
記住我
Melanoma is the deadliest form of skin cancer worldwide, accounting for 3% of these cancers but 65% of their mortality.1 The latter is largely attributed to metastases development,2 late diagnosis3 and resistance to therapies.4 Due to the high immunogenicity of melanoma,5 immunotherapies became a promising biopharmaceutical class. The recent use of immune checkpoint inhibitors (ICI) such as nivolumab (anti-programmed cell death protein-1 (PD-1)), pembrolizumab (anti-PD-1) and ipilimumab (anti-cytotoxix T-lymphocyte antigen 4 (CTLA-4)) increased the survival of patients with melanoma.6 7 However, if diagnosed lately, many cases cannot be cured by the array of current therapeutic approaches. Disappointing clinical outcomes have been linked to the inability of therapies to sustainably focus effect on cancerous tissue throughout the body, with patients non-responding to these therapies,8 severe adverse events8 9 and the development of resistance.10 The use of live microorganisms as immunotherapies may be the most promising approach to treating cancer that overcomes current therapeutic limits. To date, only one live microorganism is approved by the Food and Drug Administration and European Medicines Agency for the treatment of unresectable advanced melanoma, IMLYGIC, a type 1 herpesvirus modified to secrete Granulocyte-Macrophage Colony-Stimulating Factor (GM-CSF).11 The use of IMLYGIC led to an improved durable response rate and overall survival with minor adverse events, but despite these impressive results, approximately half of the patients does not respond to this therapy.12 Thus, there is still room for improvement to overcome issues raised by current immunotherapies.
While prior research has mainly focused on lower order microorganisms, viruses and bacteria, our group has determined high anticancer properties of a protozoan. In a previous article, we reported that Neospora caninum, an obligate intracellular protozoan non-pathogenic for humans, inhibited the development of murine EG7 thymoma when administered intratumorally or distally from the tumor site.13 The antitumoral properties of N. caninum rely on its direct oncolytic activity by intracellular multiplication within cancer cells, the stimulation of the immune system with the induction of an effective T helper 1 immune response and reprogramming of the immunosuppressive tumor microenvironment (TME). Following destruction of tumor, all traces of this protozoan rapidly become undetectable in blood or in biopsied organs from test subjects. N. caninum derived immunotherapies would have a singular safety characteristic thanks to its lack of reported pathogenicity in humans. We have shown the ability of N. caninum to vectorize molecules of therapeutic interest, by engineering a viable strain that secretes human interleukin-15 (IL-15). In several studies, the use of IL-15, combined or not, led to impaired tumor and metastases growth with enhanced survival through the recruitment of natural killer (NK) cells and CD8+ cells.14 15 As IL-15 is more stable and efficient when bound to its high affinity receptor IL-15Rα, receptor-linker-IL-15 has been developed,16 mimicking IL-15 transpresentation.17 Indeed, several studies have shown that the dominant mode of action of IL-15 in vivo is through a trans-presentation mode in which IL-15 bound to IL-15 Rα (expressed by antigen-presenting cells) activates the IL-15Rβ/γ complex expressed by neighboring naïve T or NK cells.18 19 This activation leads to T-cell proliferation, permits activation of T cells that otherwise would have remained tolerant, enhances CD8+ T-cell survival, enhances T-cell trafficking through induction of chemokines and chemokines receptor expression and induces effector responses in CD8+ T cells and NK cells.20 The engineered NC1-IL-15hRec strain exhibits enhanced antitumoral properties over the wild-type strain.
By reason of N. caninum effect on tumor at distance from the point of administration, we postulate that N. caninum immunotherapies could be effective against metastases. This article reports on an application against metastasis in a model that treats B16F10 pulmonary metastatic melanoma in syngeneic mice via intranasal and intravenous routes of administration. The B16F10 lung metastasis model is a highly reproductible and reliable model to study the efficacy of treatment for lung metastases,21–23 with numerous relevance towards clinic. This has led to widespread use of this model in preclinical studies to evaluate various treatments for lung metastases, including immunotherapies.24 25 B16F10 tumors are preferentially targeted by NK cells that exert robust control on metastatic dissemination which appears to be an ideal feature to study IL-15 efficacy on boosting NK cells antitumor effects. Here, it highlights the potential of using IL-15-secreting N. caninum in controlling development of metastases. Beneficial effect was observed with mucosal administration of the IL-15-secreting N. caninum, associated with a recruitment and activation of NK cells, CD8+T cells and macrophages polarized towards an M1 phenotype, the secretion of interferon (IFN)-γ, IL-12 and tumor necrosis factor (TNF)-α and the activation of the CCR5 and CXCR3 axes.
Materials and methodsMiceFemale 8 weeks aged C57Bl/6 were purchased from CER Janvier Labs (Le Genest Saint Isle, France) and maintained under pathogen-free conditions in the animal facility of the UMR ISP 1282. All the experiments were approved by the MESRI (Ministry of higher education, research and innovation) with the procedures 2018032615069312, 2020071016522665 and 2021052818381834 in accordance with the guidelines for animal experimentation (European Union directive 2010/63/EU).
CellsThe murine melanoma cell line B16F10 was obtained from American Type Culture Collection (CRL-6475) and the luciferase and mCherry-expressing B16F10 cell line was kindly provided by Dr Mehdi Khaled (UMR 1299, Gustave Roussy Institute, Paris-Saclay University, France). Tumor cells were cultured in Roswell Park Memorial Institute medium (RPMI, Dutscher) supplemented with 10% of fetal calf serum (FCS, PAN-Biotech) and 1% HEPES (N-(2-Hydroxyethyl)piperazine-N′-(2-ethanesulfonic acid), Dutscher) under an atmosphere containing 5% CO2. An extract of B16F10 cells was obtained through three rounds of sonication at 60 W for 60 s. The resulting suspension was centrifuged at 2000 g for 15 min at 4°C and the protein concentration was determined using bicinchoninic acid (B-9643, Sigma) and cupric sulfate (C-2224, Sigma) and bovine serum albumin (BSA) as the protein standard. Extracts were then stored at −20°C.
ProtozoaN. caninum (NC1 strain obtained from ATCC (50843), β-galactosidase expressing NC1 kindly provided by Andrew Hemphill (University of Bern) and TdTomato NC1 and NC1-IL15hRec13 were processed as previously described13 (see details in online supplemental methods).
