
記住我

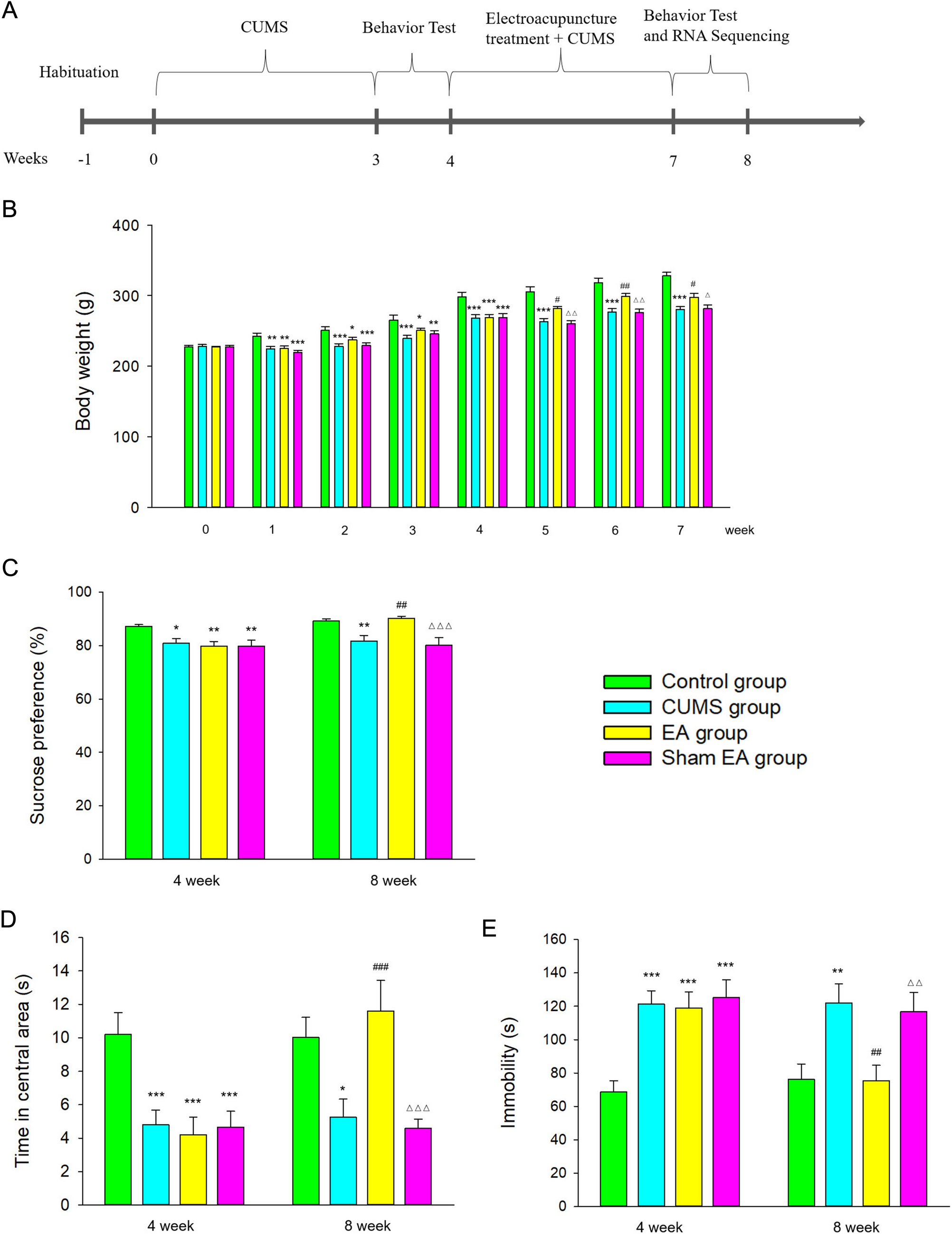



PD is one of the most prevalent neurodegenerative disorders worldwide. Motor symptoms of Parkinson’s include bradykinesia, resting tremor, and cogwheel rigidity whereas non-motor symptoms include sleep disturbance, cognitive decline, and depression [67]. The pathophysiology of PD involves the progressive destruction of the dopaminergic neurons in the nigrostriatal pathway in the midbrain [68]. Accumulation of ROS can damage the DNA leading to oxidative stress and cell death are associated with PD [69].
