
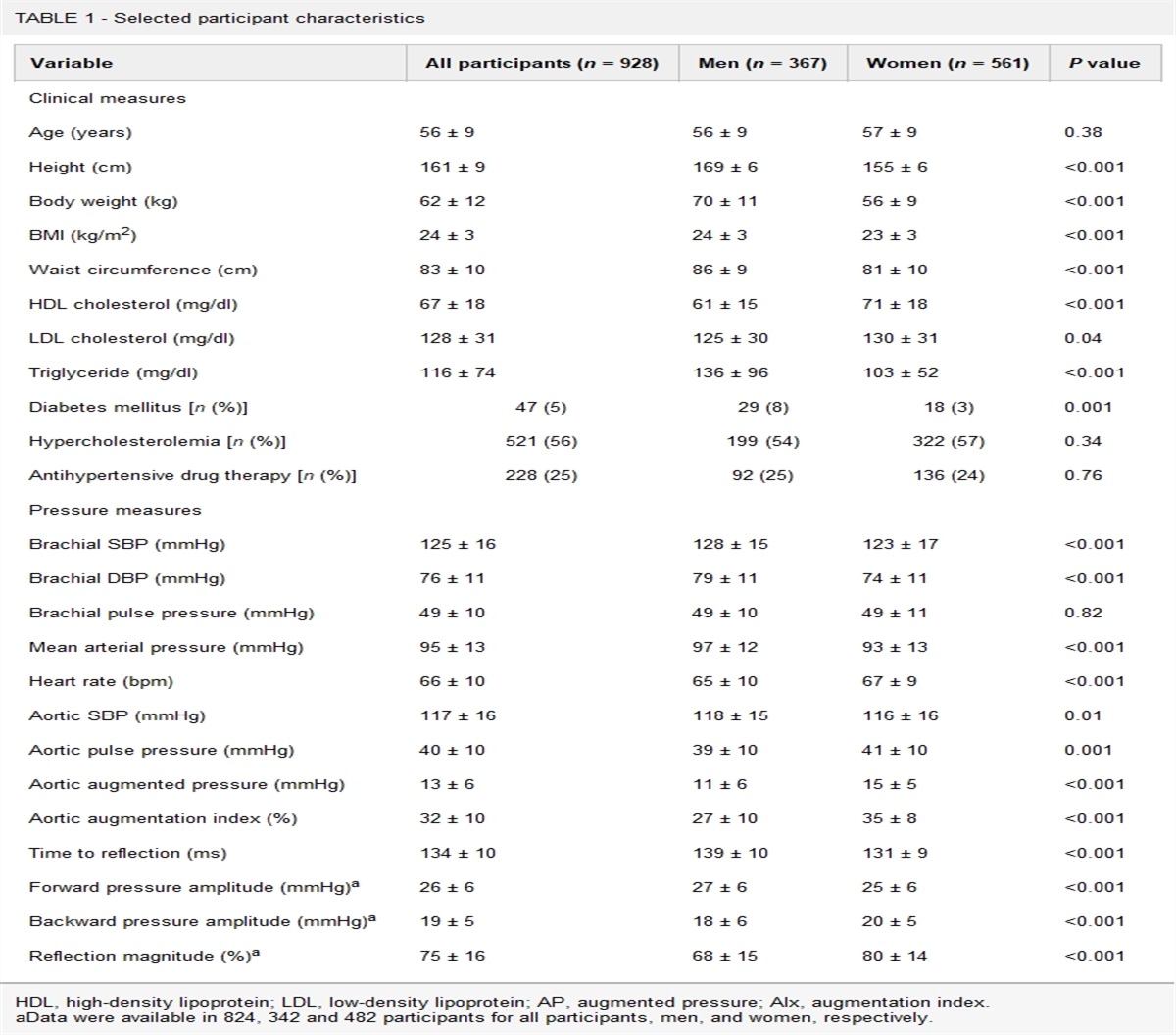
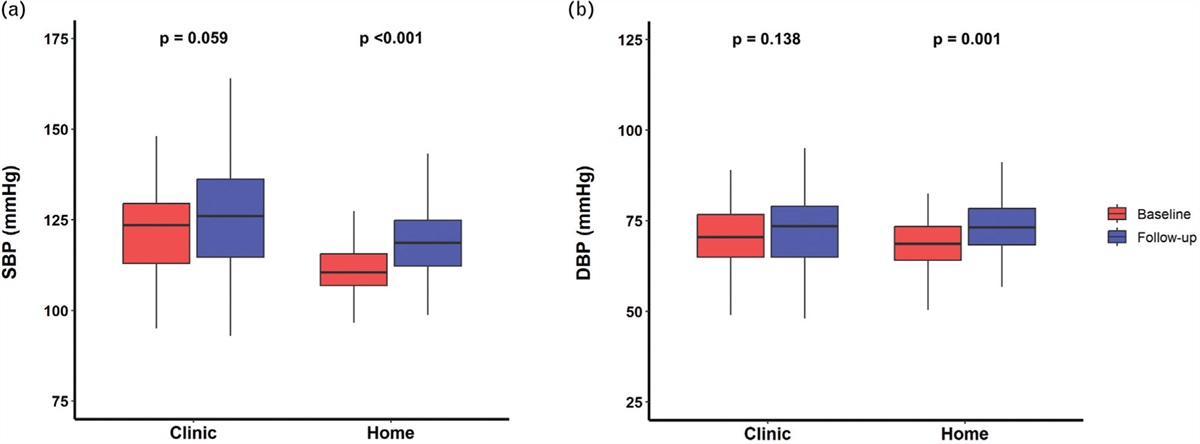
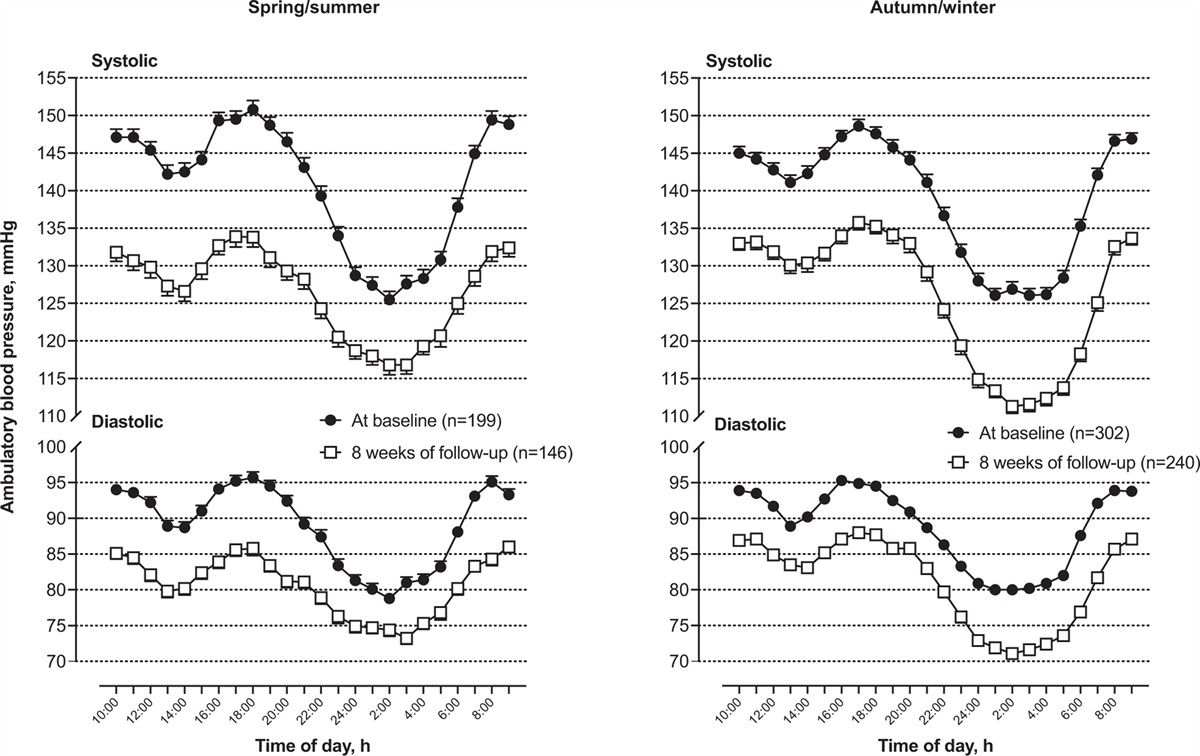
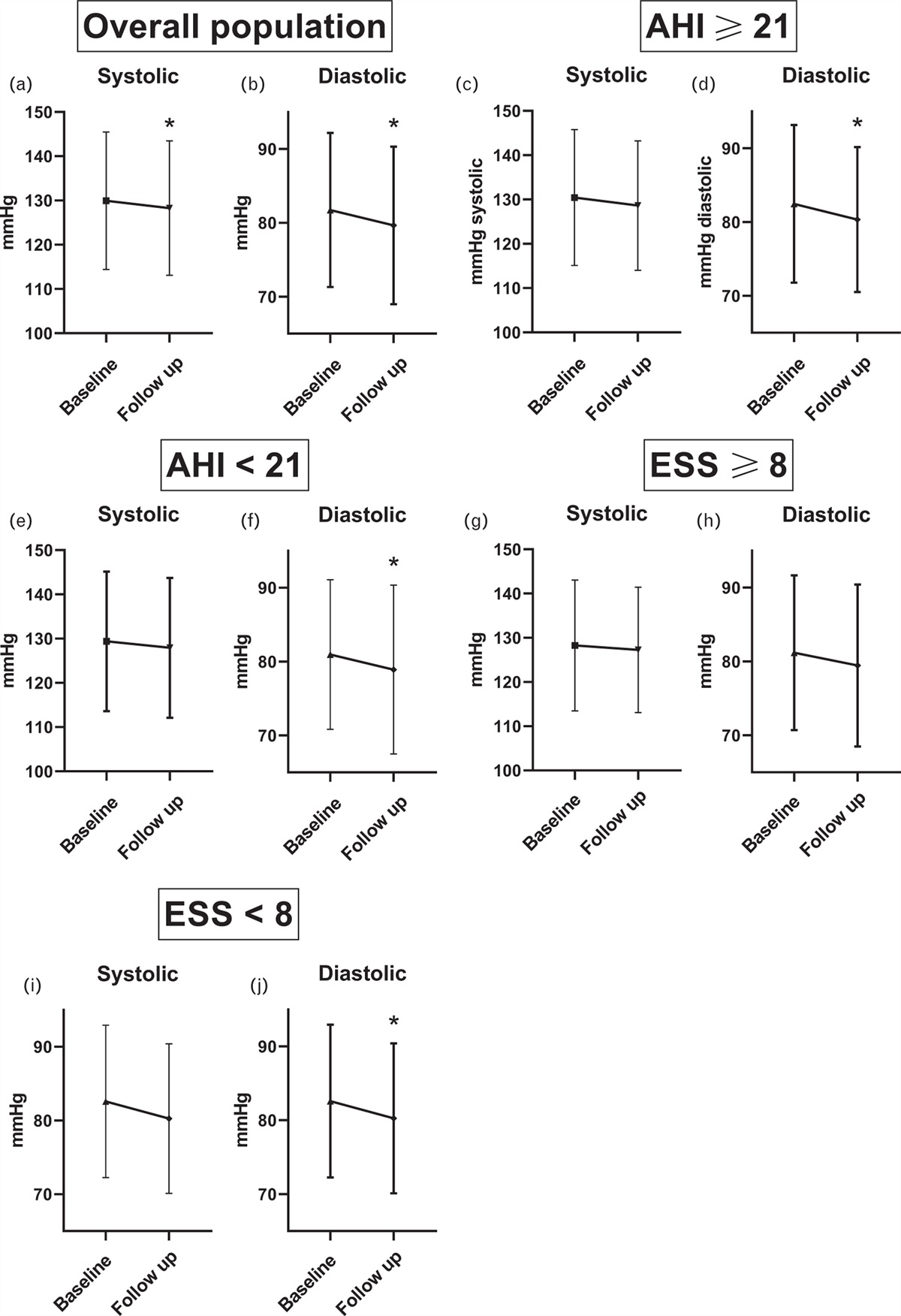
記住我

The kidney is a major site for hypertension target organ damage. High-uncontrolled blood pressure initiates the hypertension nephropathy, and then, nephropathy can further augment hypertension severity, enhancing progression to end-stage renal disease and kidney transplantation. Hypertensive kidney injury is characterized by renal fibrosis, tubular hypertrophy and glomerular alterations [1]. Fibrosis is a normal repair process triggered in response to injury; it preserves tissue/organ architecture and functional integrity. However, deregulation of fibrosis leads to pathological accumulation of extracellular matrix, mostly collagens. In chronic kidney disease, collagen replaces lost podocytes; also, tubular cell injury leads to tubulointerstitial fibrosis, which may continue to nephron loss [2]. Progression to tubulointerstitial fibrosis correlates strongly with proteinuria and disease severity [3]. Chronic kidney disease caused by hypertension is a strong independent risk factor for adverse cardiovascular outcomes.
