
記住我


Cannabis sativa derivatives have become the most widely used illicit drugs worldwide, and regular cannabis use mainly begins during adolescence. Indeed, it is estimated that 15.5% of EU inhabitants aged 15–34 used cannabis in 2021, while 19.1% of those aged 15–24 had consumed cannabis in the last year and 10.4% in the last month (1). With a recent increase in the prevalence of cannabis use disorder (CUD) and a reduction in the perceived risk of cannabis use (2), it is necessary to further understand the neurobiological consequences associated with this cannabis consumption. However, the consequences of early-life cannabis use are still poorly understood. Recent evidence indicates that early exposure to Δ9-tetrahydrocannabinol (THC), the primary psychotropic constituent of cannabis, produces changes in the structure and function of brain circuits implicated in decision-making and cognitive processes (3). However, whether such changes might be long-lasting and persistently disrupt healthy behaviors remains unknown.
The adolescent period is a critical phase of brain development, neuronal maturation, and restructuring (4–6). Imaging studies have reported that higher-order structures, such as the prefrontal cortex, are still immature at that time point (7, 8) and they cannot fully inhibit behavior favoring impulsivity and risk-taking (9). Additionally, the striatal and limbic circuits are hyperactive during adolescence, leading to greater emotional reactivity and reward-seeking behaviors (7). These studies propose that early-life cannabis exposure may impair neurodevelopment and induce changes that affect the adult brain (8). Thus, a loss of gray matter was revealed in the prefrontal areas of adolescent cannabis users (10). Moreover, THC mediates its pharmacological effect through the activation of the endocannabinoid (eCB) system, a key modulatory system crucial in synapse pruning, formation, and maturation (10, 11). Therefore, dysregulation of the eCB system by the action of THC or other exogenous cannabinoids during adolescence could disrupt normal brain development and function. These alterations have been demonstrated to be involved in the earlier onset of psychiatric disorders (6, 10, 12). In addition, a gateway hypothesis has been postulated, proposing a causal chain sequence in which cannabis consumption would be used prior to other illicit drugs and cannabis use would increase the likelihood of using other illicit drugs (9, 13, 14). However, studies on the long-term neurobiological consequences of cannabis consumption in juveniles are scarce and often contradictory.
Cannabis addiction can be conceptualized as a three-stage recurring cycle of “binge/intoxication,” “withdrawal/negative affect,” and “preoccupation/anticipation,” each sustained by neurobiological alterations mainly in the basal ganglia, extended amygdala, and prefrontal cortex, respectively (15). According to the Diagnostic and Statistical Manual of Mental Disorders (DSM-5), these stages are characterized by (1) dysregulation of rewarding or pleasurable effects, (2) emergence of a negative emotional state when access to the reward is prevented, and (3) loss of control in limiting drug intake despite repeated unsuccessful efforts to resist, respectively (8, 15). These stages are repeated and worsen over time, perpetuating the addiction cycle until they produce a maladaptive habit formation that is governed by compulsive behavior. Moreover, craving was included in the DSM-5 addiction diagnostic criteria as it is directly related to the vulnerability to relapse after abstinence (16, 17). In our mouse model of cannabis addiction, we study these three stages through the three hallmarks of addiction based on the DSM-5 addiction criteria: (1) persistence of drug-seeking, (2) motivation for the drug, and (3) compulsive-like behavior. Moreover, we also evaluate two parameters related to craving, resistance to extinction and drug-seeking behavior after abstinence, and two phenotypic traits considered to be factors of vulnerability to substance misuse, impulsivity and sensitivity to reward. According to its multifactorial origin, only a subset of individuals repeatedly exposed to the drug will develop addiction (9, 18, 19).
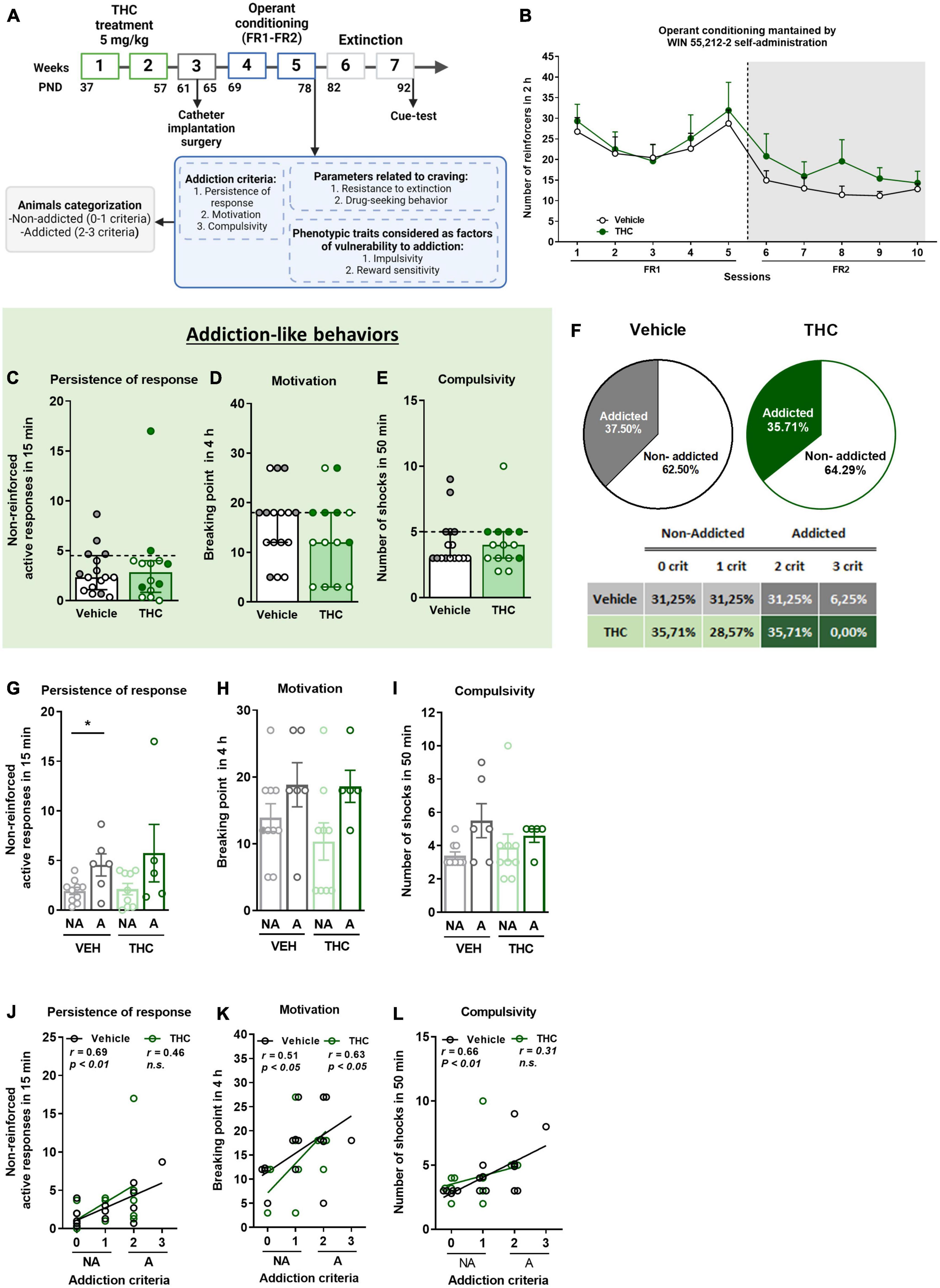
In this study, we aim to investigate the development of cannabis addiction-like behavior and related behavioral alterations in adult mice after adolescent exposure to THC. For this purpose, adolescent mice received daily administration of THC or vehicle from postnatal days (PND) 37 to 57, considered to be the adolescent period in mice, equivalent to 12–19 years old in humans (20). Then, mice were trained to acquire an operant intravenous (iv) self-administration conditioning sustained by WIN 55,212-2, a potent synthetic cannabinoid agonist (21). The addiction-like criteria were evaluated, as well as parameters related to craving and phenotypic traits, as previously described. We also evaluated molecular markers of cannabis addiction-like behavior in brain areas of the mesocorticolimbic circuit.
2. Materials and methods 2.1. AnimalsEight weeks old male C57BL/6J mice (n = 40) (Charles River, France) were housed individually in controlled laboratory conditions (21 ± 1°C, 55 ± 10%) with food and water available ad libitum. The male sex was chosen considering previous literature that has validated the operant WIN 55,212-2 self-administration model only in males (22). Mice were tested during the first hours of the dark phase of a reversed light/dark cycle (lights off at 8:00 a.m. and on at 20:00 p.m.). Body weight and food intake were monitored throughout the experiment. All animal procedures were approved by the local ethical committee (Comitè Ètic d’Experimentació Animal-Parc de Recerca Biomèdica de Barcelona, CEEA-PRBB, agreement N°9687) and conducted strictly in accordance with the guidelines of the European Communities Council Directive (2010/63/EU) regulating animal experimentation, in the animal facility at Universitat Pompeu Fabra-Barcelona Biomedical Research Park (UPF-PRBB; Barcelona, Spain). All the experiments were performed under blind and randomized conditions.