Several models of PD have been proposed in order to better understand this disorder. Examples include the use of the neurotoxin 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) and its active metabolite 1-methyl-4-phenylpyridinium (MPP+). MPP+ plays an active role in destroying the dopaminergic neurons that results in parkinsonism [70]. An in vitro study used the human neuroblastoma cells (SH-SY5Y) to investigate the role of H2S in attenuating the oxidative stress on the neurons in PD cell model [40]. This study used an MPP+-treated SH-SY5Y cell model showing that NaHS increased the cell viability, reduced cytotoxicity, and reduced oxidative stress-induced cell apoptosis in a dose dependent manner in the MPP+ treated cells [40].
Another study investigated the neuroprotective role of H2S in a mouse model of PD. In line with the study carried out by Liu et al. [40], this study also showed that H2S elicited neuroprotective properties. MPTP was used in this study to generate a mouse model of PD and NaHS was used to investigate the effect of H2S. This study demonstrated that H2S prevents MPTP-induced neuronal damage and promotes neurogenesis through Akt/glycogen synthase kinase-3β (GSK-3β)/β-catenin pathways in adult neuronal stem cells [42]. Consistently, in a similar model of MPTP-induced PD in mice established by Lu et al., it was observed that NaHS reduced primary mesencephalic neurons cytotoxicity whilst explored molecular mechanism associated highlighting that H2S was able to enhance mitochondrial uncoupling protein 2 (UCP2) antioxidation resulting in abrogated ROS generation and reduced caspase 12-induced apoptosis [43]. Mitochondria are the main site of ROS generation thus may play a key role in the development of PD whilst compounds modulating mitochondrial-ROS, such as H2S, may be potential therapeutic candidates in the treatment of PD.
Although the molecular mechanisms of H2S-induced neuroprotection in PD may be broad (Fig. 3), these new avenues in PD research position H2S as a potential mitochondrial protectant. Thus, H2S may be a suitable therapeutic candidate for clinical use in treating Parkinson’s in the future.
Fig. 3
Protective molecular mechanisms of H2S in Parkinson’s disease (PD). The neuroprotective effects of H2S in PD are associated with neurogenesis mediated by GSK3β/β-catenin pathway. H2S has antioxidant properties observed by the induction of antioxidant defence mediated by upregulation of superoxide dismutase (SOD) and glutathione (GSH). H2S can also reduce levels of reactive oxygen species (ROS) acting as a direct scavenger or indirectly by restoring the mitochondrial membrane potential (ΔΨm). The reduction in accumulation of mitochondrial-ROS results in inhibition of apoptosis mechanisms via caspase 3. Moreover, H2S may also regulate parkin functions by mediating posttranslational modifications of active cysteine residues, a process known as S-sulfhydration or persulfidation
Alzheimer’s Disease (AD)AD is the most common neurodegenerative disorder and is the leading cause of dementia in the elderly population. Oxidative stress, neuroinflammation and damage to cholinergic neurons are some of the pathophysiological changes that are involved in AD [71].Furthermore, the accumulation of the microtubule-associated protein Tau [72] and β-amyloid peptides in neuronal cells [73] have been proposed as key molecular mechanism leading to this devastating disorder.
A research study investigated the neuroprotective role of H2S in an animal model of AD by using the 3xTg-AD mouse model which includes Tau protein mutations: PS1M146V, APPSwe, and Tau P301L [46]. This study showed that H2S prevents hyperphosphorylation of the Tau protein by S-sulfhydration (post-translational modification of proteins mediated by H2S) of GSK3β. Additionally, it was observed that the CSE catalytic effect was augmented by binding wild type Tau which resulted in decreased levels of Tau protein in the cell. In contrast, CSE could not bind the mutated Tau P301L. Consequently, CSE levels were decreased in the cortex and hippocampus of the brain in the 3xTg-AD mouse model compared to wild type. This interesting report demonstrated a direct relationship between decreased endogenous levels of H2S and AD-like condition in vivo. Interestingly, it was demonstrated that administrating H2S donor, GYY4137, by daily intraperitoneal injection for 12 weeks (100 mg/kg) to 3xTg-AD mice improved their motor and cognitive function. The level of Tau protein S-sulfhydration was higher in the group treated with GYY4137 compared to the control group [46].