Excessive production of collagen by myofibroblasts is a relatively late event in the pathophysiology of chronic hypertensive kidney disease and currently is a major focus for therapy [4]. However, earlier stages involving interactions of stressed kidney cells with immune cells are very likely equally important. Recently, STING inhibition has been shown to attenuate renal fibrosis [5]; STING is a cytoplasmic DNA sensor that is frequently activated in stressed cells. STING activation is an early signalling event in the development of renal fibrosis and plays a crucial role in notifying immune cells of unusual cellular stress. Also, production of interleukin (IL)-18 by renal tubular epithelial cells activates kidney T cells during steroid/salt-induced hypertension and promotes inflammation and injury, reflected by albuminuria and fibrosis [6]. Chemokines are also important; CXCR3 chemokines are higher in patients with hypertensive nephrosclerosis than in normotensive subjects and account for large influxes in kidney CD4+ and CD8+ T cells, which may be responsible for increases in severely damaged/necrotic kidney cells [7,8]. Mitochondria are a major source of reactive oxygen species in the kidney that may also contribute during early stages of disease. The mechanisms that drive many of these processes in hypertensive chronic kidney disease are yet to be fully defined.
In this issue of Journal of Hypertension, Gao et al. [9] present an interesting study on the role of the transcription factor Interferon Regulatory Factor 4 (IRF4) in the development of kidney inflammation and fibrosis in deoxycorticosterone/salt-induced hypertension. IRF4 expression is largely restricted to immune cells such as T and B lymphocytes, macrophages and dendritic cells, wherein it regulates differentiation and is required for lymphocyte activation. The authors demonstrate that IRF4-expressing cells localize to the renal tubulointerstitium, largely composed of tubular epithelial cells, interstitial cells, peritubular capillary endothelium, pericytes and extracellular matrix. They identify the cells as CD4+ T cells, CD8+ T cells and F4/80+ macrophages. Deleting IRF4 did not affect blood pressure in the hypertensive mice but ameliorated albuminuria and nearly completely prevented CD4+ and CD8+ T-cell accumulation. These effects are consistent with important roles for IRF4 in naive CD4+ T-cell differentiation into T-helper cells, the differentiation of cytotoxic effector CD8+ T cells and CD8+ T-cell memory formation. The dependency of IL-6 mediated inflammation on IRF-4 is most likely due to effects on CD4+ T cells. Unfortunately, the authors did not examine whether CD4+ T-cell activation helps sustain CD8+ T-cell proinflammatory effects and whether the CD8+ T cells contribute to kidney damage via cytotoxic mechanisms, as has been reported in hypertensive cardiac fibrosis [10]. CD8+ T cells are increased in kidneys of humans and mice with segmental glomerulosclerosis and diabetic kidney disease wherein they promote podocyte injury and glomerulosclerosis [11]. The authors demonstrate less severe kidney injury in hypertensive mice upon deleting IRF4 presumably due to effects on CD4+ and CD8+ T cells. In the future, it will be important to define the mechanisms by which these cells promote renal injury and whether they are responding to STING activation in target cells.