2.2. DrugsDronabinol, Δ-9-tetrahydrocannabinol (THC), generously gifted by Rhodes Pharmaceuticals, Coventry, RI, USA, stored at 100 mg/ml in sesame oil, was dissolved in a vehicle containing 5% ethanol 100%, 5% cremophor-LE (C5135, Sigma-Aldrich, USA) and 90% physiological saline solution, and administered by intraperitoneal (ip) injection at a dose of 5 mg/kg of body weight. For the self-administration, WIN 55, 212-2 [(R)-(+)-WIN 55,212-2 mesylate salt, Sigma-Aldrich, USA] was dissolved in one drop of Tween 80 (TWEEN 80, Sigma-Aldrich, USA) and then diluted in heparinized (1%) sterile saline solution and made available at 0.1 mg/kg for ip injection 24 h before the first operant session and 12.5 μg/kg/infusion for the self-administered iv infusions. The preparation was covered from the light and stored at room temperature. After each self-administration session, 0.05 ml of sodic heparin (Hospira 5%, Hospira, Pfizer) was applied through the iv catheter to avoid coagulation and obstruction of the latter. Thiopental sodium (5 mg/ml, Braun Medical S.A.) was dissolved in distilled water and delivered in a volume of 0.05 ml through the iv catheter.
2.3. Operant self-administration apparatusExperiments were performed in mouse operant chambers (model ENV-307A-CT, Med Associates Inc., Georgia, VT, USA) equipped with two nose-poke holes, one randomly selected as the active hole and the other as the inactive hole. A house light was located on the ceiling of the chamber, and two stimuli lights (cues) were located one inside the active hole and the other above it. Nose-poking in the active hole resulted in the delivery of one WIN 55,212-2 infusion (under the associated schedule) paired with the activation of the stimulus light located above the active hole, while nose-poking in the inactive hole had no consequences. The chambers were made of aluminum and acrylic and were housed inside sound- and light-attenuated boxes equipped with fans that provided ventilation and white noise. The chamber’s floor was a grid made of metal bars that could conduct electrical current when performing the shock test. WIN 55,212-2 was delivered in a volume of 23.5 μl over 2 s via a syringe mounted on a microinfusion pump (PHM-100A, Med-Associates, Georgia, VT, USA) and connected with flexible polymer tubing (0.96 mm outer diameter, Portex Fine Bore Polythene Tubing, Portex Limited, Kent, United Kingdom) to a single channel liquid swivel (375/25, Instech Laboratories, Plymouth Meeting, PA, USA) and to the mouse iv catheter.
2.4. Experimental designAdolescent mice received a daily dose of 5 mg/kg of THC or vehicle from PND 37 to PND 57. Afterward, mice were implanted with an intrajugular catheter in order to perform iv drug administrations. Subsequently, mice were trained to acquire an operant drug self-administration conditioning maintained by iv infusions of WIN 55,212-2 under a fixed-ratio (FR) 1 schedule of reinforcement during five sessions, followed by five sessions of a FR2 schedule of reinforcement. After the training, three addiction-like criteria resembling DSM-5 criteria for addiction, persistence to response, motivation, and compulsive-like behavior, two parameters related to craving, resistance to extinction and drug-seeking behavior, and two phenotypic traits considered factors of vulnerability to substance misuse, impulsivity and sensitivity to reward, were evaluated in each mouse (Supplementary Figure 3).
2.5. Adolescent THC treatmentWe evaluated the long-term effect of THC exposure during adolescence on WIN 55,212-2 self-administration, reinstatement, addiction-like criteria, and phenotypic traits present in adulthood. Mice were divided into two groups and administered daily with THC (n = 20) or its vehicle (n = 20) at a dose of 5 mg/kg of body weight from PND 37 to PND 57, corresponding to the adolescent period in mice based on previous literature (23). This dose of THC was chosen according to previous studies using similar doses to translate in rodents the doses used for smoked cannabis in humans (24–27). Body weight and food intake were strictly monitored for the entire period of treatment. Subsequently, animals underwent surgical procedures, and once adulthood was reached (PND 68), self-administration experiments were started.
2.6. WIN 55,212-2 self-administration 2.6.1. Jugular vein catheterizationMice were anesthetized by ip injection (0.2 ml/10 g of body weight) of ketamine hydrochloride (75 mg/kg of body weight, Ketamidor, Richter Pharma AG, Austria) and medetomidine hydrochloride (1 mg/kg of body weight, Domtor, Esteve, Spain) dissolved in 0.9% physiological saline and then implanted with indwelling iv catheters in the right jugular vein, as previously described (22). Briefly, a 6 cm long silicone tubing (0.3 mm inner diameter, 0.6 mm outer diameter; Silastic, Dow Corning, Houdeng-Goegnies, Belgium) was adapted to a 22-gauge steel cannula (Semat, Herts, United Kingdom) angled at a right angle and then embedded in a dental cement disk (Dentalon Plus, Heraeus Kulzer, Germany) with an underlying nylon mesh. The catheter tubing was inserted 1.1 cm into the right jugular vein and attached with a suture, and the remaining tubing was placed subcutaneous (sc) to the cannula, exiting at the midscapular region. All incisions were sutured and coated with a local analgesic (Blastoestimulina, Almirall, Spain). After, a post-surgery procedure consisting of an ip injection of antibiotic (1 mg/kg of body weight, Gentamicine, Genta-Gobens, Laboratorios Normon, Spain), a sc injection of analgesic (mixture of glucose serum (GlucosaVet, B. Braun Vet Care, Spain) and meloxicam (2 mg/kg of body weight, Metacam, Boehringer Ingelheim, Rhein) and a sc injection of anesthesia reverser, atipamezole hydrochloride (2.5 mg/kg of body weight, Revertor, Virbac, Spain) was applied, all dissolved in sterile 0.9% physiological saline. Mice were allowed to recover for 3 days with follow-up analgesics prior to the initiation of the self-administration sessions. The patency of iv catheters was evaluated by a thiopental sodium test at the end of the self-administration experimental sequence. If prominent signs of anesthesia were not observed immediately after injection, the mouse was removed from the experiment.
2.6.2. WIN 55,212-2 self-administration trainingThe operant model was applied similarly to previous drug self-administration paradigms (22, 28) with the inclusion of the addiction-like criteria, parameters related to craving and phenotypic traits. To avoid the aversive effects to the drug’s first administration, mice received an ip injection of WIN 55,212-2 24 h before the first self-administration session. Subsequently, mice were trained to acquire operant self-administration maintained by iv infusions of WIN 55,212-2. The schedule was a fixed ratio (FR) 1 schedule of reinforcement (one active nose-poking resulted in one drug delivery) during five consecutive sessions, followed by a progression to FR2 (two active nose-pokings resulted in one drug delivery) for another five sessions. All sessions were performed at the same time every day. Each daily self-administration session initiated with a priming injection of the drug, followed by two 55 min active periods separated by a 15 min drug-free period. The beginning of each operant session was signaled by turning on the house-light only during the first 3 s. During the active periods, each drug infusion was paired contingently with the cue light located above the active nose-poke under the specific FR schedule. These cue lights, together with the noise of the infusion pump, acted as environmental cues signaling the drug infusion. A 10 s time-out period was fixed after each drug delivery, during which the cue light was off and no infusion was provided after responding to the active nose-poke. Responses to the active and inactive holes and all responses executed during the time-out period were recorded. During the drug-free period, neither reinforcer nor contingent cue light was delivered, and this period was signaled by the illumination of the whole self-administration chamber. The session was concluded after 50 reinforcers were delivered or after 125 min, whichever occurred first. If a mouse reached 50 reinforcers, the limit was expanded to 100 reinforcers in the following session. The acquisition of the self-administration behavior was achieved when all of the following conditions were met: (1) mice maintained 80% stability in three consecutive sessions, that is, the variance across these 3 days was 20% or less, (2) at least 75% responding in the active hole, and (3) a minimum of five reinforcers per session. After each session, mice were brought back to their home cages.
2.6.3. Three addiction-like criteriaThe development of addictive-like behaviors was evaluated at the end of the training sessions based on three addiction-like criteria that summarize the addiction hallmarks according to the DSM-5 (29, 30). The addiction-like score developed was then attributed based on the results of these three criteria, determined by the respective behavioral test:
2.6.3.1. Persistence of responseNon-reinforced active responses during the 15 min drug-free period were measured as persistence of drug-seeking behavior. Mice were scored on the three consecutive days before the progressive ratio (PR).
2.6.3.2. MotivationThe PR schedule of reinforcement evaluated the motivation for WIN 55,212-2. The responses required to receive one single drug infusion escalated following this series: 1, 5, 12, 21, 33, 51, 75, 90, 120, 155, 180, 225, 260, 300, 350, 410, 465, 540, 630, 730, 850, 1,000, 1,200, 1,500, 1,800, 2,100, 2,400, 2,700, 3,000, 3,400, 3,800, 4,200, 4,600, 5,000, and 5,500. The breaking point, the maximal number of responses the animal performs to obtain one infusion, defined as the motivation value, corresponds to the last ratio completed. The duration of the PR session was maximum 4 h-long or until mice stopped responding to any nose-poke within 1 h.
2.6.3.3. CompulsivityResistance to punishment, defined as compulsive-like behavior, corresponded to the maintenance of active responding behavior despite its association with a negative consequence. It was measured by the total number of shocks obtained in a 50 min shock test, during which each drug delivered was associated with a foot-shock-induced punishment. This shock session was performed after a stabilizing FR2 self-administration session following the PR test. Mice were placed in a different operant box than the one usually used for operant training. Then, mice underwent a FR2 self-administration schedule of reinforcement for 50 min composed of two schedule changes: after nose-poking once in the active hole, mice received an electric foot-shock (0.18 mA, 2 s), while if they performed a second active nose-poke, the electric foot-shock was paired with the drug delivery and the associated cue light. After the time-out period that follows drug delivery, the latter schedule was reinitiated. In parallel, if the second response was not completed within a min after completing the first response, the sequence was also reinitiated.