Experimental designB16F10 cells (105 in 100 µL of phosphate buffered saline (PBS) or 5×105 in 100 µL of PBS as described in respective figure legends) were intravenously injected in mice caudal vein. Mice were then inoculated, subcutaneously, intravenously or intranasally with freshly isolated tachyzoites in PBS as indicated in the legend of corresponding figures. Blood sera were collected via cheek bleed from mice at the indicated time points. Before harvesting of lungs, bronchoalveolar lavages were performed using 1 mL of PBS. A first lung lobe was collected in 4% paraformaldehyde (PAF) for immunofluorescence assays. A second lung lobe was collected and stored at −80°C for quantitative PCR analysis and transcriptomic study. The rest of the lung was dissociated using mouse tumor dissociation kit and gentleMACS Dissociator (Miltenyi Biotec). Cells were resuspended in PBS, 2 mM EDTA for staining and flow cytometry analysis. Spleens and mediastinal lymph nodes were harvested then mechanically dissociated to obtain cell suspensions. Cells were plated in RPMI (5% FCS, 1% HEPES, 1% glutamine (PAN-Biotech), 1% sodium-pyruvate (PAN-Biotech), 100 U/mL penicillin/streptomycin (PAN-Biotech) for in vitro stimulation with N. caninum extract at 10 µg/mL, B16F10 cells extract at 10 µg/mL or concanavalin A at 5 µg/mL.
Tumor size measurementsMacroscopic melanoma scoring of the lung lobes from individual animals were further quantified with Fiji software using an ImageJ macro that we developed (Fiji/ImageJ,26 macro provided in online supplemental data).
Cellular analysisFlow cytometry analysis was performed on cell suspension after tumor dissociation using antibody reagents detailed in online supplemental methods and gating strategies are disclosed in online supplemental figure 2. Flow cytometry analysis was performed using a Miltenyi 8-color MACSQuant and data were analyzed using FlowLogic (Miltenyi Biotec).
Cytokine detectionThe cytokine levels in splenocyte and lung dissociation supernatants were evaluated by multiplex dosage using MACSPlex cytokine 10 kit (Miltenyi Biotec). IFN-γ serum level was assessed by ELISA using mouse IFN-γ uncoated ELISA kit (Invitrogen).
Transcriptomic studyRNA isolation was performed on a lung lobe using TRIzol reagent (Invitrogen) protocol. RNA integrity was then checked by running them on agarose gel stained with ethidium bromide. Complementary DNA samples were obtained using RT² First Strand Kit (Qiagen). Transcriptomic study was then performed using RT² Profiler PCR Array Mouse Cancer Inflammation and Immunity Crosstalk (Qiagen) with 500 ng RNA from each sample.
Immunofluorescence assaysLung lobes fixed for 24 hours in 4% PAF were incubated overnight in 30% sucrose. Lung tissues were snap frozen in Optimal Cutting Temperature compound then cut in 8 µm cryosections. Those cryosections were incubated for 10 min in PBS 0.1% Triton X-100 and blocked for 1 hour in PBS 2% BSA, washed with PBS 0.1% Tween 20 and 1% BSA and incubated overnight at 4°C with Alexa-488-conjugated anti-CD68 (clone FA-11, eBioscience). Cell nuclei were stained using Hoechst. N. caninum tachyzoites staining was performed using rabbit infection serum as primary antibodies incubated overnight at 4°C and Alexa 488 Goat anti-rabbit IgG (Thermo Fisher Scientific) incubated at room temperature for 1 hour. Images were captured with an Olympus IX73 microscope using cellSens Dimension software or EVOS M7000 (Thermo Fisher Scientific).
Neuraminidase treatmentB16F10 cells were desialylated using neuraminidase derived from Arthrobacter ureafaciens (Roche) diluted at 0.2 U/mL. 5×106 B16F10 cells were resuspended in this neuraminidase solution then incubated for 1 hour at 37°C. Desialylation of B16F10 cells was validated by sialic acid staining. B16F10 cells were incubated for 30 min at 4°C with two lectins to stain α2,6 and α2,3-sialic acid: respectively, Sambucus Nigra lectin-FITC (SNA, Invitrogen) and Maackia amurensis lectin-FITC (MAA/MAL I+II, BioWorld) diluted at 1:200. Then, the expression level of sialic acid was determined by flow cytometry using a Miltenyi 8-color MACSQuant and data were analyzed using FlowLogic (Miltenyi Biotec).
Explants culture and spheroids obtentionLymph node dissections of metastatic melanoma from patients from Tours hospital (obtained after patients’ written informed consent) were cut in 30 mg explants and cultured in 200 µL-wells with RPMI supplemented with 6% human serum (Innovative Research), 2% penicillin/streptomycin, 1% fungizone and 50 U/mL IL-2 (Miltenyi Biotec). The explants were infected with 107 tachyzoites of N. caninum, supernatants were collected at 24 hours and 48 hours post-infection (hpi). Forty-eight hours after infection, explants were fixed in 4% formol or 4% PFA for histological analysis and immunofluorescence on the frozen section, respectively. The remaining tissues were mechanically dissociated to obtain cell suspensions that were adjusted at 5000 cells in 10 µL in the same media than explants. Then, 10 µL-drops of the cell suspensions were disposed on the lid of a Petri dish to obtain spheroids using the hanging drops technique27 and incubated 5 days at 37°C under an atmosphere containing 5% CO2. After 5 days, spheroids were infected with 5000 tachyzoites of N. caninum and the spheroids growth was monitored using EVOS M7000 (Thermo Fisher Scientific).
Principal component analysisPrincipal component analysis (PCA) was performed in R studio using the PCA and heatmap function in R.
Statistical analysisStatistical analysis was performed using GraphPad Prism software. P values<0,05 were considered as statistically significant. Error bars show the SEM. Bar graphs were analyzed by parametric one-way analysis of variance.
ResultsN. caninum is associated with therapeutic effect against metastasesThe efficacy of N. caninum on metastasis was studied using a B16F10 lung tumor bearing mouse model. In this model, lung tumors are detectable from D5 post intravenous tumor inoculation.28 29 Seven days post tumor implantation, tumors are therefore established and mice were treated with 2×106 tachyzoites of N. caninum administered either subcutaneously, intranasally or intravenously to assess the most efficient route of administration (figure 1A). We observed that regardless of the administration route, N. caninum significantly impaired tumor growth (figure 1B,C). While lung tumors represented 57% of the total lung surface of control mice, in mice treated subcutaneously, intranasally or intravenously with N. caninum, tumors represented 10%, 6% and 4% of the lung surface, respectively. Although a strong diminution of metastasis sizes was observed in subcutaneously treated mice, 57% of those still harbored lung metastases (80% for control mice). However, only 20% and 10% of mice treated intranasally and intravenously harbored lung metastases, respectively (figure 1B). Those results indicate that the administration route has bearing on the antitumor response induced by N. caninum with a greater beneficial effect following intravenous and mucosal administration.
