
記住我

Acute respiratory distress syndrome (ARDS) and sepsis are the biggest cause of mortality in critically ill patients with no specific pharmacological treatment.1 2 ARDS is a syndrome where heterogeneity of the underlying pathophysiological mechanisms presents a significant obstacle for translational research.3 4 Based on the circulating inflammatory biomarkers, patients with ARDS can be classified into subpopulations characterised by different patterns of inflammatory response.5 Using the method of latent class analysis, two broad biological phenotypes (hyper-inflammatory and hypo-inflammatory) were identified retrospectively in two large randomised clinical trials and in one observational study. The phenotypes had different clinical outcomes and responded differently to pharmacological and therapeutic interventions.6–9 Therefore, it is important to further investigate the underlying biological mechanisms responsible for differences in the inflammatory responses between ARDS phenotypes and where possible, consider the investigation of potential new therapeutics separately for each phenotype.
Mesenchymal stromal cells (MSCs) are being actively investigated as a cell-based therapy for ARDS.10 11 Clinical trials using MSCs in ARDS (including COVID-19-induced ARDS) are ongoing. Macrophages are key innate immune cells responsible for orchestrating inflammatory responses and are critical in driving inflammation and injury in ARDS.12 13 Our group and other investigators have demonstrated that macrophages are important cellular mediators of MSC immunomodulatory and anti-microbial effects in a range of inflammatory conditions including pneumonia and sepsis.14 15 Importantly, we have shown that depletion of alveolar macrophages results in the abrogation of the protective effects elicited by MSCs in the in vivo Escherichia coli pneumonia model of lung injury.16 Furthermore, we have demonstrated that in pre-clinical models of ARDS, MSCs reprogramme both human and murine macrophages towards M2-like, anti-inflammatory phenotype with enhanced phagocytic activity, an effect, that is, at least partially mediated by mitochondrial transfer resulting in macrophage metabolic reprogramming.16 17 One of the important questions, which still needs to be addressed is: how do MSCs modulate intracellular signalling in their target cells?
It is widely accepted that therapeutic effects of MSCs are mediated largely by their secretome. Importantly, MSC-derived extracellular vesicles can recapitulate many effects of MSCs themselves and are being developed as a cell-free therapeutic for multiple conditions including ARDS.18 19 MSC extracellular vesicles (EVs) are enriched in regulatory micro RNA (miRNA) content which has a potential for modulation of functional properties of recipient cells,20 however, the functional importance of individual miRNAs within the EV cargo remains largely unknown.
Suppressor of cytokine signalling (SOCS) proteins are inducible feedback inhibitors of the Janus kinase/signal transducers and activators of transcription (JAK/STAT) signalling pathway. The mammalian SOCS protein family consists of at least eight members, SOCS1–7 and CIS (cytokine‐inducible Src homology 2 protein).21 Data from transgenic animal studies suggest that these regulatory proteins play a key role in macrophage polarisation.22 Of particular interest is SOCS1, which has been shown to be a multifunctional inhibitor of the inflammatory response and capable of preventing activation of pathogen recognition receptors, cytokine receptors and receptors for growth factors.23 It has been demonstrated that SOCS1 expression is critical for the control of M2 polarisation both in vitro and in vivo.24 Additionally, mice containing a myeloid-specific deletion in SOCS1 have been reported to be more susceptible to sepsis.25 However, the role of SOCS1 protein in regulation of human macrophages in the context of ARDS and its contribution to the MSC therapeutic effect is currently unknown.
In the present study, we aimed to investigate the role of SOCS1 in the modulation of human macrophages by MSCs and decipher the mechanisms underpinning this effect.
We hypothesised that SOCS1 is crucial for the reprogramming of human macrophages and reduction of inflammation by MSCs in the ARDS environment and that MSCs modulate SOCS1 expression via the transfer of miRNAs in extracellular vesicles. We also hypothesised that therapeutic efficacy of MSC EVs may be enhanced by manipulating EV miRNA expression. Some of the results of these studies were previously reported in the form of abstracts.26 27
Materials and methodsDetailed methods are described in the online supplemental document.
Cell cultureHuman bone marrow-derived MSCs were acquired from the Institute for Regenerative Medicine at Texas A&M University, (Temple, Texas, USA) and the American Type Culture Collection (LGC Standards UK). These cells fulfil all requirements set by the International Society of Cellular Therapy for defining MSCs.28 Human monocyte-derived macrophages (MDMs) were generated using granulocyte macrophage colony-stimulating factor (GM-CSF) (R&D systems, UK) differentiation (10 ng/mL for 7 days) of monocytes from buffy coats obtained from the Northern Ireland Blood Transfusion Service.