2.6.4. Establishment of mice subpopulationsAfter performing the three behavioral tests, mice were categorized into “addicted” and “non-addicted” animals based on the number of achieved positive criteria. A mouse was considered positive for an addiction-like criterion when the score of the behavioral test was equal to or beyond the 75th percentile of the normal distribution of the vehicle group. Mice that achieved 2 or 3 criteria were considered “addicted” and categorized as vulnerable, whereas those reaching 0 or 1 criteria were considered “non-addicted” and categorized as resilient.
2.6.5. Extinction and parameters related to cravingOnly mice with patent catheters that achieved all acquisition criteria continued to the extinction phase. After thiopental testing, mice were allowed to rest for 1 day, during which they underwent a 2-h locomotion test in individual locomotor activity boxes (10.8 × 20.3 × 18.6 cm, Imetronic, Pessac, France) equipped with infrared sensors to detect locomotor activity and an infrared plane to detect rearings.
During the extinction period, neither WIN 55,212-2 infusions nor the associated environmental cues were delivered after nose-poking in the active hole. Mice were exposed to 2-h daily sessions for 12 consecutive days in the same operant chamber as the self-administration sessions. During this period, the extinction criterion was achieved when active responses were <35% of the mean responses obtained during the last 3 days of WIN 55,212-2 self-administration across three consecutive extinction sessions. Only mice that met the extinction criterion were evaluated for reinstatement. Two parameters related to craving were evaluated before and after this extinction period:
2.6.5.1. Resistance to extinctionNumber of active responses in 2 h during the first extinction session. Animals with significant sensitivity to drug withdrawal would increase their resistance to extinction by enhancing the number of active nose-pokes to seek the drug when access is prevented for 2 h the first time after the training sessions (31–33).
2.6.5.2. Drug-seeking behavior measured by cue-induced reinstatement of WIN 55,212-2-seeking behaviorThe day after reaching the extinction criterion, we performed a single cue-induced reinstatement test in the same operant chamber, in order to test reinstatement of drug-seeking behavior upon exposure to the environmental stimuli. The cue test was conducted under the same conditions used in the acquisition phase, except that active responding was not reinforced by the drug. Mice were exposed to a 90-min FR2 session, where the first 60 min were similar to an extinction session, but in the last 30 min, nose-poking in the active hole led to the presentation of all associated environmental cues (cue light, pump noise, and priming injection light), but not the delivery of WIN 55,212-2.
2.6.6. Behavioral tests to evaluate addiction-like phenotypic traitsTwo additional phenotypic traits were also evaluated as factors of vulnerability to addiction-like behavior:
2.6.6.1. ImpulsivityNon-reinforced active responses during the time-out periods (10 s) after each drug delivery were used as measures of impulsivity-like behavior, which indicated the inability to end a response once it is initiated. The three consecutive days before the PR test were considered for this criterion. Mice were then categorized as high-impulsive (HI) (score above the median) or low-impulsive (LI) (score below the median), as previously described (34, 35). The different subpopulations were split according to the median since impulsivity data was not normally distributed.
2.6.6.2. Sensitivity to rewardThe number of reinforcers obtained in 2-h sessions during the last three consecutive FR2 operant conditionings maintained by WIN 55,212-2. Animals with higher levels of sensitivity to reward will obtain a higher number of reinforcers.
2.7. Gene expression analysis 2.7.1. Tissue preparationTissue collection was performed immediately after the cue-induced reinstatement test. Mice were euthanized by cervical dislocation. Immediately afterward, brains were extracted from the skull and processed rapidly on ice. The medial prefrontal cortex (mPFC), nucleus accumbens (NAc), dorsal striatum (DS), and hippocampus (HPC) were isolated according to the following coordinates from the Paxinos and Franklin atlas (36): (mPFC) AP +1.98 mm; (NAc) AP +1.94 mm; (DS) AP +0.62 mm; (HPC) AP −2.92 mm. Samples were placed in individual tubes, frozen on dry ice and stored at −80°C until RNA isolation. The remaining brain parts from the same animals were also frozen on dry ice and stored at −80°C.
2.7.2. RNA extractionTotal RNAs from the mPFC, NAc, DS, and HPC were extracted using TRIzol™ (Thermo Fisher Scientific, USA) for subsequent RT-PCR analysis. Briefly, tissues were homogenized in TRIzol reagent and, after adding chloroform, the aqueous phase was collected and incubated in isopropanol to isolate the RNA. Samples were kept in dry ice meanwhile to ensure their integrity. Tubes were then centrifuged and pellets were resuspended in 75% ethanol, before another centrifugation. Supernatants were carefully discarded to air dry the pellet for at least 2 h. Finally, pellets were resuspended in RNase-free water, incubated for 15 min at 55–60°C and stored at −80°C. RNA concentrations were quantified by Nanodrop (NanoDrop One, Thermo Fisher Scientific, USA).
2.7.3. Quantitative polymerase chain reaction (qPCR)The quantitative conversion of the extracted RNA into single-stranded complementary DNA (cDNA) was performed using random primers included in the High Capacity cDNA Reverse Transcription (RT) kit (Applied Biosystems, 4390778, Thermo Fisher Scientific, USA). Real-time PCR analysis was carried out with the following primers (Sigma-Aldrich, USA): drd1: forward “AGATTGACCAGGAAGAGGCC,” reverse “GCAATCCAAGCCATACCAGG”; drd2: forward “CCATCTC TTGCCCACTGCTCTTTGG,” reverse “GGTGACGATGAAGGG CACGTAGAAC”; adora2a: forward “CGTCACCAACTT CTTCGTGG,” reverse “GCTGAAGATGGAACTCTGCG”; cnr1: forward “CCTGGGAAGTGTCATCTTTGT,” reverse “GGTAACCCCACCCAGTTTGA.” These primers were used in combination with the PowerSYBR Green PCR MasterMix kit (Applied Biosystems, Thermo Fisher Scientific, USA). Assays were analyzed with the QuantStudio™ 12K Flex real-time PCR system (Applied Biosystems, Thermo Fisher Scientific, USA). Relative expression of mRNAs was determined after normalization with a housekeeping gene using the ΔΔCt method. All validated differentially expressed genes were normalized using β-actine. The gene expression of β-actine was measured as housekeeping gene in these samples using quantitative polymerase chain reaction (qPCR) to verify that the expression of these genes was not affected by the operant model used.
2.8. Statistics 2.8.1. Statistical analysis of behavioral dataThe number of mice (n) in each experimental condition is indicated in figure legends. All statistical comparisons were performed with SPSS (IBM, version 25). Comparisons between two groups were performed by Student’s t-test or U Mann–Whitney test depending on the distribution defined by the Shapiro–Wilk test. ANOVA with repeated measures (or Friedman test when non-parametric) was employed when required to test the evolution over time, with subsequent post hoc analysis (Fisher LSD). Two-way ANOVA with subsequent post hoc analysis (Tukey) was used for multiple group comparisons. The Pearson correlation coefficient was performed to analyze the relationship between values in each addiction-like criterion and the final criteria achieved. The Chi-square analysis was used to compare the percentage of “addicted” with the “non-addicted” mice. Results were expressed as individual values with the median and the interquartile range or with the mean ± SEM, specified in the figure legend. A p-value < 0.05 was used to determine statistical significance.
The sample size was calculated based on the power analysis. The significance criterion (alpha) was set at 0.050 and the statistical test used was the two-sample t-test. With the sample size of 14–16 mice per group, our studies achieved a power superior to 80%. Supplementary Tables 1–7 give details of the statistical results for the data presented in the figures.
2.8.2. Principal component analysisThe principal component analysis (PCA) was performed to evaluate the multidimensional behavioral data by reducing it to fewer dimensions to observe trends, jumps, clusters, and outliers. PCA and varimax rotation were conducted using the three addiction-like criteria, the two parameters related to craving and the two phenotypic traits considered as vulnerability factors of addiction-like behavior, and dimensionality was reduced to the minimum number of components that best explain and maximize the variance present in the data set. An eigenvalue greater than 1 was set as the selecting criterion.
3. Results 3.1. Effects of THC pre-treatment during adolescence on the operant model of cannabinoid addiction-like behavior using WIN 55,212-2 self-administration training in adulthoodWe explored the effects of THC exposure in adolescent mice on specific behavioral signatures of cannabis addiction-like behavior in adulthood. Mice were trained to acquire an operant self-administration conditioning maintained by iv infusions of WIN 55,212-2 under an FR1 schedule of reinforcement during 5 sessions followed by 5 sessions under FR2, for a total of 10 consecutive sessions (Figure 1A). Under the FR1 schedule, no significant differences were found between THC and vehicle pre-treated mice, suggesting similar reinforcing effects of WIN 55,212-2 (Repeated measures ANOVA, F(1,28) = 0.00, n.s., Figure 1B). When the requirement was increased to FR2, THC pre-treated mice showed a tendency to obtain a higher number of reinforcers (mean ± SEM of the last three operant sessions = 15.7 ± 3.18) compared to vehicle pre-treated mice (mean ± SEM of the last three operant sessions = 11.81 ± 1.31), which was maintained until the end of the sessions (Repeated measures ANOVA, F(1,28) = 1.00, n.s.). The acquisition of operant learning was equivalent between groups (percentage of mice having acquired the behavior: 58.82% of vehicle and 50% of THC, Chi-square, C-S = 0.45, n.s.).