Aligned with the study carried out by Giovinazzo et al. [46], a study by Vandini et al. [47] using 3xTg-AD transgenic mouse model, previously showed that intraperitoneal injection of sulphur water and NaHS daily for three consecutive months improved memory and cognitive functions in both young and aged animals Additionally, treatment with NaHS and sulphur water in 3xTg-AD mouse decreased the size of the β-amyloid plaques in the cortex and hippocampus. The molecular mechanisms implicated suggest the inhibition of c-Jun N-terminal kinases, extracellular signal-regulated kinases, and p38 protect against neuroinflammation and Tau protein hyperphosphorylation, leading to a decrease in accumulation of β-amyloid plaques in the cortical and hippocampal regions of the brain [47].
Moreover, a recently synthesised H2S donor targeted to the mitochondria, AP39, was probed against a model of AD in mice (APP/PS1 double-transgenic mice) [49]. AP39 injected intraperitoneally for 6 months enhanced cellular bioenergetic function and had mitochondrial protective effect on AD neurons and mice. Neurons treated with AP39 showed reduced ROS levels and increased ATP production. Within the AD neurons, it was observed that mitochondrial DNA was damaged, however AP39 treatment prevented these alterations by increasing mitochondrial DNA integrity. Furthermore, APP/PS1 double-transgenic mice treated with AP39 observed an improvement in their learning and memory impairments [49] suggesting that H2S, targeted to the mitochondria, could be a potential therapeutic candidate for AD. Given the fact that accumulation of Tau protein disturbs the neuronal mitochondrial respiration, molecular mechanism implied may include the restoration of the flow of electrons in the oxidative phosphorylation and/or antioxidant effects abrogating neurotoxicity induced by Tau, however, these theories warrant further exploration.
Overall, H2S has proven to play a significant neuroprotective role in AD models (Fig. 4). However, the translation of these results to the clinical setting might be hindered by complex dosage regimen of the H2S donor, requiring multiple and extensive dose regimes (3 to 6 months) as observed in these studies [46, 47, 49]. This emphasises the necessity to further explore patient-friendly approaches including better delivery systems and/or more stable H2S donors to reduce frequency of administration.
Fig. 4
Protective mechanisms of H2S in AD. The neuroprotective effects of H2S in AD are associated with inhibition of anti-inflammatory cascade via reduction of TNF-β and IL-6 levels. Moreover, H2S has been linked to reduction of size of β-amyloid plaques and reduction in hyperphosphorylation of Tau. H2S has been observed to reduce levels of mitochondrial reactive oxygen species (ROS) leading to inhibition of apoptosis via caspase 3. H2S may also maintain the integrity of mitochondrial DNA whilst it may also regulate GSK3β protein functions via S-sulfhydration (persulfidation)
Huntington’s Disease (HD)HD is an autosomal dominant progressive neurodegenerative disorder resulting from the toxic accumulation of mutant huntingtin protein in neurons [74]. The pathophysiology of HD involves the selective destruction of the corpus striatum in the brain which controls motor activities leading to involuntary and irregular muscle movements, behavioural changes and cognitive decline [75].
Paul et al. [76] explored the neuroprotective role of H2S in a mouse model of HD showing a direct correlation between reduction in CSE levels and increased neurodegeneration in the striatum. This study used both in vivo and in vitro models to demonstrate the impact of CSE depletion in the pathophysiology of HD. CSE depleted (CSE−/−) mice model exhibited abnormal hindlimb clasping and clenching resembling HD. Additionally, this study demonstrated that CSE level was significantly decreased in the striatal cell line of HD cell model consisting of 111 glutamine repeats (STHdhQ111/Q111(Q111) compared to the control group [76]. These observations suggest the importance of H2S in HD and imply the potential application of H2S donors in halting the progression of the disorder.
In an attempt to explore the potential for NaHS in abrogating HD-like effects induced by 3-nitropropionic acid (3NP), a study in a rat HD model revealed that H2S improved overall cognitive and locomotor deficits whist provided antioxidant, anti-inflammatory and anti-apoptotic effects observed by reduced levels of oxidative stress marker malondialdehyde (MDA), TNF-α and caspase 3 activation [51]. Interestingly, this study also revealed that treatment with NaHS significantly enhanced CBS expression, suggesting that H2S availability may have a crucial effect in modulating endogenous H2S pathways. Nonetheless, these assumptions require further exploration to understand the potential molecular mechanisms implicated.
Cerebral Vascular DiseaseStroke is the second most prevalent cause of mortality worldwide. The two types of strokes include haemorrhagic and ischemic stroke [77]. Both lead to ischemia and consequent tissue damage/death.