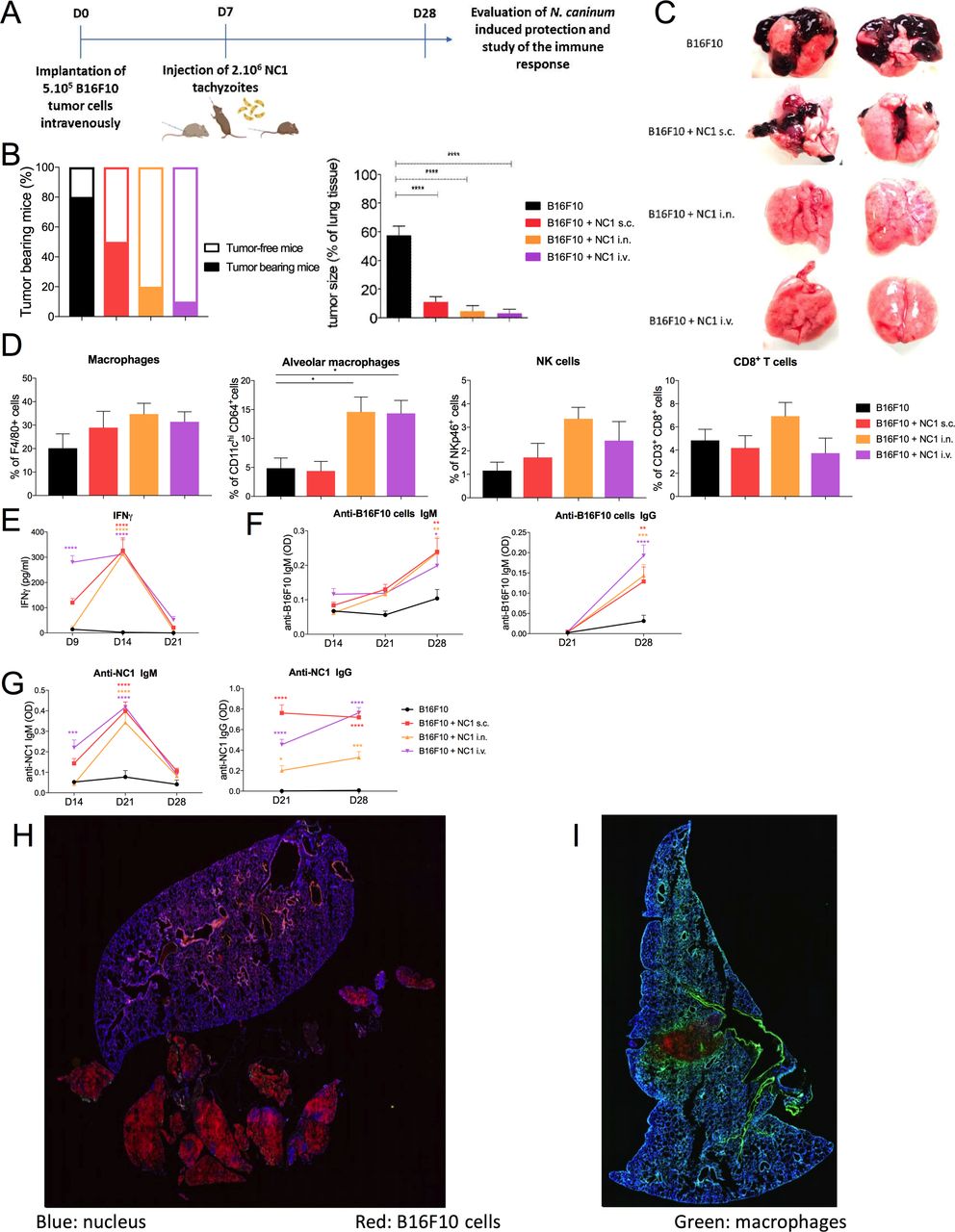 Figure 1
Figure 1 Different routes of administration of Neospora caninum and effect on inhibition of tumor development. B16F10 cells (1×105 in 100 µL of phosphate buffered saline) were inoculated intravenously in C57Bl/6 mice and 2×106 NC1 tachyzoites were injected subcutaneously, intravenously, or intranasally 7 days later (A). Twenty-eight days after tumor cell inoculation, mice were euthanized to determine macroscopic observation of lung metastases implantation (B) and tumor size as a percentage of lung tissue using ImageJ software (C). Lungs were collected and dissociated, tumor cell suspensions were centrifuged and cells were resuspended for staining and flow cytometry analysis (D). Sera were collected at day 9, 14, 21 and 28 after tumor cell injection, and levels of IFN-γ (E), anti-B16F10 cells IgM and IgG (F) and anti-NC1 IgM and IgG (G) were assayed. Sections of lung from control mice (H) and intranasally treated (I) mice were stained by immunofluorescence using an anti-CD68 antibody to reveal macrophages (green) and Hoechst for cell nucleus staining (blue), whereas B16F10 cells are visualized due to mCherry expression (red). n=10 mice per group. *p<0.05; ****p<0.0001. IFN, interferon; i.v., intravenous; i.n., intranasally; NK, natural killer; s.c., subcutaneously.
Interestingly, a similar dichotomy was found while studying the immune response induced by N. caninum treatment. Subcutaneous inoculation of N. caninum did not increase the immune cell number in the lung 21 days post-treatment. However, intravenous and intranasal administration induced a threefold increase in the number of alveolar macrophages and an increasing trend of CD8+ T cells and NK cells for the intranasal route (figure 1D). Immunofluorescence staining of lung cryo-sections revealed a high number of macrophages localized in periphery and within tumor masses after intranasal administration of N. caninum (figure 1I), while in untreated mice, few macrophages were localized only in the periphery of some metastases (figure 1H).
The stimulation of a local antitumor immune response induced by N. caninum was associated with a peak of IFN-γ in the sera of treated mice 7 days post-treatment (figure 1E), and a slight increase of anti-B16F10 (figure 1F) and anti-NC1 antibodies (figure 1G) from 14 to 21 days post-treatment with no significant difference between the intranasal and intravenous routes.