Figure 1. THC administration during adolescence led to the development of an addictive-like phenotype after WIN 55,212-2 operant self-administration in adult mice. (A) Timeline of the experimental sequence of the WIN 55,212-2 self-administration mouse model. (B) Number of infusions obtained by vehicle and THC pre-treated groups during 2 h of operant self-administration maintained by intravenous infusions of WIN 55,212-2 under both FR1 and FR2 schedules of reinforcement (mean ± SEM, repeated measures ANOVA). (C–E) Mice presented similar responses in the three addiction-like criteria tests (individual data with median and interquartile range). (C) Persistence of response: number of active nose-poke responses during the 15 min drug-free period (U Mann–Whitney). (D) Motivation: breaking point determined during a 4 h progressive schedule of reinforcement represents the maximal number of responses that an animal is able to emit to obtain one drug infusion (Student’s t-test). (E) Compulsivity: number of shocks received following the schedule described in section “Materials and methods”, reflecting the compulsivity level of each group (U Mann–Whitney). The dashed horizontal line indicates the 75th percentile of the distribution of the group, used as the threshold to consider a mouse positive for one criterion. “Addicted” mice are represented in gray-filled circles for the vehicle group and green-filled circles for the THC group. (F) Percentage of mice categorized as “addicted” (Chi-square). (G–I) Behavioral tests of the three addiction-like criteria when separated between “addicted” and “non-addicted” (individual data with median and interquartile range, Student’s t-test for persistence and motivation, U Mann–Whitney for compulsivity, *p < 0.05). Pearson correlations between individual values of addiction-like criteria, and (J) non-reinforced active responses in 15 min, (K) breaking point in 4 h, and (L) number of shocks in 50 min (nVehicle = 16, nTHC = 14; statistical details are included in Supplementary Table 1).
3.2. Mouse model of addiction-like behavior using WIN 55,212-2 self-administration and THC pre-treatment during adolescenceAfter the operant training, the addiction-like behaviors were evaluated, as explained above. No significant differences were found between THC and vehicle pre-treated mice in any of the addiction-like criteria, namely, persistence of response, motivation, and compulsivity (Figures 1C–E). As expected, extreme subpopulations that present a high persistence of response, motivation, and compulsivity were observed in both groups. Additionally, positive correlations between the number of responses to each addiction-like criterion (non-reinforced active responses, breaking point or number of shocks) and the number of addiction-like criteria achieved were found in the vehicle group, while the THC group presented only a positive correlation for the motivation (Pearson correlations, p < 0.05, p < 0.01, Figures 1J–L), where “addicted” mice showed the highest values. Mice individually classified into the “addicted” group following the previously described criteria were 37.5% in the vehicle pre-treated group and 35.71% in mice receiving THC during adolescence (Chi-square, C-S = 0.019, n.s., Figure 1F). Based on this classification, addiction-like behaviors were compared, revealing that vehicle pre-treated “addicted” mice had higher persistence of response than “non-addicted” mice (Student’s t-test, t = −2.72, p < 0.05, Figure 1G), whereas no significant differences were found between “addicted” and “non-addicted” mice in terms of motivation and compulsivity regardless of the treatment (Figures 1H, I).
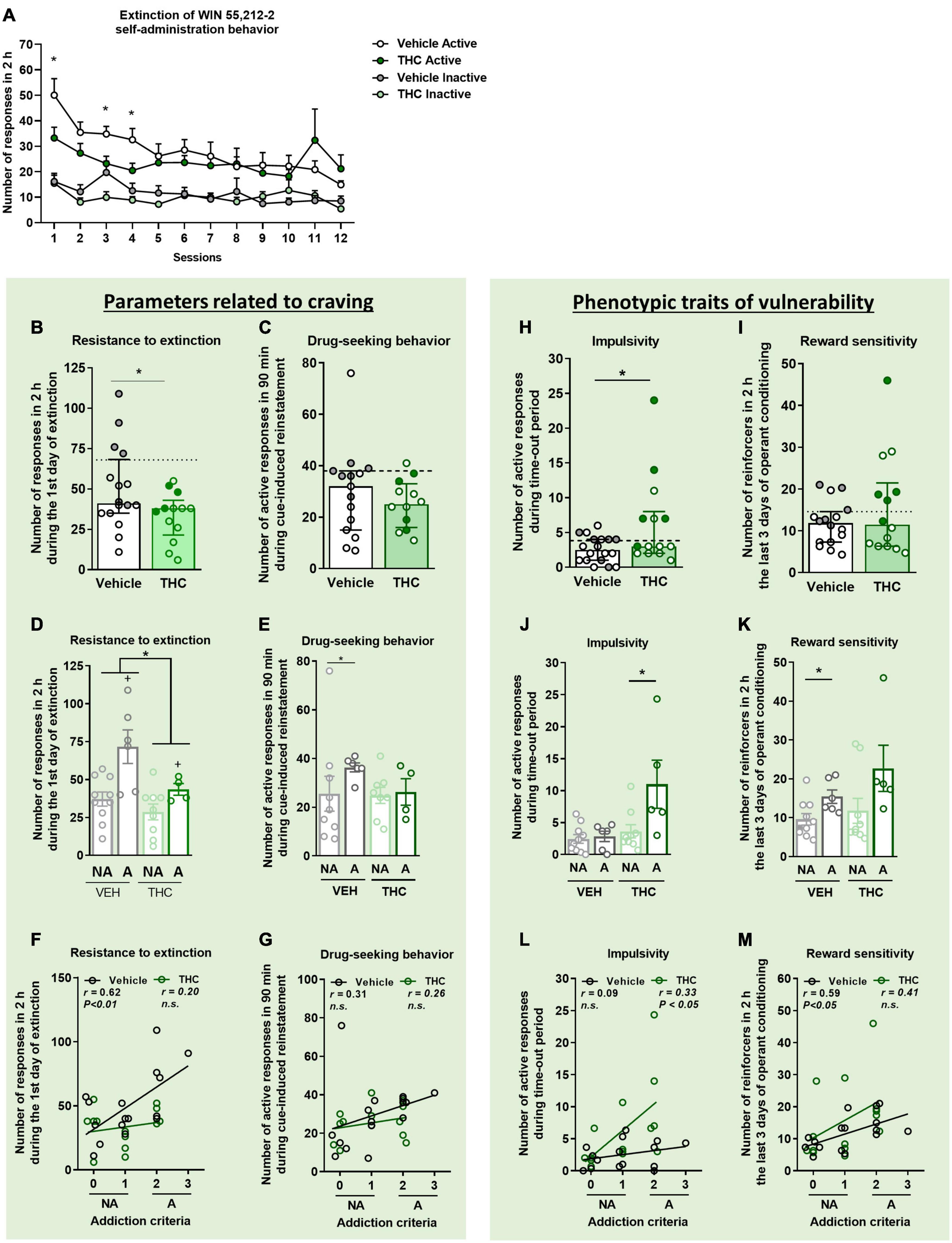
3.3. THC pre-exposure during adolescence decreased the resistance to extinction in adulthoodMice then underwent 12 sessions of extinction. The extinction trend of both groups was similar, indicating that both displayed the same ability to extinguish the self-administration behavior (Repeated measures ANOVA, F(1,26) = 1.20, n.s., Figure 2A). A total of 26.7% of vehicle and 8.3% of THC pre-treated mice (Chi-square, C-S = 2.06, n.s.) acquired the extinction criteria, as previously described.
Figure 2. THC administration during adolescence favored the parameters related to craving and the phenotypic vulnerability traits to addiction-like behavior after WIN 55,212-2 operant self-administration in adult mice. (A) Extinction pattern of the WIN 55,212-2 self-administration behavior (mean ± SEM, repeated measures ANOVA). (B,C) Behavioral tests of the two parameters related to craving (individual data with median and interquartile range). (B) Resistance to extinction: number of active nose-poke responses during the first 2-h extinction session is significantly higher in vehicle compared to THC pre-treated mice (Student’s t-test, *p < 0.05). (C) Drug-seeking behavior with the cue-induced reinstatement after abstinence: number of active responses performed during the 90 min cue-induced drug-seeking test performed after extinction (Student’s t-test). The dashed horizontal line indicates the 75th percentile of distribution of the group, used as the threshold to consider a mouse positive for one criterion. “Addicted” mice are represented in gray-filled circles for the vehicle group and green-filled circles for the THC group. (D,E) Behavioral tests regarding the parameters related to craving when separating between “addicted” and “non-addicted” mice (individual data with mean ± SEM). (D) In resistance to extinction, an effect of THC pre-treatment was detected between vehicle and THC groups as well as an effect of phenotype between “addicted” and “non-addicted” mice (two-way ANOVA, Treatment effect = *p < 0.05, Phenotype effect = +p < 0.01). (E) Higher drug-seeking behavior was observed in “addicted” vehicle pre-treated animals compared to “non-addicted” (U Mann–Whitney, *p < 0.05). Pearson correlations between individual values of addiction-like criteria, and (F) number of responses in 2 h during the first day of extinction and (G) number of responses in 90 min during the cue-induced reinstatement test. (H,I) Behavioral tests used to evaluate phenotypic traits considered to be factors of vulnerability to addiction-like behavior (individual data with median with interquartile range). (H) Impulsivity: number of responses to the active nose-poke during the 10 s time-out period (Student’s t-test, *p < 0.05). (I) Reward sensitivity: number of reinforcers performed to the active nose-poke during the 2 h of the last three sessions of self-administration (U Mann–Whitney). (J,K) Behavioral tests of the phenotypic traits showing higher reward sensitivity in “addicted” mice pre-treated with vehicle compared to “non-addicted”, whereas this difference was observed in the THC group for impulsivity (individual data with mean ± SEM, U Mann–Whitney, *p < 0.05). (L,M) Pearson correlations between individual values of addiction-like criteria, and (L) number of active responses during time-out period and (M) number of reinforcers in 2 h in the last three consecutive days of operant training (nVehicle = 16, nTHC = 14; statistical details are included in Supplementary Table 2).