Given the association between ischemic stroke and ROS, H2S as an antioxidant compound, may be a suitable candidate for management of these conditions. A study investigated the neuroprotective role of H2S in acute ischemic stroke using a model of cerebral ischemia/reperfusion injury in mice by administrating NaHS before reperfusion following an ischemic insult in the brain. By means of H-magnetic resonance spectroscopy/magnetic resonance spectroscopy (H-MRI/MRS) and immunohistochemistry, it was shown that the total infarct volume was reduced compared to sham, and administration of NaHS 1 min before perfusion exerted stronger effect that of 30 min [52]. Another study exploring the effect of NaHS on transient cerebral ischemia immediately after reperfusion, this, combined with mild hypothermia resulted in a decrease in ischemic-reperfusion injury via upregulation of N-methyl-D-aspartate receptor (NMDAR) [53].
To study mechanisms of haemorrhagic stroke in vivo, models such as the middle cerebral artery occlusion (MCAO) plus intravenous injection of tissue plasminogen activators (tPA), have been used [78]. Using this model, it was demonstrated that H2S reduces tPA-induced cerebral haemorrhage following MCAO. In this study, co-administration of two structurally distinct H2S donors, ADT-OH and NaHS reduced the tPA-induced cerebral haemorrhage through the inhibition of AKT-VEGF-MMP9 signalling cascade [55]. As one of the risk factors associated with tPA in the management of ischemic stroke, is the increased risk of cerebral haemorrhage, observations by Liu et al. [55] suggest that H2S donors may reduce the risk of haemorrhagic stroke in tPA-exposed subjects. Although more research is necessary to confirm this hypothesis, the co-administration of H2S and tPA may bring new avenues and may provide patients requiring treatment for ischemic stroke a better recovery profile.
Brain TumoursBrain tumours carry a high morbidity and mortality rate which highlights their clinical significance and burden to health systems worldwide. The most malignant brain tumour is glioblastoma multiform (GBM) which can rapidly spread and invade brain parenchyma [79]. Despite the growing number of treatment options available for the patients, brain malignancies carry a poor prognosis [80].
An in vitro study investigated the anti-cancer properties of H2S in C6 glioma cells [58]. It was shown that NaHS induces C6 glioma cell apoptosis via upregulation of caspase 3 and Bax proteins and downregulation of Bcl-2 protein through p38 MAPK signalling pathway [58]. In contrast to this report, another in vitro study demonstrated that H2S promotes C6 glioma cell growth. However, when CBS activity was compromised, the enhanced cell proliferation was blunted [60]. This study showed that cell apoptosis was reduced resulting in increased proliferation and viability rates of C6 glioma cells when exposed to NaHS suggesting that p38 MAPK/ERK1/2-COX-2 pathways are involved in NaHS-induced cancer cell proliferation and anti-apoptosis in C6 glioma cells [60]. An in vivo study reported by Li et al. showed that the size of the tumour was significantly increased in the rats exposed to NaHS in GBM group compared to the GBM only group, and H2S promotes C6 glioma cell growth via augmenting neurovascular formation and increasing hypoxia-inducible factor-1alpha (HIF-1α) expression [61]. These contrasting results more investigation is required to better understand the role of H2S in promoting/mitigating the expansion of brain tumours.
EpilepsyEpilepsy is one of the most common neurological disorders affecting around 50 million of the global population. This condition is characterised by recurrent seizures which is the clinical manifestation of alerted neuronal electrical activities [81]. Additionally, despite antiepileptic drugs offering many patients symptomatic relief, effective management is not achieved in almost one third of patients with current pharmacotherapies. Hence, new medical therapies for epilepsy are needed [82].
Pro- inflammatory cytokines such as IL-1β, IL-6, and TNF-α play an important role in the pathophysiology of the epileptic seizures and the level of these cytokines is raised in the serum and cerebrospinal fluid of epileptic patients [83]. Increased level of pro-inflammatory cytokines results in increased excitability of neurons which may be crucial in the pathophysiology epilepsy [84]. As H2S promotes the release of anti-inflammatory cytokines and is involved in preventing neuroinflammation [85], it might have a potential clinical application in the treatment of epilepsy.