Generation of MSC conditioned medium and extraction of MSC-derived EVsMSC conditioned medium (CM) was generated from MSCs cultured in RPMI-1640 with 1% fetal bovine serum (FBS) for 24 hours. For MSC EV isolation, MSCs were cultured in serum-free α-MEM-medium for 48 hours before EV extraction using ultracentrifugation as previously described.29 EVs were resuspended in phosphate-buffered saline (PBS) and characterised according to the International Society for Extracellular Vesicles30 guidelines (online supplemental figure 1).
Co-culture of MDMs with MSC CM and MSC EVsMDMs were treated with MSC CM or EVs in the presence of E. coli lipopolysaccharide (LPS) O111:B4 (Millipore) (10 ng/mL) or 10% plasma or 30% bronchoalveolar lavage fluid (BALF). Plasma samples used in the study were from patients recruited to HARP-2 study.31 These samples were previously classified into two phenotypes based on concentrations of plasma inflammatory biomarkers.7 Ten plasma samples representative of each phenotype were pooled and diluted in 1% complete medium to final concentration of 10% before use, plasma from healthy volunteers was used as a control. BALF samples were from HARP study,32 nine BALF samples were pooled to generate a stock and the pooled sample was then diluted to 30% in RPMI 1% FBS+Penicillin/Streptomycin (PS) before stimulation. Only baseline samples obtained prior to intervention were used for experiments. Ethical approval for use of patient samples for research was granted by the Office for Research Ethics Committees Northern Ireland.
Small RNA sequencing of EVsMSCs were exposed to pooled ARDS BALF (30%) for 24 hours,32 cell supernatants were collected for EV isolation, cells washed and RNA isolated. EVs were isolated from cell supernatants and BALF by ultracentrifugation. RNA was extracted using ‘miRNeasy’ Kit, (Qiagen). RNA integrity was assessed on Qubit RNA HS Assay Kit (Invitrogen). Small RNAs were converted to complementary DNA libraries using the NEXTFLEX Small RNA-Seq Kit V.3 (PerkinElmer). Quality control of the libraries and sequencing was performed by the Genomics Core Technology Unit at Queens University Belfast on a NextSeq 550 System (Illumina). FASTq files were uploaded onto CLC Genomics Workbench and analysed using the small RNA pipeline analysis tool (Qiagen Digital Insights). This tool was used for trimming of sequencing reads, counting, annotation of the results using miRBase V.21 and differential expression analysis.
In vivo LPS-induced lung injury modelAll animal experiments were approved by Animal Welfare Ethical Review Body of Queen’s University Belfast, in accordance with UK Animals (Scientific Procedures) Act 1986. C57BL/6 male mice (8–12 w.o., Envigo RMS (UK) Station Road Blackthorn Bicester Oxon) were used. Mice were anaesthetised by xylazine/ketamine (0.25 mg/kg and 0.025 mg/kg, respectively) intraperitoneally and LPS was instilled intratracheally (2 mg/kg of body weight), facilitated by a laryngoscope. Four hours after LPS instillation, mice were divided into groups and administered 50 µl of PBS or EVs isolated from 106 of MSCs via tail vein. Mice were euthanised and BALF was taken for analysis 24 hours after LPS administration.
Statistical analysisStatistical analysis was performed using Prism V.7 software (GraphPad, USA). Experiments were done at least in triplicate, the average of three technical replicates was taken as a single data point for each MDM donor, and the points were pooled together for statistical analysis. Data were presented as the mean with SD. Mann-Whitney U test or Kruskal-Wallis test with Dunn’s selected comparisons were used. Statistical significance level was set at p<0.05.