Two parameters closely related to craving and reinstatement of drug-seeking were evaluated. First, the pattern of activity on the first extinction day was evaluated every 10 min as a measurement of resistance to extinction (Supplementary Figure 2). All groups highly responded to the previous active nose-poke at the beginning of the session, meaning that mice were seeking the reinforcer and the cue light associated with it. However, as non-reinforced sessions continued, we see that this behavior diminished, which is characteristic of an extinction behavior. THC pre-treated mice responded less during the first extinction session than vehicle pre-treated mice (Student’s t-test, t = −2.18, p < 0.05, Figure 2B). In a two-way ANOVA analysis, this global treatment effect was also observed between THC and vehicle pre-treated mice. When separating groups into “addicted” and “non-addicted,” an increased resistance to extinction was observed in “addicted” mice compared to “non-addicted” [two-way ANOVA, Treatment effect: F(1,25) = 6.872, p < 0.05, Phenotype effect: F(1,25) = 12.52, p < 0.01, Interaction: F(1,25) = 1.998, n.s., Figure 2D]. In the vehicle group, active nose-pokes in the first session were very high and decreased across sessions until reducing by 70% in the last session compared to the first one, whereas THC pre-treated mice decreased only by 30% (Figure 2A). In addition, active nose-poking by both groups was significantly higher than inactive nose-poking (Repeated measures ANOVA, F(1,48) = 15.26, p < 0.001, post-hoc DMS test: Actives vs. Inactives: p < 0.001 in all the comparisons). On the first, third and fourth extinction sessions, vehicle pre-treated mice showed less extinction responding compared to mice pre-treated with THC (Student’s t-test, p < 0.05).
The reinstatement of drug-seeking behavior was evaluated after mice achieved extinction. No significant differences were found between vehicle and THC pre-treated groups regarding the cue-induced reinstatement of drug-seeking behavior. Only a significant difference was revealed when comparing “addicted” and “non-addicted” mice pre-treated with vehicle. Indeed, “addicted” mice pre-treated with vehicle presented higher responses than vehicle “non-addicted” mice (U Mann–Whitney, U = 10.000, p < 0.05) (Figures 2C–E).
3.4. THC exposure during adolescence increased impulsivity-like behavior in adulthoodTwo additional phenotypic traits considered as factors of vulnerability to substance misuse were evaluated. THC pre-treated mice displayed higher impulsivity-like behavior compared to vehicle pre-treated mice (Student’s t-test, t = −2.17, p < 0.05, Figure 2H), an effect that was more pronounced in “addicted” mice pre-treated with THC (U Mann–Whitney, U = 6.500, p < 0.05, Figure 2J). Contrary, no significant differences were found between vehicle and THC pre-treated groups for the reward sensitivity during the last 3 days of operant training (Figure 2I), although “addicted” mice pre-treated with vehicle had more reward sensitivity compared to “non-addicted” (U Mann–Whitney, U = 10.000, p < 0.05, Figure 2K). These results were emphasized by the positive correlations found between the number of responses to each addiction-like criterion and the number of addiction-like criteria achieved (Pearson correlations, Figures 2F, G, L, M).
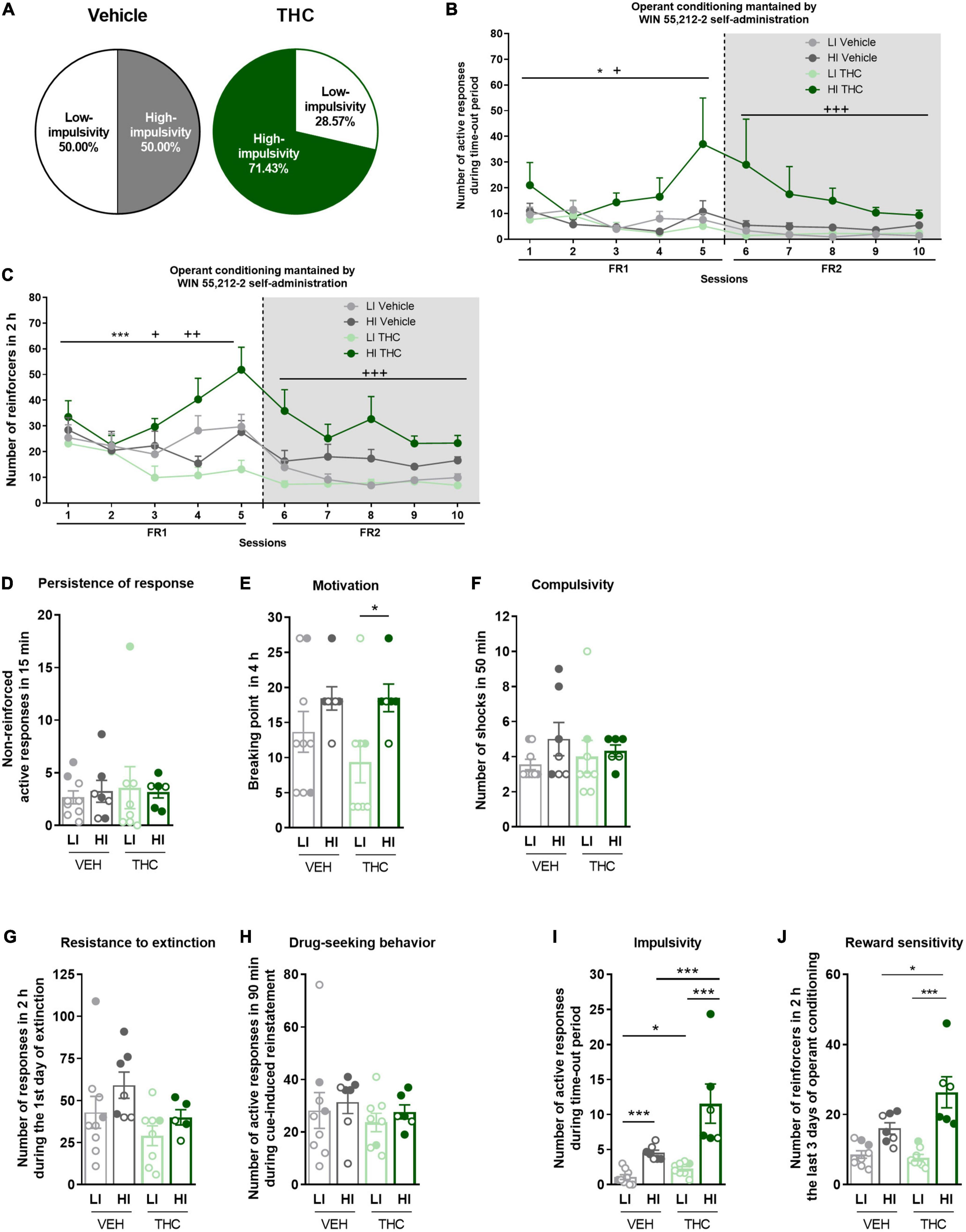
3.5. High- and low-impulsive subgroups reveal behavioral differences in adult mice after WIN 55,212-2 self-administrationTo further investigate the enhanced impulsive behavior promoted by THC pre-exposure, mice were categorized into high-impulsive and low-impulsive subgroups defined according to the median (see section “Materials and methods”). We find 50.0% of high-impulsive mice in the vehicle group and 71.43% in the THC group (Chi-square, C-S = 2.571, n.s., Figure 3A). High-impulsive mice pre-treated with THC showed a higher number of active responses during the time-out period compared to high-impulsive mice pre-treated with vehicle, as well as to low-impulsive mice under both the FR1 (Repeated measures ANOVA, F(1,26) = 3.46, p < 0.05, post-hoc DMS test: HI THC vs. HI vehicle: p < 0.05, HI THC vs. LI vehicle: p < 0.05, Figure 3B) and FR2 schedules (Repeated measures ANOVA, F(1,26) = 7.86, p < 0.001, post-hoc DMS test: HI THC vs. HI vehicle: p < 0.01, HI THC vs. LI vehicle: p < 0.001). Moreover, high-impulsive mice pre-treated with THC obtained a higher number of reinforcers during the operant conditioning when tested under the FR1 schedule compared to THC pre-treated low-impulsive mice (Repeated measures ANOVA, F(1,26) = 3.44, p < 0.05, post-hoc DMS test: HI THC vs. LI THC: p < 0.01, Figure 3C), as well as to high-impulsive mice pre-treated with vehicle during sessions 4 (U Mann–Whitney, U = 1.000, p < 0.01) and 5 (U Mann–Whitney, U = 7.000, p < 0.05). Also, high-impulsive mice pre-treated with THC presented higher responses in the FR2 schedule compared to high-impulsive mice pre-treated with vehicle and to both low-impulsive mice (Repeated measures ANOVA, F(1,26) = 12.73, p < 0.001, post-hoc DMS test: HI THC vs. HI vehicle: p < 0.01, HI THC vs. LI vehicle: p < 0.001, and HI THC vs. LI THC: p < 0.001). Furthermore, high-impulsive mice presented higher motivation compared to low-impulsive mice only in the THC pre-treated group, whereas no significant results were revealed for other addiction-like criteria (U Mann–Whitney, U = 7.000, p < 0.05, Figures 3D–F).
Figure 3. Characterization of low-impulsive (LI) and high-impulsive (HI) subgroups. (A) Percentage of mice categorized as LI and HI (Chi-square). (B) HI mice pre-treated with THC showed a higher number of responses during the time-out period under both FR1 and FR2 schedules compared to HI mice pre-treated with vehicle, as well as to LI mice (repeated measures ANOVA, *p < 0.05 session effect in FR1, +p < 0.05 treatment effect in FR1, +++p < 0.001 treatment effect in FR2). (C) HI mice pre-treated with THC showed higher acquisition of the operant conditioning under FR2 schedule compared to HI mice pre-treated with vehicle, as well as to LI mice (Repeated measures ANOVA, ***p < 0.001 session effect in FR1, ++p < 0.01 treatment × session effect in FR1, +p < 0.05 treatment effect in FR1; +++p < 0.001 treatment effect in FR2). (D–F) Behavioral tests of the three addiction-like criteria when separated between LI and HI (mean ± SEM, U Mann–Whitney, *p < 0.05). (G–J) Behavioral tests of the (G,H) two parameters related to craving (Student t-test or U Mann–Whitney) and (I,J) the two phenotypic vulnerability traits when separated between LI and HI (mean ± SEM, impulsivity: Student t-test or U Mann–Whitney, reward sensitivity: two-way ANOVA with Tukey’s multiple comparisons test, *p < 0.05, ***p < 0.001) (nVehicle = 16, nTHC = 14; statistical details are included in Supplementary Table 3).