The role of H2S in pilocarpine-induced status epilepticus (SE) in vivo has been explored. Adult male C57BL/6 mice were exposed to pilocarpine to induce SE whilst treatment consisted of exposure to a novel carbazole-based H2S donor. This study demonstrated that SE + H2S group had a shorter duration of seizure compared to the SE group. Additionally, this group showed delayed onset of seizure, reduced damage to the hippocampus and reduced pro-inflammatory state in microglia compared to the SE control group [62].
In contrast, a study performed by Luo et al. [63] showed that H2S delivered as NaHS exacerbates Pentylenetetrazole (PTZ)- and pilocarpine‐induced seizures in rats [63]. PTZ is a GABA-A receptor antagonist that when sequentially injected to animals, results in the development of chemical kindling, an epilepsy model [86]. Luo et al. [63] exposed rats to PTZ/pilocarpine and PTZ/pilocarpine + NaHS showing that the severity of seizures increased in the group treated with NaHS compared to the control group. Furthermore, the duration of seizure was longer in those rats exposed NaHS suggesting that H2S increased membrane excitability of entorhinal cortex neurones via facilitating the function of voltage‐gated sodium channel, AMPAR, and NMDAR [63]. These observations are not consistent about the effect of exogenous H2S on the manifestation of epilepsy. However, they suggest a potential role of H2S signalling in the modulation and severity of epilepsy. Further research is warranted to better understand the potential of the modulation of H2S signalling in the management of epilepsy.
Multiple Sclerosis (MS)MS is an autoimmune disease affecting the myelin structure in the CNS. Currently, 2.8 million people are affected worldwide, however, this number is on the rise [87]. MS results in a wide range of signs and symptoms that are variable depending on the severity and location of the lesions with optic neuritis being the most common [88]. Currently, the available immune-based therapies do not significantly improve MS prognosis, therefore, new treatment strategies, such as H2S donors, warrant exploration in the pursuit of better patient outcomes.
In physiological settings, the BBB protects the CNS by making it inaccessible to immune cells, while in MS, the BBB gets damaged due to endothelial cells’ impairment [89]. This results in an exacerbated influx of inflammatory mediated cells into the brain tissue, which induces demyelination and axonal dysfunction. Recurrent inflammatory responses in MS irreversibly damage the CNS nerve cells with the involvement of ROS [90]. The pathophysiology of MS also involves chronic platelet activation that promotes differentiation of autoreactive T cells resulting in initiation and progression of autoimmune neuroinflammation [91]. Therefore, two potential molecular mechanisms by which H2S may show potential beneficial effects in MS include its antioxidant role as well as its ability to inhibit platelet activation and adhesion molecule-mediated aggregation [92].
An in vitro study by Talaei et al. [65] examined the effect of NaHS on peripheral blood mononuclear cells (PBMNCs) obtained from healthy individuals to access its transmigration across an endothelial cells’ barrier representing the BBB. This study showed that pre-treatment of human endothelial cells with NaHS decreased PBMNC transmigration of both control and serotonin-treated cells. Additionally, treatment with NaHS resulted in upregulation of IL-10 and downregulation of adhesion molecules: lymphocyte function-associated antigen 1 (LFA-1) and vascular cell adhesion protein 1 (VCAM-1). This suggests that whilst NaHS resulted in relaxation/expansion of endothelial cells it may also cause morphological changes to the BBB leading to increased BBB permeability during MS attacks [65].
Lazarević et al. [66] studied the in vitro effect of H2S donor GYY4137 on differentiated dendritic cells and T cells which are both involved in the pathogenesis of MS. GYY4137 enhanced transforming growth factor β (TGF-β) expression in dendritic cells, suggesting an anti-inflammatory effect of GYY4137 in an MS-like environment. Additionally, isolated lymph node and spinal cord T cells were obtained from mice and rats immunised with CNS antigens and treated with GYY4137. GYY4137 significantly reduced interferon gamma (IFN-γ) and IL-17 production and reduced the proportion of FoxP3 + regulatory CD4 + T cells in the lymph node and spinal cord T cells. Interestingly, when the expression of H2S-producing enzymes (CBS, CSE, and 3-MST) was assessed in immune cells from healthy donors and drug-naïve relapse-remitting MS patients, the authors reported no differences in the CSE and CBS expression but lower expression of 3-MST [66].
Given the relevance of 3-MST in the generation and availability of H2S within the mitochondria, this study further suggests a potential link between H2S signalling and mitochondria in the modulation of anti-inflammatory response in MS. However, this theory awaits more exploration.
留言 (0)