Those results highlight the interest of using N. caninum for its potential application not only in primary tumors but also against metastasis since a single administration by intravenous or intranasal routes led to inhibition of lung metastasis development.
A series of three doses via intranasal administration of IL15-secreting N. caninum completely prevented implantation of metastasesDespite the drastic inhibition of metastases implantation in mice treated with a single administration of N. caninum, small metastases were still harbored in the lungs of treated mice, suggesting the need for improving N. caninum efficacy and/or performing several treatments to fully control the tumor development. To this aim, we engineered an N. caninum strain to secrete human IL-15 associated with the alpha subunit of the IL-15 receptor (NC1-IL15hRec) which showed an increased efficacy in an EG7 thymic lymphoma mice model.13 Therefore, the potential of this engineered strain was assessed in the B16F10 melanoma metastases mouse model. To decipher potential added-value of the IL-15-secreting strain, the number of B16F10 cells injected intravenously was increased by fivefold, making it easier to accurately detect differences in treatment response by inducing a more challenging tumor burden. Due to the therapeutic benefit of the intranasal and intravenous treatments, a focus was made on those two administration routes. We treated B16F10 metastases with three administrations of N. caninum at 7 days, 14 days and 21 days post-tumor cells inoculation (figure 2A). Regardless of the route of administration and of the strain administered, N. caninum treatment significantly inhibited metastasis development (figure 2B,C). NC1 strain administered either intranasally or intravenously partially inhibited metastases growth with metastases implantation representing 36% of the lung surface in untreated mice, versus 14% in intravenously treated mice and 4% in intranasally treated mice (figure 2B). The NC1-IL-15hRec showed an improved efficacy with metastases representing 3% in mice treated intravenously and only 0.08% in mice treated intranasally (figure 2B). Such antitumor protection was associated with a decrease of circulating B16F10 cells in the peripheral blood directly after the first intravenous treatment and after the second intranasal treatment regardless of the strain (figure 2D). Such potentiated antitumoral protection induced by NC1-IL-15hRec administered intranasally improved the mice’s survival with a median survival of 45.5 days with 30% of mice treated alive at 2 months. Median survival of the control group and group treated intranasally with NC1 were 27 and 33, with no surviving mice post-30 and 45 days, respectively (figure 2G).
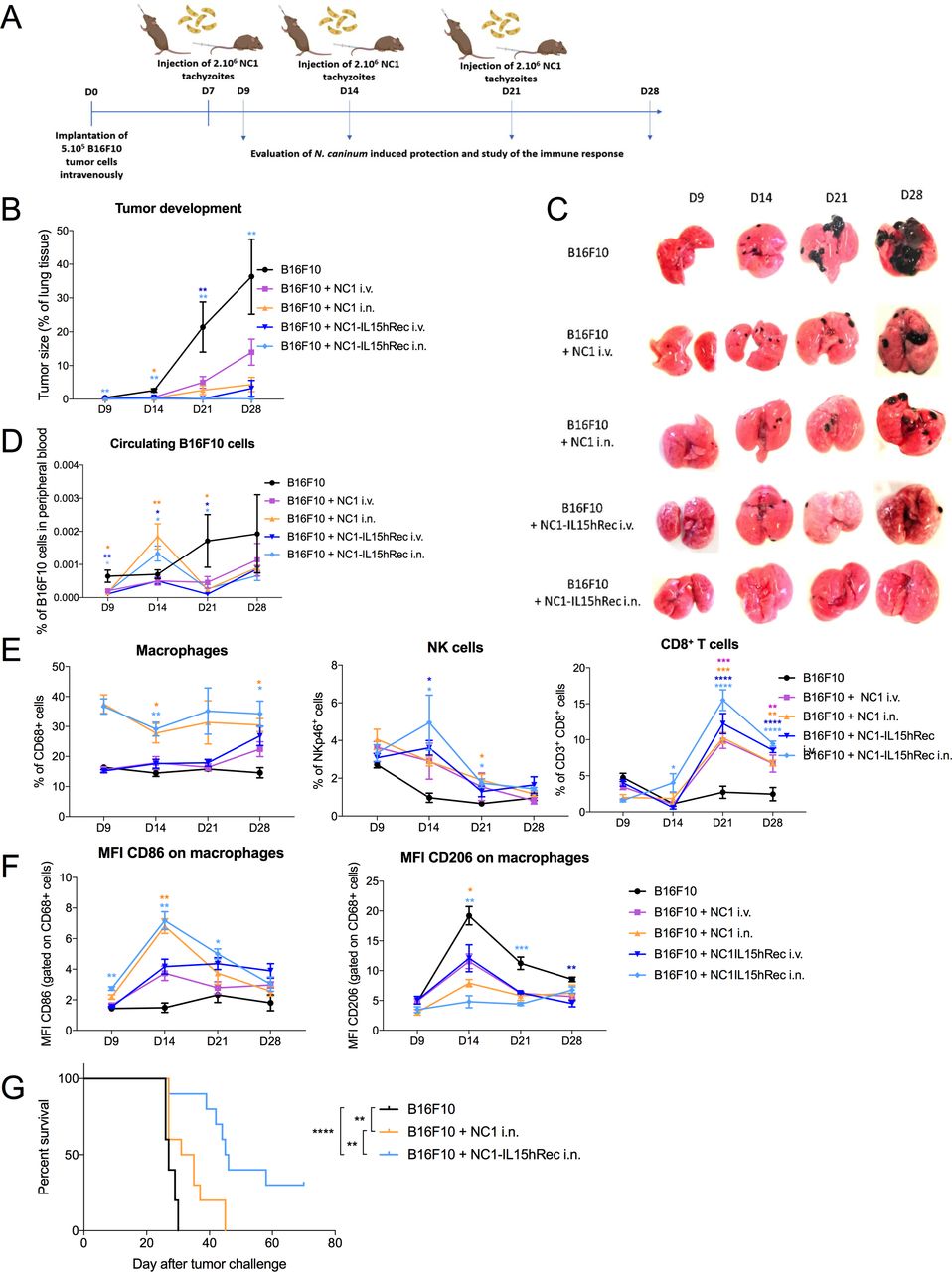 Figure 2
Figure 2 Tumor development inhibition comparing wild-type strain of Neospora caninum (NC1) and IL15-secreting strain of N. caninum (NC1-IL15hRec). B16F10 cells (5×105 in 100 µL of phosphate buffered saline) were inoculated intravenously in C57Bl/6 mice and 2×106 tachyzoites were administered intravenously or intranasally (i.n.) at day 7, day 14 and day 21. Five mice per group were euthanized at day 9, day 14, day 21 and day 28 (A) tumor size in percentage of lung tissues were determined using ImageJ software (B) and by macroscopic assessment of lung metastasis implantation (C). Circulating tumor cells in peripheral blood were assayed by flow cytometry (D) and the percentage of macrophages, NK cells and CD8+ T cells in the lung (E). Macrophage phenotype was characterized by flow cytometry by assaying expression levels of CD86 and CD206 on macrophages (F). n=20 mice per group (five per day). Kaplan-Meier survival curve comparing untreated mice and mice treated intranasally with NC1 or NC1-IL-15hRec. Survival curves were compared, and p values calculated by log-rank test (G), n=10 per lot. *p<0.5; **p<0.01; ***p<0.001; ****p<0.0001. MFI, Mean of Fluorescent Intensity; IL, interleukin; i.v., intravenous; NK, natural killer.