No significant differences were revealed in the parameters related to craving (Figures 3G, H). Both low-impulsive (U Mann–Whitney, U = 15.000, p < 0.05) and high-impulsive mice (U Mann–Whitney, U = 0.000, p < 0.001) receiving THC during adolescence show more extreme values of impulsivity compared to those pre-treated with vehicle (Figure 3I). As expected, high-impulsive mice have higher impulsivity than low-impulsive mice regardless of the treatment (U Mann–Whitney, U = 0.000, p < 0.001 for vehicle, and U Mann–Whitney, U = 0.000, p < 0.001 for THC). In terms of reward sensitivity, a phenotypic difference was found between low-impulsive and high-impulsive mice, as well as an effect of the treatment and of the interaction between both [two-way ANOVA, Phenotype effect: F(1,26) = 39.38, p < 0.001, Treatment effect: F(1,26) = 5.194, p < 0.05, Interaction: F(1,26) = 7.314, p < 0.05, Figure 3J]. High-impulsive animals pre-treated with THC showed higher reward sensitivity compared to low-impulsive mice pre-treated with THC (Tukey’s multiple comparisons, p < 0.001), as well as to high-impulsive mice pre-treated with vehicle (Tukey’s multiple comparisons, p < 0.05).
3.6. THC pre-treatment did not produce major long-term behavioral or somatic alterationsΔ9-Tetrahydrocannabinol exposure has been reported to produce a series of behavioral and somatic alterations (37). Several behavioral and somatic responses (body weight, food intake, and locomotor activity) were monitored throughout the experiment in order to verify that THC pre-treatment during adolescence at the dose of 5 mg/kg did not produce any long-term effect in adulthood that could bias our self-administration protocol. No significant differences were observed between THC and vehicle pre-treated mice in body weight and food intake during the whole operant sequence, even though the weight gain of mice pre-treated with THC was apparently lower than that of vehicle pre-treated mice during the adolescent treatment (Supplementary Figures 1A–C). This is in accordance with previous studies employing similar protocols that have shown that THC treatment may reduce weight gain in adolescent mice compared to vehicle pre-treated animals (27, 38). Moreover, no significant differences were observed between groups in the number of beam breaks per 10 min during 120 min (Supplementary Figure 1D), which supports the absence of major behavioral alteration promoted by THC pre-exposure.
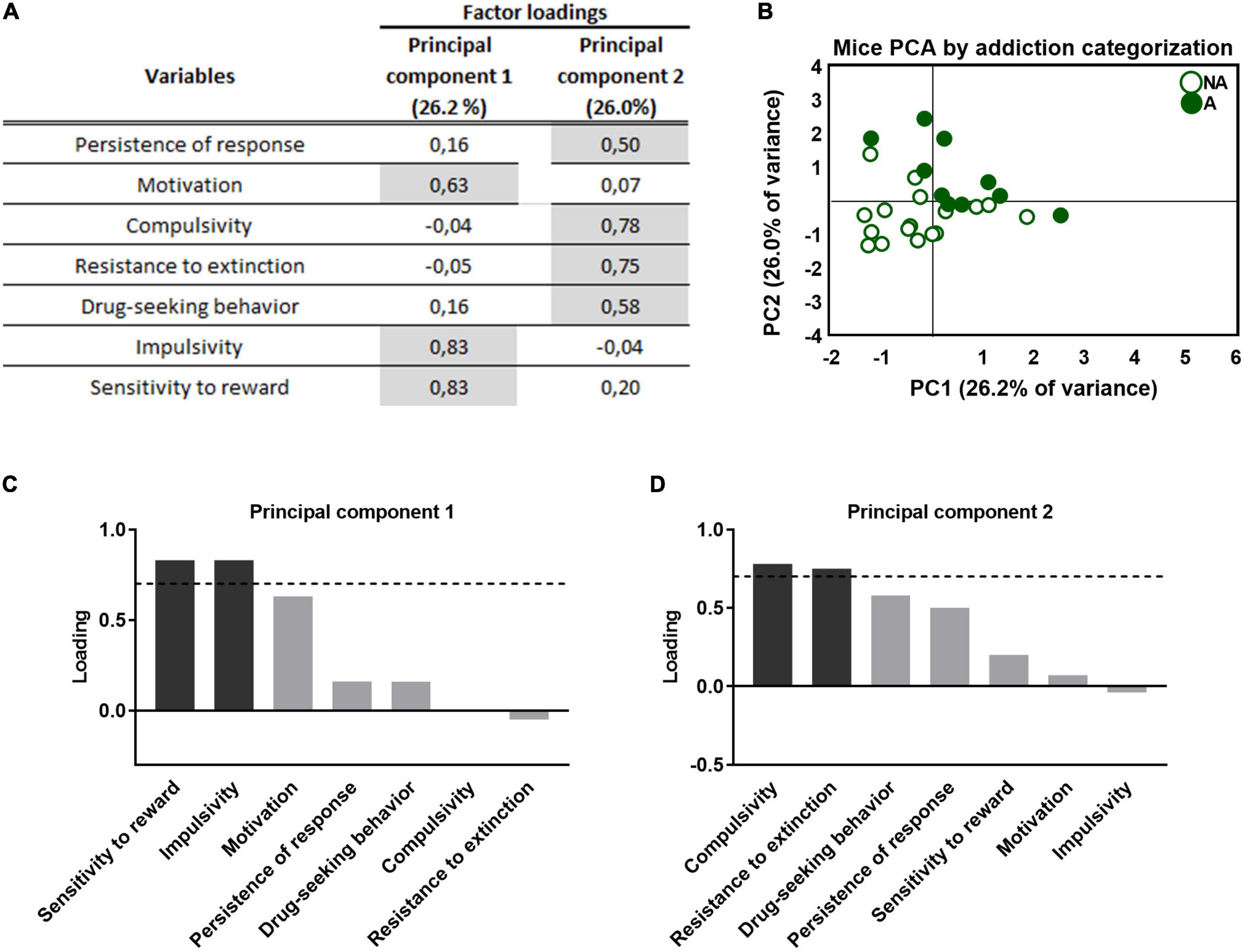
3.7. Principal component analysis of THC exposure during adolescence in self-administration of WIN 55,212-2 in adulthoodPrincipal component analysis was used to characterize whether the behavioral outcomes previously described could be reduced to fewer dimensions. All addiction-like criteria, parameters related to craving, and phenotypic traits were taken into account. This analysis separated two clear clusters representing the separation between the “addicted” and “non-addicted” groups and both populations present clear behavioral differences that allow for a distinction (Figure 4B). Component 1 accounts for 26.2% of the variance (Figures 4A, B) and has strong loadings (>0.7) from three behavioral variables: motivation, impulsivity, and reward sensitivity. The second component is orthogonal to component 1, accounts for 26.0% of the variance, and comprises four variables: the criteria of persistence of response, compulsivity, resistance to extinction, and drug-seeking behavior. Impulsivity participates more in the first component, while compulsivity is more critical in the second component (Figures 4C, D), resembling the sequential feature of the transition from impulsivity to compulsivity that has been described in addiction (39). Also, motivation and compulsivity belong to different components, as the neural substrate of each addiction criterion is different (15).
Figure 4. Principal component analysis of the effect of adolescent THC exposure in WIN 55,212-2 operant self-administration in adulthood. (A) Factor loadings of principal component 1 (PC1) and principal component 2 (PC2) for all variables studied. (B) Individual mice clustered according to addiction or non-addiction in the space yielded by the 2 PCA components, which accounted for the maximum data variance, with factor loadings of 26.2% for PC1 and 26.0% for PC2. (C,D) Order of factor loading of the different variables in PC1 and PC2. The dashed horizontal line marks loading greater than 0.7, mainly contributing to the component. In regards to the addiction-like criteria, a dissociation between motivation, mainly contributing to PC1, and compulsivity, mainly contributing to PC2, can be observed. Moreover, impulsivity and reward sensitivity weighted more in the PC1, while resistance to extinction and drug-seeking weighted more in the PC2.
