Structural and functional analysis of cytochrome P450 (CYP or P450) genes and proteins in an environmental context has provided critical information on the roles, regulation, and evolution of the P450 superfamily [1]. For example, comparisons of expression and induction of CYP1 genes in fish inhabiting polluted and clean sites have pointed to a pollution-driven evolutionary adaptation in the aryl hydrocarbon (AH) receptor pathway [2]. P450 adaptations to environmental conditions include gene duplication (e.g. in pesticide resistance [3]), as well as single nucleotide substitutions, as found in the sexual selection of CYP2J19 to produce red carotenoids in birds [4]. Other environmental selection pressures may lead to sequence changes that result in functional adaptation to the stressor. This paper concerns the structural adaptation in P450s to an extreme environment, the deep sea, and the possible impact of environmental chemicals known to occur in the deep ocean.
Molecular adaptation to life in the deep sea is essentially an adaptation to maintain functional control at high hydrostatic pressures [5]. Pressures in the ocean increase by approximately 1 atmosphere (atm) (0.1 MPa) per 10 m of depth. At 4000 m, the average depth of the North Atlantic Ocean, the pressure is roughly 400 atm (40 MPa). The pressures in the deep ocean are high enough to necessitate evolution of P450 protein structural modifications to account for pressure influence(s) on volumes associated with ordered water molecules in channels and around amino acid side-chains, as well as volume changes during catalysis, to continue enzyme function in a controlled way [6,7]. Increased intracellular concentrations of osmoregulatory compounds such as trimethylamine oxide compensate for some water effects, but typically such molecules cannot penetrate into protein water channels [8]. We previously described positionally unique amino acid residues in the CYP1A protein from a deep-sea fish, Coryphanoides armatus [9], which occurs in the ocean worldwide, at depths between 2000 and 5000 m [10]. C. armatus, also known as the abyssal grenadier, typically grows to 40 cm, may live to 30 years or more, and maintains low metabolic rates during foraging [11]. The amino acid changes observed in CYP1A were ones that were on the surface of the protein rather than in access channels or the active site, and which we predict reduces the overall ordering of water molecules around the protein [9].
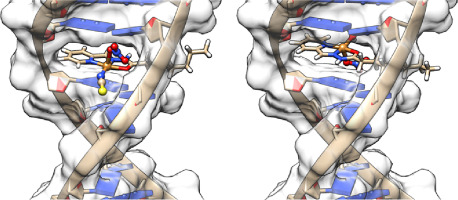
In the present study, we have now examined structural models of an essential P450, CYP51, also from C. armatus. In many animals CYP51 catalyzes a critical step during cholesterol biosynthesis, the stepwise oxidative demethylation of lanosterol [12] (Fig. 1). The objective for this study was two-fold. First, we hypothesize that, like CYP1A, CYP51 will show positionally unique amino acid residues on the surface of the protein. Second, we are interested in CYP51 as a possible target for pollutant chemicals that occur throughout the world and which penetrate even the abyssal and hadal depths of the oceans [[13], [14], [15], [16]]. The apparent induction of CYP1A protein in deep sea fish, including C. armatus, is associated with polychlorinated biphenyl (PCB) pollutants in the deep ocean and in tissues of the fish [15,17]. Given the essential role of CYP51 in sterol biosynthesis, we are interested in this protein as a possible target for azoles or other inhibitors of CYP51 that can occur in the ocean. Plastic particles are now ubiquitous in the oceans including in the abyssal realms [[18], [19], [20]], and are being found in deep sea fish [21]. UV-protectants contained in plastic particles include compounds such as hydroxybenzophenones and benzotriazoles, that potentially can bind to CYP51 proteins. Indeed, 4,4′ dihydroxybenzophenone was reported to bind to CYP51 from Mycobacterium tuberculosis [22] (Fig. 1). Such chemicals were examined for binding using recombinant C. armatus CYP51. In addition to changes in amino acid residues on the surface of the protein, computed protein structures of the deep sea CYP51 and other CYP51s were examined for architectural differences, which identified here-to-fore unknown supplemental cavities in human and other CYP51 proteins.




留言 (0)