The control of metastases development induced by N. caninum was associated with a strong increase of immune cells within the lung, such as macrophages, NK cells and CD8+ T cells. The NC1-IL15hRec strain induces an enhanced increase of NK cells and CD8+ T cells, respectively, at day 14 and from day 21 (figure 2E). As rapidly as 2 days after N. caninum treatment (day 9), a potent increase of macrophages was observed in the lungs of mice treated intranasally. Whereas this increase of macrophages was constant over time in intranasally treated mice, it was only transiently observed at day 28 in intravenously treated mice. Furthermore, expression of surface markers of macrophages, CD86 for M1 and CD206 for M2, was assessed to determine their phenotype. Interestingly, N. caninum intranasal treatment induced a CD86 expression increase and CD206 decrease in macrophages, revealing a polarization towards an M1 phenotype30 (figure 2F). It is worth mentioning that only a slight increase of CD86 expression associated with a slight decrease of CD206 was observed in intravenously treated mice. These results highlight the benefits of intranasal administrations of NC1-IL-15hRec for the treatment of murine melanoma metastases.
NC1-IL-15hRec treatment through intranasal route induces an enhanced protective immune responsePCA was performed to generate a heat map based on cytometry analysis of multiple immune cells and tumor growth parameters from mice 2 days after a first treatment (day 9) or a week after a second treatment (day 21). The heat map generated with data (online supplemental table 1) at day 9 underlines the added value of the non-invasive intranasal route to induce a swifter antitumoral protection sustained by a strong induction of a local innate immune response. This innate immune response relies on the recruitment of F4/80+ macrophages and CD68+ CD64+ CD11c+ alveolar macrophages with an increased expression of CD86 as well as CD11b+ cells, Ly6C+ cells and NK cells. At that time point, no or only few differences distinguished mice treated by intravenous administration of N. caninum versus untreated mice (figure 3A). At 21 days post-implantation (data in online supplemental table 2), the heat map discloses a shift in the immune response signatures with the indication of an adaptive immune response induction with an increase of CD4+ T and CD8+ T-cell numbers, sustained by an increase of NK cells, DCs, macrophages and alveolar macrophages with an increased expression of CD86 (figure 3B). Furthermore, the heat map reveals the clustering of NC1-IL-15hRec treated mice and displays the highest increased parameters described above at day 21 regardless of the administration route.
 Figure 3
Figure 3 Mechanisms underlying the antitumoral protection induced by NC1 WT and NC1-IL-15hRec. Principal component analysis was performed on flow cytometry and tumor size data either at day 9 (A) or at day 21 post tumor cells inoculations (B). The expression of genes involved in cancer inflammation and immunity crosstalk in the lungs of mice treated intranasally by NC1 and NC1-IL-15hRec was assayed by quantitative real-time PCR. Messenger RNA expression was normalized to the expression of five housekeeping genes to calculate the ΔCt. Both treated groups were compared with the control group to calculate the ΔΔ Ct. Hierarchical clustering of gene expression was performed on median centered 2-ΔΔ Ct value. Higher and lower expressions compared with the median are represented, respectively, in deep to light red or blue (C). Fold-changes superior and inferior at two for both treated groups comparing control are represented and grouped by function: chemokines, chemokine receptors, antitumoral factors, cytokines, regulation of the immune response, protumoral factors (D). Data represent pooled samples (n=5 per group). MFI, Mean of Fluorescent intensity; PMNs, Polymorphonuclear neutrophils; DC, dendritic cell; IL, interleukin; i.n., intranasally; i.v., intravenous; NK, natural killer; Treg, regulatory T cells.
Those results highlight the added value of NC1-IL-15hRec inoculated intranasally to obtain a swift induction of a local innate immune response associated with the induction of a potent adaptive immune response.
To better understand the mechanisms of action of N. caninum treatment and the mechanisms underlying the added value of NC1-IL-15hRec, a transcriptomic study was performed on lung samples from intranasally treated mice at day 21. An upregulation (from 2 to 32-fold) of expression of genes coding for chemokines and their receptors was identified after N. caninum treatment (figure 3C). Those chemokines and receptors are known to play a major role in the recruitment and homing of cellular actors required for an antitumoral immune response, such as CCR5 and its ligands CCL4 and CCL5 involved in the recruitment of DCs, NK cells and T CD8+cells, CXCR3 and its ligands CXCL9 and CXCL10 involved in the recruitment of NK cells, T CD8+cells and Th1 T CD4+cells. The CCR5 and CXCR3 axes, which are overexpressed in treated mice, could drive the recruitment of immune cells observed after N. caninum treatment. In the lungs of N. caninum-treated mice, messenger RNA (mRNA) expression of both granzyme GzmA and GzmB, as well as FasL, Tnf, Ifng, Il12b and Tnfsf10 increase. The proteins coded by those genes are involved in the killing of target cells by CD8+ T cells and NK cells. Further modulation of the immune system was observed with changes in mRNA expression of the immune response modulator. N. caninum treatment induces an overexpression of major histocompatibility complex (MHC) mRNAs, H2-K1 and H2-D1. That result is correlated with an increase of MHC-II expression in dendritic cells (DCs) and macrophages (online supplemental figure 1). Interestingly, the IL-15 secreting strain of Neospora seems to induce lower expression of MHC-II on macrophages and DCs, probably due to the IL-15 capacity to induce IFN-γ secreting phenotype on DCs, characterized by a lower expression of common DC markers such as CD11c and MHC-II.31
While supporting tumor regression, the activation of the immune system also allows Neospora elimination. Although the latter is well described in the literature,32 we aimed to assess if the presence of lung tumor could affect Neospora distribution after intranasal administration. No difference in distribution was found between tumor bearing and healthy mice (online supplemental table 3). Interestingly, mRNAs expression of some protumoral factors as Egf, Nos2, Mif and Myc is downregulated. Thus, downregulation of those factors sustains the antitumoral properties of N. caninum. However, an overexpression of Ctla4, Cd274 and Pdcd1 mRNAs, respectively translated in CTLA-4, programmed death-ligand 1 (PD-L1) and PD-1, was observed in the lungs of N. caninum-treated mice. Those proteins are known to downregulate the immune response and to be protumoral factors.