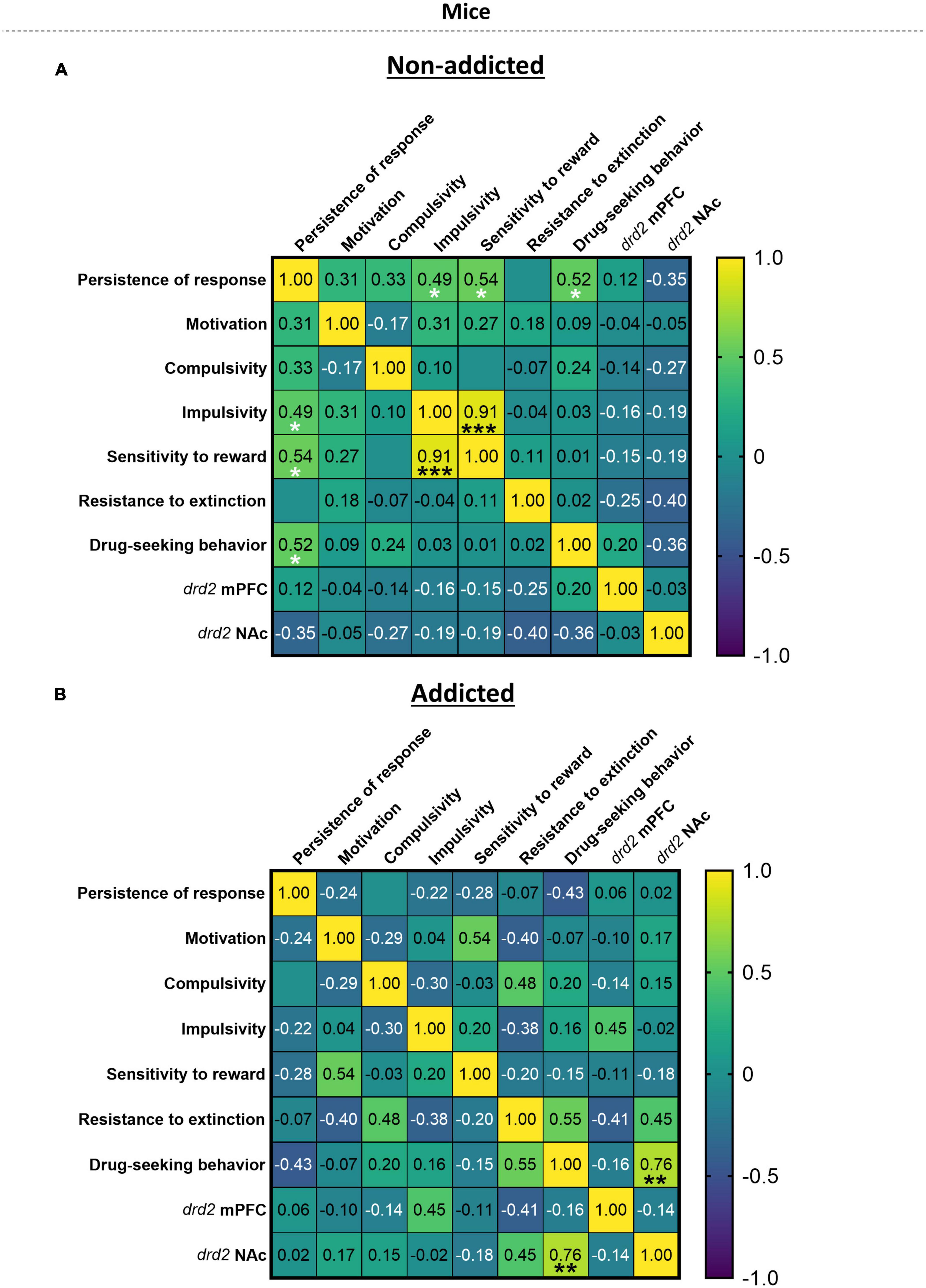
3.8. Correlation heatmap between the addiction-like criteria, parameters related to craving and vulnerability phenotypic traitsWhen representing the addiction-like criteria, parameters related to craving and phenotypic traits in a heat map (Figure 5), we revealed significant correlations in “non-addicted” animals between response persistence and impulsivity (r = 0.49, p < 0.05), persistence of response and sensitivity to reward (r = 0.54, p < 0.05), persistence of response and drug-seeking behavior (r = 0.52, p < 0.05), and sensitivity to reward and impulsivity (r = 0.91, p < 0.05). In “addicted” animals, a significant correlation was revealed between drug-seeking behavior and drd2 NAc (r = 0.76, p < 0.01).
Figure 5. Correlation heatmap of the variables of cannabis addiction-like criteria, parameters related to craving and vulnerability phenotypic traits. (A,B) Pearson correlations between the three addiction-like criteria, the two parameters related to craving and the two phenotypic traits in both (A) “non-addicted” and (B) “addicted” groups (Pearson correlation, *p < 0.05, **p < 0.01, ***p < 0.001).
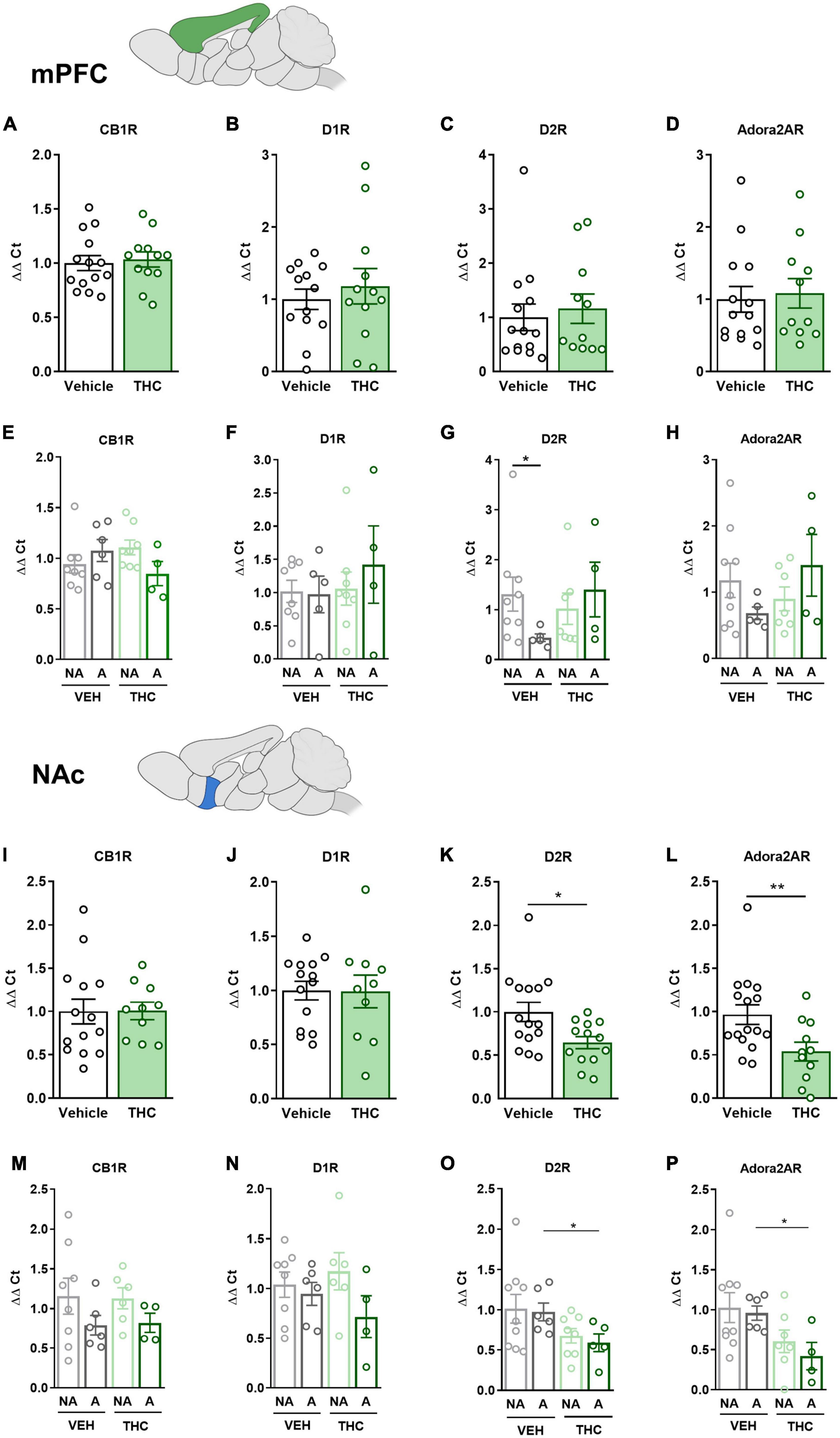
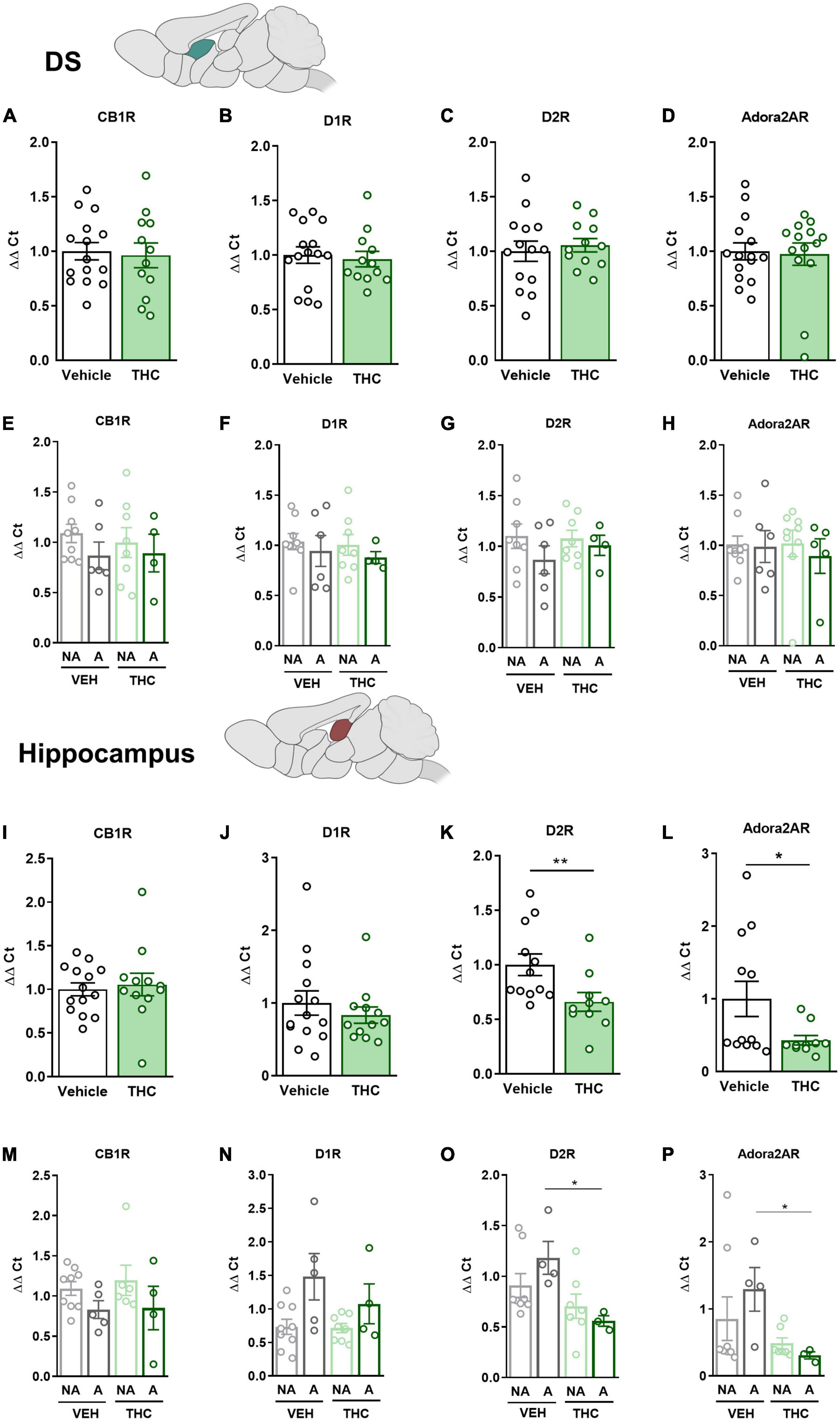
3.9. THC exposure during adolescence produced a downregulation of the drd2 and adora2a gene expression in the NAc and HPCThe mPFC, NAc, DS, and HPC were extracted at the end of the operant evaluation to study potential neurobiological alterations promoted by THC pre-exposure during adolescence. In a previous study, the expression of drd1, drd2, and adora2a genes was found to be upregulated in food- and cocaine-addicted mice after operant self-administration paradigms (40). Moreover, the cnr1 gene has been revealed to be involved in addictive disorders (41). Consequently, RT-qPCR assays were performed to compare the differential gene expression of these candidate genes in areas of the mesocorticolimbic circuit. No significant differences between THC and vehicle pre-treated mice were revealed in the mPFC (Figures 6A–D). However, a downregulation in the expression of the drd2 gene was observed in “addicted” mice pre-treated with vehicle compared to “non-addicted” in this brain area (U Mann–Whitney, U = 6.000, p < 0.05, Figure 6G). No significant differences were found for cb1 (Figure 6E), drd1 (Figure 6F) or adora2a (Figure 6H). In the NAc, drd2 and adora2a expression was significantly downregulated in THC pre-treated mice compared to vehicle (Student’s t-test, t = 2.60, p < 0.05 and t = 2.83, p < 0.01, respectively, Figures 6K, L), and these differences were specifically found in “addicted” mice (Student’s t-test, t = 2.44, p < 0.05 and t = 3.10, p < 0.05, respectively, Figures 6O, P). Similarly, drd2 and adora2a expression was downregulated in THC pre-treated mice in the HPC (Student’s t-test, t = 2.54, p < 0.01 and U Mann–Whitney, U = 31.000, p < 0.05, respectively, Figures 7K, L), and these differences were specifically found in “addicted” mice (Student’s t-test, t = 3.18, p < 0.05 and t = 3.06, p < 0.05, respectively, Figures 7O, P). No significant differences were found for cb1 or drd1 in the NAc (Figures 6I, J, M, N) or the HPC (Figures 7I, J, M, N). No significant differences were found in the DS (Figures 7A–H).
Figure 6. drd1, drd2, adora2a, and cnr1 gene expression in the medial prefrontal cortex (mPFC) and nucleus accumbens (NAc) after adolescent THC exposure in WIN 55,212-2 operant self-administering adult mice. Gene expression measured by the ΔΔCt after RT-qPCR of cnr1, drd1, drd2, and adora2a in the PFC: (A–D) between vehicle and THC pre-treated mice respectively (mean ± SEM, Student’s t-test for CB1R and D1R, U Mann–Whitney for D2R and Adora2AR), (E–H) between “addicted” and “non-addicted” mice respectively (mean ± SEM, Student’s t-test or U Mann–Whitney, *p < 0.05). Gene expression measured by the ΔΔCt after qPCR of cnr1, drd1, drd2, and adora2a in the NAc: (I–L) between vehicle and THC pre-treated mice respectively (mean ± SEM, Student’s t-test, *p < 0.05, **p < 0.01), (M–P) between “addicted” and “non-addicted” mice respectively (mean ± SEM, Student’s t-test or U Mann–Whitney, *p < 0.05) (nVehicle = 16, nTHC = 14; statistical details are included in Supplementary Table 4).
Figure 7. drd1, drd2, adora2a, and cnr1 gene expression in the dorsal striatum (DS) and hippocampus (HPC) after adolescent THC exposure in WIN 55,212-2 operant self-administering adult mice. Gene expression measured by the ΔΔCt after RT-qPCR of cnr1, drd1, drd2, and adora2a in the DS: (A–D) between vehicle and THC pre-treated mice respectively (mean ± SEM, Student’s t-test for CB1R, D2R, and Adora2AR, U Mann–Whitney for D1R), (E–H) between “addicted” and “non-addicted” mice respectively (mean ± SEM, Student’s t-test). Gene expression measured by the ΔΔCt after qPCR of cnr1, drd1, drd2, and adora2a in the NAc: (I–L) between vehicle and THC pre-treated mice respectively (mean ± SEM, Student’s t-test for CB1R, D1R, and D2R, U Mann–Whitney for Adora2AR, *p < 0.05, **p < 0.01), (M–P) between “addicted” and “non-addicted” mice respectively (mean ± SEM, Student’s t-test or U Mann–Whitney, *p < 0.05) (nVehicle = 16, nTHC = 14; statistical details are included in Supplementary Table 5).