ResultsMSCs limit pro-inflammatory responses of human macrophages via the upregulation of phosphorylated STAT5-SOCS1 signallingHuman MDMs were exposed to LPS with or without the presence of MSC CM for 24 hours. As expected, LPS stimulation resulted in the robust upregulation of tumour necrosis factor (TNF)-α and interleukin (IL)-8 secretion by MDMs, and these responses were significantly ameliorated by MSC CM (figure 1A). At the same time, MSC CM induced significant upregulation of SOCS1 protein expression levels in macrophages in the presence of LPS (figure 1B). To corroborate the role of SOCS1 in macrophage modulation by MSCs, SOCS1 expression in MDMs was silenced by transfection with small interfering (siRNA). siRNA transfection resulted in approximately 60% downregulation of SOCS1 protein expression (figure 1C). Transfection with SOCS1 siRNA but not with scrambled siRNA abrogated the ability of MSC CM to downregulate LPS-induced TNF-α secretion by MDMs, suggesting that SOCS1 is a critical mediator of MSC modulation of macrophages (figure 1D). SOCS proteins act as feedback inhibitors of JAK-STAT signalling and their expression is activated by interaction with phosphorylated STATs (pSTATs). In order to investigate which of the STATs was activated by MSC CM in MDMs in the setting of human ARDS, we co-cultured MDMs with MSCs on Transwell inserts, and exposed them to the samples of BALF from patients with ARDS. After 24 hours of BALF exposure, macrophages were lysed and the cell lysates subjected to a membrane-based phospho-kinase antibody array. Interestingly, it was observed that among four different STATs included in the array, only STAT5a/5b phosphorylation was increased in macrophages by MSCs in the presence of ARDS BALF. Notably, STAT2, STAT3 and STAT6 were not activated by MSCs (figure 1E). These results were further confirmed by Western blotting for STAT3, STAT5 and STAT6. We also probed for STAT1 phosphorylation, which was not included in the array, and found that pSTAT1 was also downregulated by MSCs (figure 1F). Furthermore, stimulation of MDMs with LPS in the presence of MSC CM led to significant pSTAT5 upregulation (figure 1G).
 Figure 1
Figure 1 Upregulation of SOCS1 protein expression in human macrophages is critical for the paracrine effect of MSCs. SOCS1 upregulation is accompanied by the activation of STAT5 phosphorylation. (A) Levels of TNF-α and IL-8 in MDM conditioned medium after 24 hours of exposure to LPS (measured by ELISA) (n=4–7). (B) Immunoblot for protein expression levels of SOCS1 and β-actin in human MDM lysates after stimulation with LPS for 24 hours. Immunoblots were quantified by densitometry and normalised using β-actin expression (n=5). (C) Immunoblot of human MDM lysates, after MDMs were transfected with SOCS1 or scrambled siRNA and stimulated with LPS for 24 hours. Immunoblots were quantified by densitometry and normalised using β-actin expression (n=6). (D) Levels of TNF-α in the conditioned medium of MDMs transfected with scrambled or SOCS siRNA after LPS stimulation for 24 hours, measured by ELISA (n=4). (E) On the left, schematic image showing the co-culture of human MDMs with MSCs using a Transwell system which involved exposure to patient with ARDS bronchoalveolar lavage fluid (BALF) for 24 hours. On the right, phosphokinase array data from human MDMs cell lysates after exposure to ARDS BALF with and without MSC co-culture. Red arrows showed only STAT5a and STAT5b in MDMs could be upregulated by MSCs (n=1). (F) Immunoblots of different STAT proteins in human MDM cell lysates. (G) Immunoblot of pSTAT5 and STAT5 in human MDM lysates at 24 hours after MSC-CM treatment and LPS stimulation. Immunoblots of pSTAT5 were quantified by densitometry and normalised using total STAT5 protein expression (n=6). Data are represented as mean±SD. Kruskal-Wallis test with post-hoc Dunn’s test (A, B, D, G), Mann-Whitney test (C). ARDS, acute respiratory distress syndrome; CM, conditioned medium; IL, interleukin; LPS, lipopolysaccharide; MDM, monocyte-derived macrophages; MSC, mesenchymal stromal cells; pSTAT, phosphorylated STAT; siRNA, small interfering RNA; SOCS1, suppressor of cytokine signalling 1; STAT, signal transducers and activators of transcription; TNF, tumour necrosis factor.
EVs but not EV-free CM recapitulate effects of complete MSC CM on pSTAT5 and SOCS1 expression in human MDMsThe next step was to identify which fraction of the MSC CM (EVs or soluble mediators) was responsible for the observed effects on macrophage modulation. EVs were isolated from MSC CM by ultracentrifugation as previously described29 and characterised according to guidelines of International Society for Extracellular Vesicles (ISEV) (online supplemental figure 1A-D). EV supernatants after ultracentrifugation were subjected to nanoparticle tracking analysis, which has confirmed their depletion from EVs (online supplemental figure 1E). Macrophages were stimulated with LPS and co-cultured with MSC CM, EVs or EV-free supernatants for 24 hours. Interestingly, EVs and MSC CM were comparably effective in downregulation of LPS-induced TNF-α and IL-8 secretion levels (figure 2A) and also in stimulation of pSTAT5 and SOCS1 activation in MDMs (figure 2B), while EV-free CM had no effect. Thus, we decided to further focus on MSC EVs as the active component of MSC CM. To further investigate the role of STAT5 signalling in MDMs, AC-4–130, a specific pharmacological STAT5 inhibitor was used.33 AC-4–130 treatment of MDMs abolished the effect of MSC EVs on SOCS1 upregulation (figure 2C) and pro-inflammatory cytokine secretion (figure 2D).