Potential affinity of N. caninum for cancer cellsN. caninum is an obligate intracellular protozoan, the infection of target cells is mandatory to allow its multiplication, causing direct oncolytic activity of infected tumor cells. To decipher that process, lungs from intranasally treated mice were harvested 3 days post-treatment to ascertain the presence of N. caninum tachyzoites. Parasitophorous vacuoles were observed proving the active N. caninum replication in the lung after intranasal administration (figure 4A). Those vacuoles were also observed in vitro in B16F10 cells, as well as the vacuoles rupture provoking cell lysis and therefore a direct oncolytic effect (figure 4B). Then, N. caninum infectivity of cancer cell was investigated. As cancer cells are mainly hypersialylated,33 we investigated if sialylation could promote N. caninum entry to cancer cells. A β-galactosidase expressing strain of N. caninum (NC1-βgal) was used to assess entry (4 hours) and replication (24 hours) of N. caninum in sialylated versus unsialylated (with neuraminidase treatment) B16F10 cells. Higher levels of β-galactosidase were found in sialylated cells as compared with unsialylated cells at 4 hpi and 24 hpi (figure 4C). However, in both conditions, the β-galactosidase level increases threefold between 4 hpi and 24 hpi, indicating no difference in the multiplication capacity of N. caninum in desialylated and sialylated cells. Thus, N. caninum more efficiently infects sialylated cells than non-sialylated cells, suggesting a role of the sialylation in the preferential targeting by N. caninum of cancer cells over normal cells. However, this result raises the need of further studies to decipher the expression level of sialic acid between normal cells and cancer cells since cells from the respiratory tract are also known to express a high level of sialic acid. Furthermore, many other mechanisms could be involved in the potential affinity of N. caninum for cancer cells and will have to be identified.
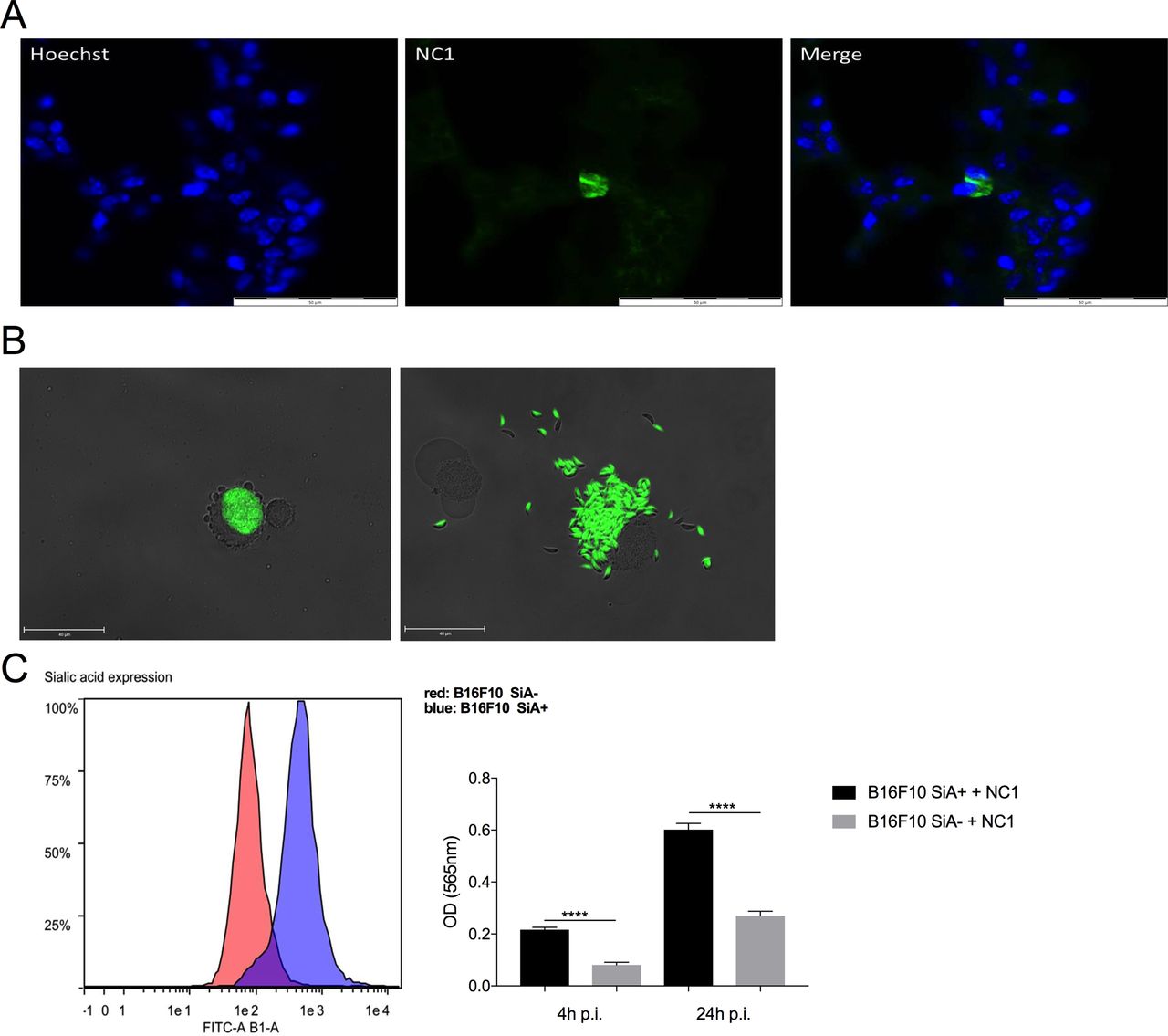 Figure 4
Figure 4 Mechanisms involved in Neospora caninum entry into cancer cells. Indirect immunofluorescence assays were performed on lung cryosections to reveal N. caninum tachyzoites 3 days after intranasal administration using a polyclonal N. caninum infection serum from rabbit and an Alexa 488 goat anti-rabbit IgG combined with a nucleus staining using Hoechst acquired at ×600 using Olympus IX73 microscope (A). B16F10 cells were infected with GFP-expressing N. caninum tachyzoites at MOI 1, and images acquired at 24 h p.i. at ×600 using Olympus IX73 microscope (B). B16F10 cells were desialylated thanks to neuraminidase treatment, desialylation was verified by flow cytometry using FITC-lectin (B). Desialylated B16F10 cells (B16F10 SiA−) and untreated B16F10 cells (B16F10 SiA+) were then cultured with β-galactosidase-expressing tachyzoites of N. caninum (NC1- βgal) at MOI 1 and incubated for 4 hours or 24 hours. Cells were then washed to dose intracellular β-galactosidase (B, right panel), two independent experimentations were performed and show similar results. ****p<0.0001. hpi, hours post-infection; MFI, Mean of Fluorescence Intensity; MOI, Multiplicity of Infection.