4. DiscussionIn this study, we assessed the effects of THC exposure during adolescence on the vulnerability to develop cannabis addiction-like behavior and related behavioral alterations in adulthood by using a WIN 55,212-2 self-administration mouse model. We found that chronic exposure to THC from PND 37 to 57 increased impulsivity in adult mice, which was more pronounced in the subgroup of mice that developed the addiction-like criteria. Furthermore, THC treatment during adolescence increased the percentage of high-impulsive mice and favored the reinforcement of WIN 55,212-2 in this group. Moreover, high-impulsive mice pre-treated with THC presented an increased reward sensitivity compared to low-impulsive mice pre-treated with THC, as well as to high-impulsive mice pre-treated with vehicle. Inversely, pre-exposure to THC during adolescence decreased the resistance to extinguishing the operant behavior.
Impulsivity is a complex construct defined as a “predisposition toward rapid, unplanned reactions to internal and external stimuli without regard for the negative consequences of these reactions to themselves or others” (42) that is composed of motor and choice impulsivity (43–47). Motor impulsivity is described as the inability to inhibit behavior by changing the course of action or to stop a response once it is initiated (48). We have defined motor impulsivity as the non-reinforced active nose-pokes during the time-out periods that reflect the motor disinhibition, as previously described (47). Compulsivity has its roots in the signs associated with obsessive-compulsive disorder and might comprise repetitive behaviors in the face of adverse consequences after the loss of control (49). As such, we implemented a punishment-based behavioral test where perseverance in responding despite punishment defined compulsive behavior. Finally, reward sensitivity represents the tendency of individuals to approach a positive reinforcer (50). In this article, it is operationally defined as the mean of drug infusions obtained during the last three sessions of operant training.
Δ9-Tetrahydrocannabinol pre-treatment during adolescence increased impulsivity in adulthood. Although impulsive behavior may be a pre-existing trait that promotes the usage of drugs, acute and chronic cannabis consumption may result in behavioral changes, including increases in impulsivity, which may facilitate further drug use or the transition to use other drugs of abuse according to the gateway theory (51). In humans, marijuana use has been associated with increased impulsivity when given in an acute (52) or chronic manner (53). Concomitantly, individuals with CUD have been shown to present increased impulsivity (54). Interestingly, impulsivity is suggested to predispose compulsivity, which leads to drug abuse and addiction (55, 56). Indeed, it has been shown that high impulsivity predicts the transition from controlled to compulsive cocaine-taking in rats, as high impulsive rats displayed greater resistance to punishment of the cocaine-taking response compared to low impulsive rats, while a correlation analysis revealed, at the population level, that impulsivity predicts compulsivity (56). Compulsivity shares similarities with impulsivity in inhibitory control dysfunction, which relates to alterations in the mPFC networks to subcortical regions. However, impulsivity and compulsivity may be on a continuum, with compulsivity being repetitive and perseverative (57). In our study, THC pre-treatment during adolescence did not increase the percentage of mice reaching cannabinoid addiction-like criteria but increased the levels of impulsivity-like behavior. Furthermore, within this group, “addicted” mice showed the highest values of motor impulsivity compared to the rest of the groups. This result highlights that the interaction between THC adolescent exposure and impulsivity in addiction development should be explored further.
Our results are in accordance with observations suggesting an overrepresentation of highly impulsive individuals with substance abuse disorders and a high comorbidity between impulsive behavior and substance misuse (56). Indeed, the development of drug addiction would represent a progression from initial impulsivity mediated by the NAc to compulsive, habitual responding
留言 (0)