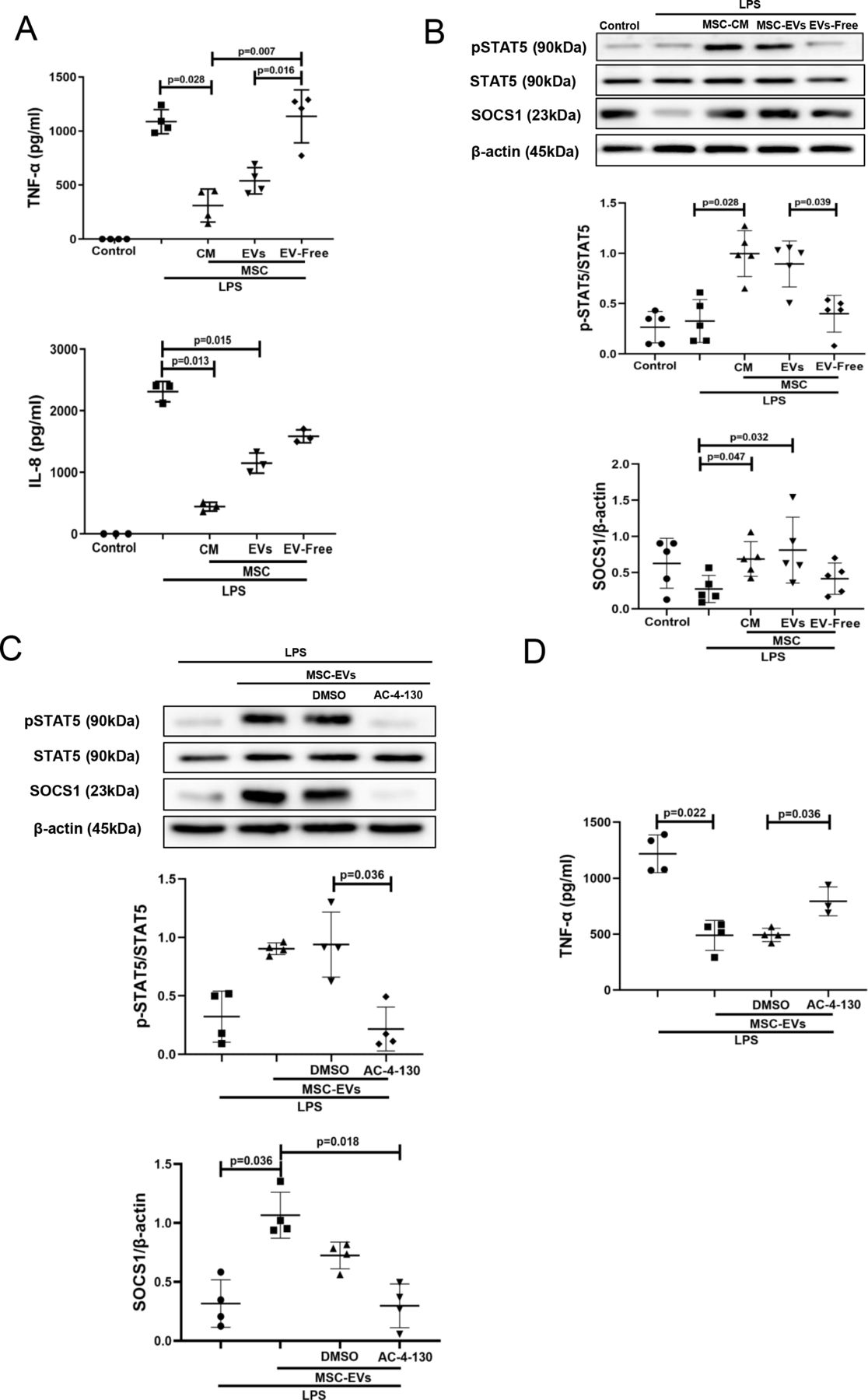 Figure 2
Figure 2 MSC-EVs but not EV-free MSC conditioned medium are responsible for the inhibition of LPS-induced cytokine secretion and upregulation of pSTAT5 and SOCS1 expression in human MDMs. STAT5 phosphorylation is critical for SOCS1 upregulation by EVs. (A) Levels of TNF-α and IL-8 in MDM conditioned medium (measured by ELISA) after LPS stimulation for 24 hours (n=3–4). (B) Immunoblot for protein expression levels of pSTAT5, STAT5, SOCS1 and β-actin in human MDMs lysates after stimulation with LPS for 24 hours. Immunoblots were quantified by densitometry and normalised using STAT5 protein expression for pSTAT5 or β-actin for SOCS1 (n=5). (C) Immunoblot of pSTAT5, STAT5, SOCS1 and β-actin in MDM lysates after MDMs were pre-treated with pharmacological STAT5 inhibitor AC-4–130. Immunoblots were quantified by densitometry and normalised using total STAT5 expression levels for pSTAT5 or β-actin expression levels for SOCS1 (n=4). (D) Levels of TNF-α in MDM conditioned medium after pre-treatment with STAT5 inhibitor and exposure to LPS for 24 hours (n=3–4). Data are represented as mean±SD. Kruskal-Wallis test with post-hoc Dunn’s test (A, B, C, D). CM, conditioned medium; DMSO, dimethyl sulfoxide; IL, interleukin; EVs, extracellular vesicles; IL, interleukin; LPS, lipopolysaccharide; MDM, monocyte-derived macrophages; MSC, mesenchymal stromal cells; pSTAT, phosphorylated STAT; SOCS1, suppressor of cytokine signalling 1; STAT, signal transducers and activators of transcription; TNF, tumour necrosis factor.