N. caninum treatment induces a reduction of size of human spheroids, and stimulates IFN-γ secretion from melanoma explantsThe study of N. caninum-induced antitumoral response in human models is a key step to further develop protozoan-based immunotherapy. Patient-derived spheroids were obtained from human melanoma metastasis biopsies using the hanging-drop method. Spheroids treated with 5000 N. caninum tachyzoites were significantly smaller than untreated spheroids 5 days after treatment (figure 5A). Moreover, spheroids treated by NC1-IL-15hRec were significantly smaller than NC1 treated spheroids 3 days after treatment, while no difference was observed at D5 (figure 5B). At this time point, treated spheroids, regardless of the strain, had shrunk to 75–80% of their original size while untreated spheroids had grown to 110% of their original size. Thus, N. caninum also possesses antitumoral properties in human melanoma cells from metastatic origin, whether it may be by direct oncolytic activity, by stimulation of the immune system, or both. To determine if N. caninum can stimulate an immune response in humans, 30 mg explants from four patient biopsies were treated with 107 NC1 and NC1-IL-15hRec tachyzoites. At both 24 hpi and 48 hpi a sixfold increase of IFN-γ production was found in explant supernatant with both NC1 and NC1-IL-15hRec groups, implying that N. caninum stimulates immune response in human tumor (figure 5D).
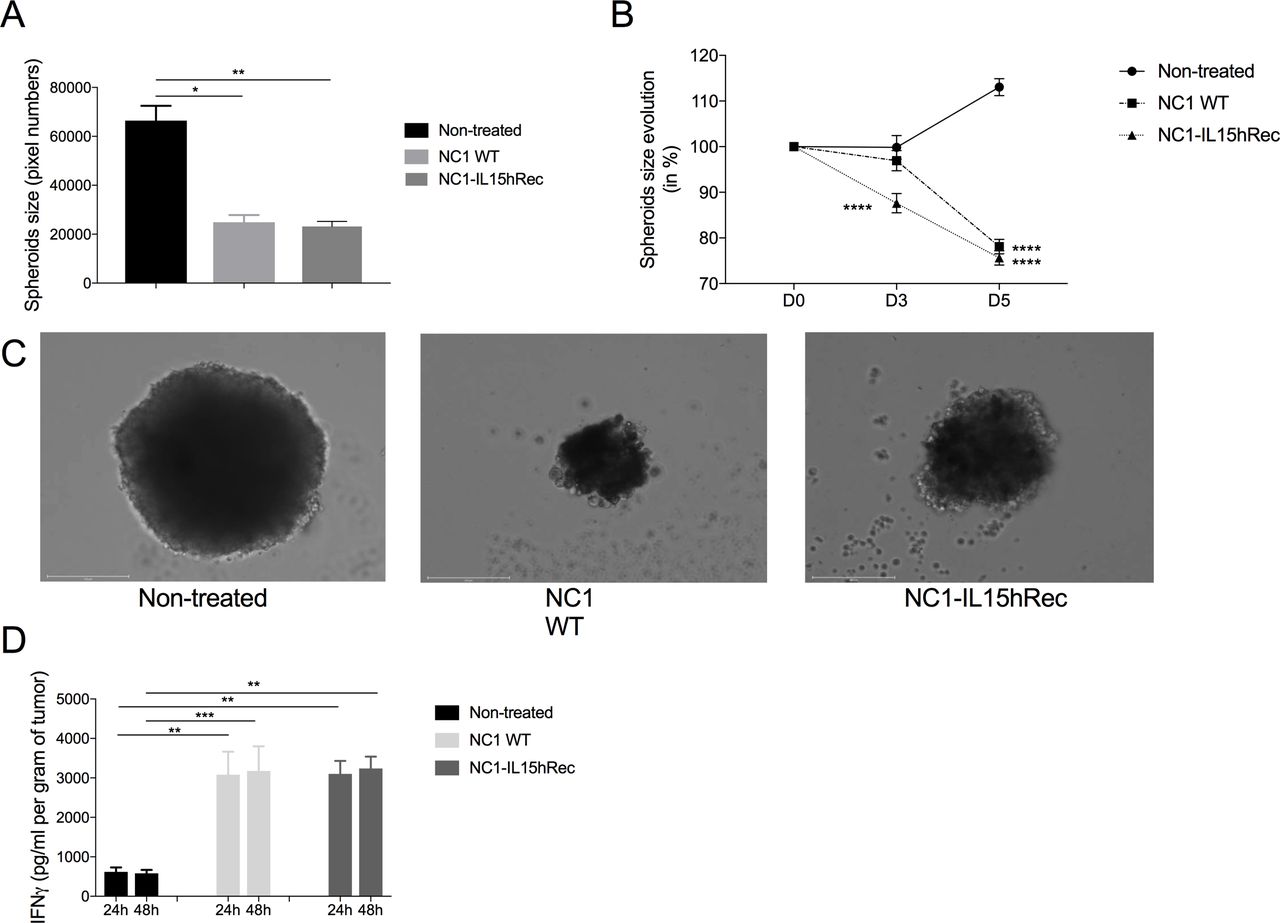 Figure 5
Figure 5 Neospora caninum treatment can control patient-derived-spheroid growth and induces a strong production of IFN-γ. Melanoma biopsies obtained from four patients were dissociated into a cell suspension and 10 µL containing 5000 cells were disposed in the lid of a Petri dish to obtain patient-derived spheroids through the hanging drop method. Culture media was changed every 2 days to ensure optimal growth conditions. Five days later, spheroids were treated with 5000 NC1 and NC1-IL-15hRec tachyzoites and spheroids sizes were monitored by microscopy for 5 days, n=10 spheroids per group (A, B, C). Melanoma biopsies were processed into 30 mg explants which were treated with 107 NC1 and NC1-IL-15hRec tachyzoites. Supernatants were collected 24 hpi and 48 hpi and IFN-γ was dosed to determine the immunostimulatory potential of N. caninum in humans (D). n=8 per group. **p<0.01; ****p<0.0001. IFN, interferon; IL, interleukin; hpi, hours post-infection.