The authors demonstrate important roles for IRF4 in the development of renal fibrosis. Myofibroblasts are major fibrogenic cell types that are rare in normal kidneys but greatly increase during chronic hypertensive kidney disease. Myofibroblasts can derive from multiple cell types, for example, interstitial stromal cells, including fibroblasts, pericytes and mesenchymal stem cells; bone marrow derived cells such as macrophages; tubular epithelial cells; and endothelial cells. The authors focus on myeloid myofibroblasts and implicate IRF-4 in their accumulation. They provide evidence that IRF-4 is critically important for macrophage to myofibroblast transition, specifically M2 macrophages, via the PI3K/AKT pathway. Because global IRF-4 knockout mice were used, it is not possible to assess the importance of myeloid myofibroblasts relative to myofibroblasts derived from other cell types. Although collagen deposition was reduced substantially – by approximately 50% – it was not completely abolished, suggesting possible contributions from other myofibroblasts not dependent on IRF4. The reported dependency of transforming growth factor (TGF)-β1, interleukin (IL)-1β and MCP-1 on IRF-4 is most likely due to effects on macrophages; macrophages are important drivers of TGF-β1 fibrosis [10]. Although the current findings provide strong evidence for a role of IRF4 in renal inflammation and fibrosis in deoxycorticosterone/salt-induced hypertension, caution should be taken before extrapolating these finding to other forms of hypertension. Both T cells and macrophages express mineralocorticoid receptors, which affects their activity [12]. Thus, the effects observed in the deoxycorticosterone-salt hypertensive mice might include an IRF-4 component that is independent of the hypertension. Additional studies will be required in other hypertensive models of renal disease to confirm the magnitude of IRF-4's effects in hypertensive nephropathy.
The finding that IRF-4 is important for development of hypertensive kidney inflammation and fibrosis in a mouse model raises the question about translation to hypertensive humans. IRF-4 expression in renal inflammation and fibrosis is strongly linked with IL-4 expression by CD4+ Th2 and iNKT cells [13]; IL-4 strongly activates IRF-4 expression in human immune cells [14,15]. These findings, together with reports of a strong association of variable number of tandem repeat (VNTR) polymorphism in the IL-4 gene in humans with end-stage renal disease, suggest that IRF-4 is also important for progression of renal inflammation and fibrosis in humans (Fig. 1). More detailed investigations are required to confirm these associations in hypertensive chronic renal disease.
 FIGURE 1: Relationship between IL-4 gene VNTR polymorphism and end-stage renal disease in humans. IL-4 upregulates IRF4 in key human immune cells and myeloid fibroblasts by enhancing expression and activity of jumonji domain-containing protein 3 (JMJD3) demethylase [14], leading to hypertension nephropathy and end-stage renal disease.ACKNOWLEDGEMENTS
Conflicts of interest
FIGURE 1: Relationship between IL-4 gene VNTR polymorphism and end-stage renal disease in humans. IL-4 upregulates IRF4 in key human immune cells and myeloid fibroblasts by enhancing expression and activity of jumonji domain-containing protein 3 (JMJD3) demethylase [14], leading to hypertension nephropathy and end-stage renal disease.ACKNOWLEDGEMENTS
Conflicts of interest
There are no conflicts of interest.