MSC EVs modulate pro-inflammatory cytokine secretion and pSTAT5-SOCS1 signalling in MDMs in the presence of plasma from patients with ARDSTo test if this mechanism of macrophage modulation is relevant in the human ARDS environment, and to investigate EV effects in the different ARDS phenotypes, MDMs were cultured for 24 hours in the presence of plasma samples of patients with ARDS which were previously classified into hypo-inflammatory and hyper-inflammatory phenotypes.7 In this model, macrophage exposure to both types of ARDS plasma elicited robust upregulation of TNF-α and IL-8 secretion, and MSC EVs were capable of significant alleviation of pro-inflammatory cytokine production in the presence of both types of ARDS plasma (figure 3A). This effect was coupled with EV activation of the pSTAT5-SOCS1 signalling pathway in both ARDS environments, reaching statistical significance in the presence of hyper-inflammatory plasma (figure 3B).
 Figure 3
Figure 3 MSC EVs regulate pro-inflammatory cytokine secretion and pSTAT5 and SOCS1 expression in MDMs exposed to plasma samples from patients with ARDS in vitro. (A) Levels of TNF-α and IL-8 in MDMs supernatants (measured by ELISA) after stimulation with pooled healthy or hypo-inflammatory or hyper-inflammatory ARDS plasma for 24 hours (n=3–5). (B) Immunoblot of pSTAT5, STAT5, SOCS1 and β-actin protein expression in human MDM lysates after MDM exposure to healthy or ARDS plasma for 24 hours (n=4). Immunoblots were quantified by densitometry and normalised using total STAT5 expression for pSTAT5 or β-actin expression for SOCS1. Data are represented as mean±SD. Kruskal-Wallis test with post-hoc Dunn’s test (A, B). ARDS, acute respiratory distress syndrome; CM, conditioned medium; IL, interleukin; EV, extracellular vesicles; IL, interleukin; MDM, monocyte-derived macrophages; MSC, mesenchymal stromal cells; pSTAT, phosphorylated STAT; SOCS1, suppressor of cytokine signalling 1; STAT, 5, signal transducers and activators of transcription 5; TNF, tumour necrosis factor.
The effect of MSC EVs on MDM modulation is mediated by transfer of miR-181a which regulates phosphatase and tensin homolog-pSTAT5-SOCS1 axisTo explore the miRNA contents of MSC EVs produced by MSCs in the ARDS environment, MSCs were stimulated with ARDS BALF for 24 hours, EVs isolated and subjected to small RNA sequencing. Analysis revealed that EVs express up to 284 known miRNAs as identified in miRBase V.21. Of those, 20 miRNAs were found to be significantly enriched in EVs compared with their parent MSCs, suggesting that these are selectively incorporated