DiscussionIn recent years, intracellular protozoa have been investigated to induce antitumor immune responses and to overcome tumor escape, thereby inhibiting tumor growth, angiogenesis, and metastasis in several animal models. Multiple immune cells activation pathways are involved: CD69/25, CD80/86, MHC-II activation, promoting the NK T cell to secrete TNF-α, CD 8+ T and NK cells to kill tumor through the IL-12/IFN-γ axis and stimulate B cells to produce specific IgG against tumor cells.34 We previously showed that N. caninum, a non-pathogenic protozoan for human, had great antitumor properties in an EG7 mouse model of flank tumor. Recently, another group have subsequently confirmed our results in a murine model of flank B16F10 melanoma tumor, highlighting the increasing interest for such therapies.35 In melanomas, several factors have been widely reported to facilitate tumor development such as epidermal growth factor, nitric oxide synthase, macrophage migration inhibitory factor and the proto-oncogene c-Myc. All the above factors are associated with poor patient prognosis through, respectively, facilitation of lymph node metastasis,36 inhibition of the tumor suppressor PTEN,37 induction of angiogenesis38 and vasculogenic mimicry.39 In our study, we found that expression of genes coding for those proteins is downregulated in the lungs of N. caninum treated mice. Several studies have shown that inhibiting those proteins and their interactions could lead to the development of new anticancer therapeutics.40–42
With N. caninum, such downregulation associated with all the above-mentioned mechanisms led to an interruption of metastatic development. However, treatment of advanced and/or refractory tumors might require enhanced antitumor responses and functions. This could be obtained by combinatory strategies or genetic engineering of oncolytic agent(s). Much of the enthusiasm surrounding engineered oncolytic therapies revolves around their appeal as multi-mechanism therapeutics.43 Accordingly, combination therapies are developed to improve the sensitivity of tumor cells to immunotherapy, accelerate the killing of tumor cells, and reduce the emergence of drug resistance by tumor cells.44 Thus, combining N. caninum with a potent effector could allow an improved efficacy, notably against refractory tumors.
In this study, we have investigated the therapeutic effect of an engineered N. caninum strain to secrete recombinant human IL-15 fused to the sushi domain of IL-15 receptor (NC1-IL-15hRec) in a syngeneic metastatic B16F10 lung mouse model. We revealed that intranasal administration of NC1-IL-15hRec induced higher number of tumor associated macrophages, NK cells and CD8+ T cells compared with intravenously administered NC1-ILhRec, highest expression of M1 markers on macrophages and an increased expression of genes coding for chemokines and antitumoral cytokines. Described phenomenon ceased metastasis development.
Clinical trials of IL-15 monotherapy for patients with advanced cancer have frequently resulted in stable disease as the best response, and many of these trials were limited by systemic toxicity.45 46 Interestingly, a recent study investigated an alternative route of administration, through an inhaled IL-15 therapeutic protocol, leading to improved protection against lung metastasis in dogs with naturally occurring osteosarcoma or melanoma with a clinical benefit rate of 39%.47 Inhaled cytokine therapy offers the benefit of local delivery and passive addressing of immunotherapy to the lungs, a frequent site of metastatic disease, while limiting systemic exposure and potential toxicity with most administered agent restricted at the tumor site. Inhaled IL-2 has also been used in human cancer trials with response rates of approximately 15% with no major side effects observed.48 Our approach to vectorize IL-15 with a protozoa locally delivered and replicating in the TME could then importantly reduce systemic toxicity of IL-15.
In addition, mucosal administration to the lung allows the targeting of specific mucosal immune cell populations. Indeed, the lungs are known to harbor a unique immune microenvironment, where tissue-resident NK and CD8+ T-cell immune populations display both a memory-like and heightened cytotoxic phenotype.49 Accordingly, those two populations were more abundant in the lung after NC1-IL-15hRec treatment, associated with improved protection. Our combinatory approach of oncolytic NC1-IL-15hRec administered intranasally could therefore meet all criteria for a highly effective, multi-mechanism strategy for elimination of lung metastases with a much-needed low toxicity thanks to local delivery. Only in the groups treated by intranasal administration of N. caninum, an induction of a rapid and local innate immune response in the lung was observed 2 days post-treatment. This innate immune response relies mainly on the increased number of macrophages. Such an increase is generally associated with poor prognosis, by contributing to tumor growth and metastasis progression50 as well as impairing clinical outcomes of anticancer therapies.51 Further analyses on these macrophages revealed that after N. caninum treatment, macrophages are polarized towards an M1 phenotype. This phenotype has antitumor effects with its ability to identify tumor cells and kill them directly through Nitric Oxyde (NO) and Reactive Oxygen Species (ROS) production or indirectly by antibody-dependent cell-mediated cytotoxicity.52 In our study, immunostaining of macrophages on cryosections of lung treated by N. caninum disclosed numerous intratumoral macrophages while macrophages in untreated lungs seem to be circumscribed only to the periphery of metastases. Thus, the increase of intratumoral macrophages and their polarization towards an M1 phenotype induced by intranasal administration of N. caninum are potent mechanisms leading to early metastases regression. However, it would be interesting to investigate how advanced, longly established metastatic tumors respond to N. caninum therapy. In the B16F10 model, lung metastases are present at the time of Neospora treatment, but treating tumors at much later time points would be of interest for translation to
留言 (0)