REFERENCES 1. Ruiz-Ortega M, Rayego-Mateos S, Lamas S, Ortiz A, Rodrigues-Diez RR. Targeting the progression of chronic kidney disease. Nat Rev Nephrol 2020; 16:269–288. 2. Meng XM, Nikolic-Paterson DJ, Lan HY. Inflammatory processes in renal fibrosis. Nat Rev Nephrol 2014; 10:493–503. 3. Wehrmann M, Bohle A, Held H, Schumm G, Kendziorra H, Pressler H. Long-term prognosis of focal sclerosing glomerulonephritis. An analysis of 250 cases with particular regard to tubulointerstitial changes. Clin Nephrol 1990; 33:115–122. 4. Li L, Fu H, Liu Y. The fibrogenic niche in kidney fibrosis: components and mechanisms. Nat Rev Nephrol 2022; 18:545–557. 5. Zeng H, Gao Y, Yu W, Zhong C, Su X, Wen S, et al. Pharmacological inhibition of STING/TBK1 signaling attenuates myeloid fibroblast activation and macrophage to myofibroblast transition in renal fibrosis. Front Pharmacol 2022; 13:940716. 6. Thomas JM, Ling YH, Jelinic M, Sharma P, Saini N, Ferens DM, et al. IL-18 (Interleukin-18) produced by renal tubu940716.lar epithelial cells promotes renal inflammation and injury during deoxycorticosterone/salt-induced hypertension in mice. Hypertension 2021; 78:1296–1309. 7. Youn J-C, Yu HT, Lim BJ, Koh MJ, Lee J, Chang D-Y, et al. Immunosenescent CD8+ T cells are C-X-C chemokine receptor type 3 chemokines are increased in human hypertension. Hypertension 2013; 62:126–133. 8. Abdelbary M, Rafikova O, Gillis EE, Musall JB, Baban B, O’Connor PM, et al. Necrosis contributes to the development of hypertension in male but not female, spontaneously hypertensive rats. Hypertension 2019; 74:1524–1531. 9. Gao Y, Liu B, Guo X, Nie J, Zou H, Wen S, et al. Interferon regulatory factor 4 deletion protects against kidney inflammation and fibrosis in deoxycorticosterone acetate/salt hypertension. J Hypertens 2023; 41:794–810. 10. Brassington K, Kanellakis P, Cao A, Toh BH, Peter K, Bobik A, et al. Crosstalk between cytotoxic CD8+ T cells and stressed cardiomyocytes triggers development of interstitial cardiac fibrosis in hypertensive mouse hearts. Front Immunol 2022; 13:1040233. 11. Li L, Tang W, Zhang Y, Jia M, Wang L, Li Q, et al. Targeting tissue-resident memory CD8+ T cells in the kidney is a potential therapeutic strategy to ameliorate podocyte injury and glomerulosclerosis. Mol Ther 2022; 30:2746–2758. 12. Munoz-Durango N, Arrese M, Hernandez A, Evelyn E, Kalgeris AM, Cabrera D. A mineralocorticoid receptor deficiency in myeloid cells reduces liver steatosis by impairing activation of CD8+ T cells in a nonoalcoholic steatohepatitis mouse model. Front Immunol 2020; 11:563434. 13. Liu B, Jiang J, Liang H, Xiao P, Lai X, Nie J, et al. Natural killer T cell/IL-4 signaling promotes bone marrow-derived fibroblast activation and M2 macrophage-to myofibroblast transition in renal fibrosis. Int Immunopharmacol 2021; 98:107907. 14. Hsu AT, Lupancu TJ, Lee M-C, Fleetwood AJ, Cook AD, Hamilton JA, et al. Epigenetic and transcriptional regulation of IL4-induced CCL17 production in human monocytes and murine macrophages. J Biol Chem 2018; 293:11415–11423. 15. Vasudevan R, Norhasniza MN, Patimah I. Association of variable number of tandem repeats polymorphism in the IL-4 gene with end-stage renal disease in Malaysian patients. Genet Mol Res 2011; 10:943–947.
留言 (0)