into EVs during EV maturation in the ARDS environment. Notably, miR-181a was among the most highly expressed miRNAs in EVs compared with parent MSCs and BALF (figure 4A), its expression in EVs was further confirmed by RT-PCR (data not shown). We were particularly intrigued by miR-181a because one of its well-established target genes is phosphatase and tensin homolog (PTEN),34 which, among other important regulatory roles, negatively regulates STAT5 signalling.35 36
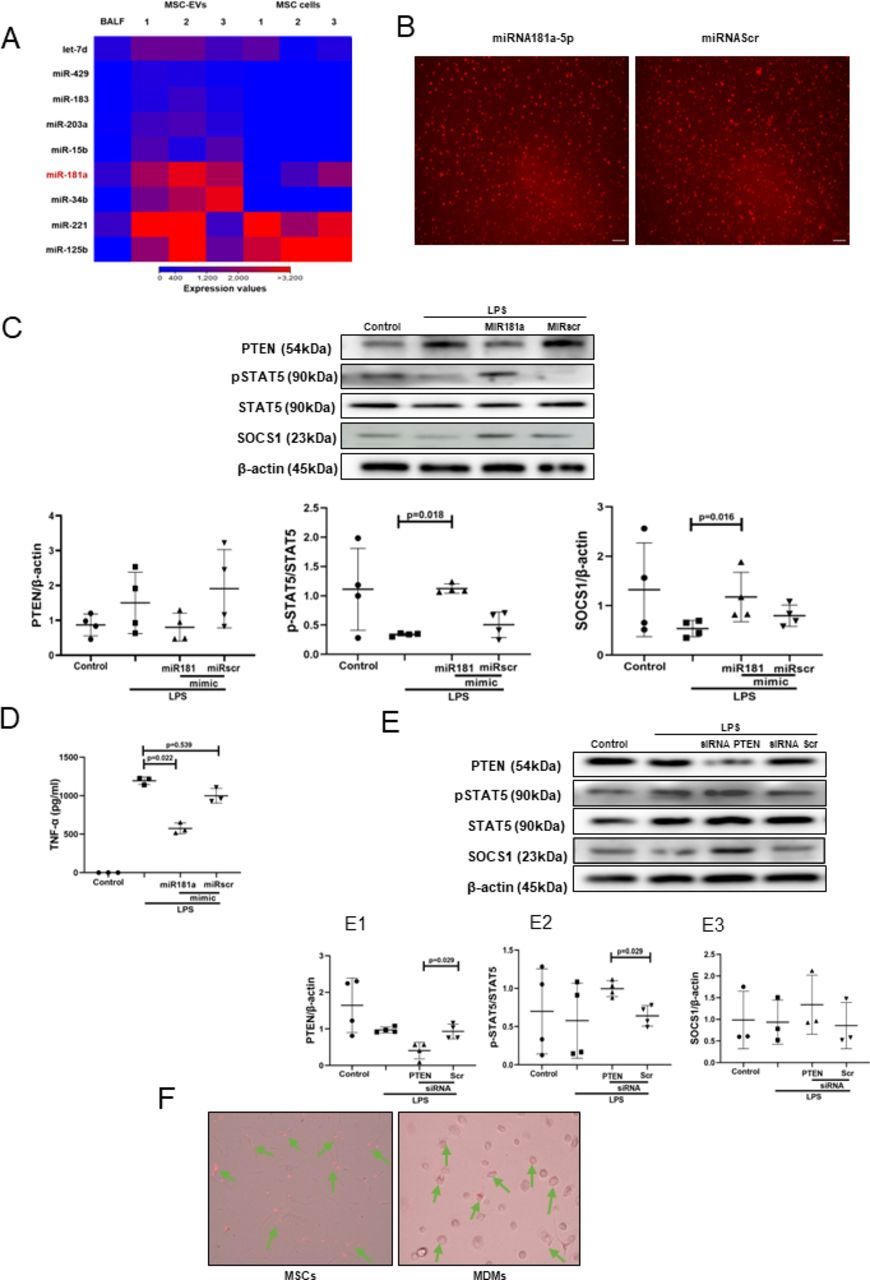 Figure 4
Figure 4 Transfer of miRNA-181a-5p in MSC EVs modulates LPS-induced secretion of pro-inflammatory cytokines through PTEN-pSTAT5-SOCS1 axis. (A) Heat map of next generation sequencing data comparing BALF, cell and MSC-EV expression of selected micro RNAs. ‘1’, ‘2’, ‘3’ labelling refers to different MSC donors. (B) Representative live microscopy images of human MDMs transfection with Dy574-labelled miRNA181a-5p mimic (left) and miRNA Scramble/Negative control mimic (right) as an indicator of efficiency of transfection, monitored by immunofluorescence. Images were taken using EVOS FL Auto epifluorescent microscope (Objective lens 10×, scale bar=50 µm). (C) Immunoblot of PTEN, pSTAT5, STAT5, SOCS1 and β-actin in human MDMs lysates transfected with miRNA181a-5p mimic and miRNA Scramble (negative control) mimic after LPS stimulation for 24 hours. Immunoblots were quantified by densitometry and normalised using total STAT5 expression for pSTAT5 or β-actin expression for PTEN/SOCS1 (n=4). (D) Levels of TNF-α secretion by MDMs after stimulation with LPS for 24 hours (n=3). (E) Immunoblot of PTEN, pSTAT5, STAT5, SOCS1 and β-actin in human MDMs lysates after MDMs were stimulated with LPS for 24 hours. Immunoblots were quantified by densitometry and normalised using total STAT5 expression for pSTAT5 or β-actin expression for PTEN/SOCS1 (n=3–4). (F) Representative live microscopy images of MSCs (left) and human MDMs (right) transfection with Dy574-labelled miRNA181a-5p mimic. The images were taken using EVOS FL Auto epifluorescent microscope (Objective lens 40×, scale bar=50 µm). Data are represented as mean±SD. Kruskal-Wallis test with post-hoc Dunn’s test (C, D), Mann-Whitney test (E). EV, extracellular vesicles; LPS, lipopolysaccharide; MDM, monocyte-derived macrophages; miRNA, micro RNA; MSC, mesenchymal stromal cells; pSTAT, phosphorylated STAT; PTEN, phosphatase and tensin homolog; siRNA, small interfering RNA; SOCS1, suppressor of cytokine signalling 1; STAT, 5, signal transducers and activators of transcription 5; TNF, tumour necrosis factor.
