densitometric analyses of the effect of galectins on FGF4-Fc (20 ng/mL) signaling (right panel). Mean values from at least three independent experiments +/- SEM are shown. Statistical analyses were performed with Student’s t-test (*p < 0.05; **p < 0.005 and ***p < 0.001). The schemes of the wild type galectins and their engineered variants with altered valency are shown (left panel). D Effects of the wild type galectins and their engineered variants with altered valency on the cell binding and endocytosis of FGF4-Fc. U2OS-R1 cells were incubated for 30 min with FGF4-Fc (20 µg/mL) either at 4 °C (for cell binding analysis) or at 37 °C (for growth factor endocytosis analysis) in the presence of the studied galectins and their variants (20 µg/mL). Cells were either fixed (4 °C samples) or fixed and permeabilized (37 °C samples), nuclei were labelled with NucBlue and FGF4-Fc was detected with Zenon-AF-488 using fluorescence microscopy. Representative images from at least three independent experiments are shown. Scale bars represent 20 μm. To quantify cell binding by FGF4-Fc in the presence of galectin variants, the total fluorescence of at least 20 cells from three fields of view/condition was measured in three independent experiments using Zeiss ZEN 2.3 software. Statistical analyses were performed with Student’s t-test (*p < 0.05; **p < 0.005 and ***p < 0.001). The schemes of the wild type galectins and their engineered variants with altered valency are shown (left panel)
To evaluate whether multivalency of galectins is required for modulation of FGF4-Fc signaling we produced truncated variants of galectin-1, -3 including solely CRD domains and lacking regions responsible for oligomerization [18, 36, 37]. CRD domains of galectin-1 and − 3, in contrast to the wild type proteins, had no effect on FGF4-Fc signaling (Fig. 3C). Similarly, uncoupled CRD1 and CRD2 domains of galectin-8 displayed no FGF4-Fc stimulatory activity (Fig. 3C).
Using fluorescence microscopy, we evaluated the effect of wild type galectins and their monovalent variants on the efficiency of FGF4-Fc binding and internalization by cells. We observed increased FGF4-Fc interaction with cells in the presence of all tested galectins (Fig. 3D). The strongest increase in the signal of FGF4-Fc at the cell surface was found when the growth factor was co-incubated with galectin-8 (Fig. 3D). Importantly, the increased binding of FGF4-Fc to cells strictly required the multivalency of galectins, as their monomeric engineered variants: gal-1CRD, gal-3CRD, gal-8CRD1 and gal-8CRD2 were unable to enhance FGF4-Fc interaction with cells (Fig. 3D). To further prove that multivalency of galectins is required for their action on FGF4-Fc, we engineered pentavalent galectin-1, gal-1CRD-CC.5x, by fusing gal-1CRD with coiled coil motif enabling pentamerization [27]. We detected the massive accumulation of FGF4-Fc at the cell surface after co-incubation with multivalent gal-1CRD-CC.5x (Fig. 3D).
When FGF4-Fc was incubated with cells at 37 °C, the growth factor was effectively endocytosed and no cell surface-bound FGF4-Fc signal was detected, which we confirmed by co-localizing the intracellular FGF4-Fc signal with an early endosome marker protein, EEA1 (Fig. S5). FGF4-Fc internalization was differentially adjusted by the galectins tested, where galectin-1 and − 3 enhanced the efficiency of FGF4-Fc internalization, while galectin-7 had no significant effect on cellular uptake of FGF4-Fc (Fig. 3D). In the case of galectin-8, in addition to numerous spots indicating endosomes with internalized FGF4-Fc, we observed a strong signal of FGF4-Fc remaining on the cell surface (Fig. 3D). Internalization of FGF4-Fc was not affected by the monovalent engineered galectins: gal-8CRD1, gal-8CRD2, gal-1CRD and gal-3CRD, while gal-1CRD-CC.5x, like the wild type galectin-8, caused, additionally to intracellular FGF4-Fc spots, partial accumulation of FGF4-Fc on the cell surface (Fig. 3D).
We next determined the cellular consequences of galectin-induced changes in FGF4 signaling. We demonstrated that FGF4-Fc signaling results in highly efficient stimulation of NIH3T3 cell division (Fig. 4A). The combination of the studied galectins with FGF4-Fc significantly enhanced the mitogenic activity of the cells, and this effect was completely abolished by the presence of lactose (Fig. 4A, Fig. S6). We also observed that galectins alone could also inhibit apoptosis in a dose-dependent manner (Fig. S7) and improve viability of cells even at high concentration (Fig. S8). Galectins have been previously implicated in regulation of cellular glucose uptake [38]. Although FGF4-Fc and galectins alone stimulated glucose uptake by 3T3 adipocytes, FGF4-Fc/galectin mixtures displayed the same metabolic activity as FGF4-Fc alone (Fig. 4B).
Fig. 4
Galectins modulate cellular activities of FGF4. A The effect of FGF4-Fc (5 ng/mL), galectins (5 µg/mL) and a mixture of FGF4-Fc (5 ng/mL)/ galectin (5 µg/mL) on NIH3T3 cell proliferation assessed with the Presto Blue Cell Viability Reagent. Mean values +/-SD from at least three independent experiments are shown. Statistical analyses were performed with Student’s t-test (*p < 0.05; **p < 0.005 and ***p < 0.001). B Effects of FGF4-Fc (108 ng/mL), galectins (50 µg/mL) and a mixture of FGF4-Fc (108 ng/mL) / galectin (50 µg/mL) on glucose uptake by adipocytes. Mean values +/-SD from at least three independent experiments are shown. Statistical analyses were performed with Student’s t-test (*p < 0.05; **p < 0.005 and ***p < 0.001). C A hypothetical model of the modulation of FGF4 signaling by galectins. Simultaneous binding of multivalent galectins to the N-glycans of FGF4 and to yet unidentified extracellular matrix (ECM) components attracts FGF4 to the cell surface, thereby forming a reservoir of growth factor in the vicinity of plasma membrane-embedded FGFRs. Galectin-facilitated cell binding is reflected in altered FGF4/FGFR signaling and endocytosis, which ultimately shapes cell division without affecting metabolic activity of cells. The diversity of galectins’ effects on FGF4 is likely determined by different binding partners of particular galectins in the ECM
All these data indicate that galectins, through binding to FGF4 N-glycans, modulate cell binding, signaling and growth factor internalization, and adjust FGF4-dependent cellular processes. Moreover, our data suggest that the multivalency of galectins is critical for their activity towards N-glycosylated FGF4.
留言 (0)