To investigate whether miR-181a alone can modulate MDMs, we transfected MDMs with Dy574-labelled miR181a-5p mimic (efficiency of transfection was monitored by immunofluorescence) (figure 4B). In the presence of LPS, overexpression of miR-181a in MDMs resulted in downregulation of PTEN and significant upregulation of pSTAT5 and SOCS1 protein expression levels (figure 4C) coupled with significantly decreased TNF-α production by macrophages (figure 4D).
To confirm the critical role of PTEN for the activation of pSTAT5-SOCS1 signalling pathway in MDMs, PTEN expression in MDMs was silenced using siRNA. Consistent with the above results, PTEN silencing resulted in the upregulation of pSTAT5 and SOCS1 protein levels in the presence of LPS (figure 4E). To further confirm that miR-181a could be transferred to MDMs via MSC EVs, MSCs were transfected with fluorescently labelled Dy574-miR181a-5p, EVs isolated and applied to MDM cultures. miRNA uptake was visualised by EVOS fluorescent microscopy (figure 4F).
miR-181a-5p-PTEN-pSTAT5-SOCS1 pathway is responsible for MSC EV modulation of MDMs in the presence of ARDS plasmaSpecific locked nucleic acid (LNA) 181a inhibitor was used to silence miR-181a-5p expression in MSCs.16 EVs isolated from MSCs transfected with LNA181a were unable to downregulate expression of PTEN, activate phosphorylation of STAT5 or upregulate SOCS1 expression, compared with EVs isolated from untransfected MSCs or MSC transfected with scrambled LNA inhibitor (figure 5A). Consistently, knockdown of miR-181a expression abrogated ability of the EVs to downregulate LPS-induced secretion of TNF-α by MDMs (figure 5B).
 Figure 5
Figure 5 miRNA-181a transfer through MSC EVs negates the pro-inflammatory response in human MDMs when stimulated with LPS or hyper-inflammatory ARDS plasma. (A) Immunoblot of PTEN, pSTAT5, STAT5 and β-actin in human MDM lysates after stimulation with LPS for 24 hours. Immunoblots were quantified by densitometry and normalised using total STAT5 expression for pSTAT5 or β-actin expression for PTEN/SOCS1 (n=4). (B) Levels of TNF-α secretion by MDMs after stimulation with LPS for 24 hours (n=3–4). (C) Levels of TNF-α secretion by MDMs after stimulation with healthy or pooled hypo-inflammatory or hyper-inflammatory ARDS plasma for 24 hours (n=3–6). (D) Immunoblot of PTEN, pSTAT5, STAT5, SOCS1 and β-actin in human MDMs lysates after stimulation with healthy or hypo-inflammatory or hyper-inflammatory ARDS plasma for 24 hours. Immunoblots were quantified by densitometry and normalised using total STAT5 expression for pSTAT5 or β-actin expression for PTEN/SOCS1 (bottom panel, n=2). Data are represented as mean±SD. Kruskal-Wallis test with post-hoc Dunn’s test (A, B, C). ARDS, acute respiratory distress syndrome; EVs, extracellular vesicles; LNA, locked nucleic acid; LPS, lipopolysaccharide; MDM, monocyte-derived macrophages; miRNA, micro RNA; MSCs, mesenchymal stromal cells; pSTAT, phosphorylated STAT; PTEN, phosphatase and tensin homolog; SOCS1, suppressor of cytokine signalling 1; STAT, signal transducers and activators of transcription ;TNF, tumour necrosis factor.
To further investigate if miR-181a-5p transfer in MSC EVs is relevant for MDM modulation in the human ARDS environment, MDMs were exposed to plasma from patients with ARDS for 24 hours (as in figure 3) and co-cultured with EVs, LNA181a EVs or scrambled LNA EVs. Interestingly, silencing of miR-181a in EVs (LNA181a EVs) significantly abrogated inhibitory effect of EVs on MDM TNF-α production only in the presence of hyper-inflammatory plasma (figure 5C). Activation of the PTEN-pSTAT-SOCS1 signalling pathway by EVs was comparably negatively affected in the LNA181a group in both types of ARDS plasma (figure 5D)
Transfer of miR181a-5p in MSC EVs is critical for the EV immunomodulatory effect in vivoTo induce lung injury, C57BL/6 mice were instilled with LPS intratracheally and 4 hours later treated with PBS, EVs, LNA181a EVs or scramble LNA EVs via tail vein. LPS instillation resulted in significant lung injury and inflammatory cell infiltration into alveolar spaces as demonstrated by increased protein levels in the BALF (figure 6A), BALF total inflammatory cell and absolute neutrophil counts (figure 6B,C) as well as increased levels of inflammatory cytokines TNF-α and keratinocyte (KC) (figure 6D). While control EVs and LNAscr EVs were able to comparably reverse these effects, LNA181a EVs were not effective.
 Figure 6
Figure 6 Transfer of miR181a is critical for the immunomodulatory effects of MSC-EVs in vivo. (A) Total protein concentrations in the BALF samples 24 hours after LPS administration (n=5–7 mice per group). (B) Total leucocyte counts (left graph) and neutrophil cell counts (right graph) in the BALF samples (total cells (n=5–7 mice per group); neutrophils (n=2–4 mice per group)). (C) Representative images of BALF cytospin preparations demonstrating cell recruitment to the airspaces 24 hours after LPS administration. Images were taken using Leica Epifluorescence DM5500 microscope (Objective lens ×20). (D) BALF levels of TNF-α and keratinocyte-derived chemokine (KC, murine analogue of interleukin-8) (n=5–7 mice per group). (E) Representative confocal microscopy of alveolar macrophages, isolated from BALF 24 hours after LPS administration and stained with anti-CD68 (alveolar macrophage marker) and anti-pSTAT5 Ab. Arrows indicate co-localisation of CD68 and pSTAT5. The images were taken using Leica SP8 confocal microscope with a 100× oil-immersion objective (n=3, scale bar=50 µm). Quantitative fluorescence intensity was analysed by Image J software (MFI-pSTAT5). (F) Representative confocal microscopy of alveolar macrophages, isolated from BALF samples 24 hours after LPS administration and stained with anti-CD68 and anti-SOCS1 Ab. Arrows indicate co-localisation of CD68 and SOCS1. Images were taken using Leica SP8 confocal microscope with a 100× oil-immersion objective (scale bar=50 µm). Quantitative fluorescence intensity was analyses by Image J software (MFI-SOCS1). Data are represented as mean±SD. Kruskal-Wallis test with post hoc Dunn’s test (A, B, D, E, F). Ab, antibody; BALF, bronchoalveolar lavage fluid; DAPI, 4',6-diamidino-2-phenylindole; EVs, extracellular vesicles; LNA, locked nucleic acid; LPS, lipopolysaccharide; MFI, mean fluorescent intensity; MSCs, mesenchymal stromal cells; PBS, phosphate-buffered saline; pSTAT, phosphorylated STAT; SOCS1, suppressor of cytokine signalling 1; STAT, signal transducers and activators of transcription ;TNF, tumour necrosis factor.
To investigate if EVs were able to activate pSTAT5-SOCS1 signalling in vivo, in the same experiments, murine alveolar macrophages (AMs) were isolated from BALF at 20 hours after EV treatment, seeded onto the chamber slides (Thermo Fisher), fixed and expression levels of pSTAT5 and SOCS1 in AMs was investigated by immunofluorescence. Consistently with the in vitro results, LPS injury resulted in downregulation of pSTAT5 and SOCS1 expression levels in AMs, which was restored by control EVs and LNAscr EVs, however the restoration was not evident in animals which received LNA181a EVs (figure 6E and F).
Overexpression of miRNA181a-5p in MSCs enhances the therapeutic efficacy of MSC EVs in vivoMSCs were transfected with miR-181a mimic (or scrambled mimic), EVs isolated and administered as a treatment to LPS-injured mice as above. EVs isolated from miR-181a overexpressing MSCs demonstrated significantly stronger ability to reduce lung injury compared with miRscr EVs as indicated by protein levels in the BALF (figure 7A). In addition, miRNA181a-overexpressing EVs had a significantly stronger effect on the reduction of inflammatory cell infiltration (figure 7B and C) and BALF levels of TNF-α and KC cytokines (figure 7D) compared with miRscr EVs.
留言 (0)