
記住我
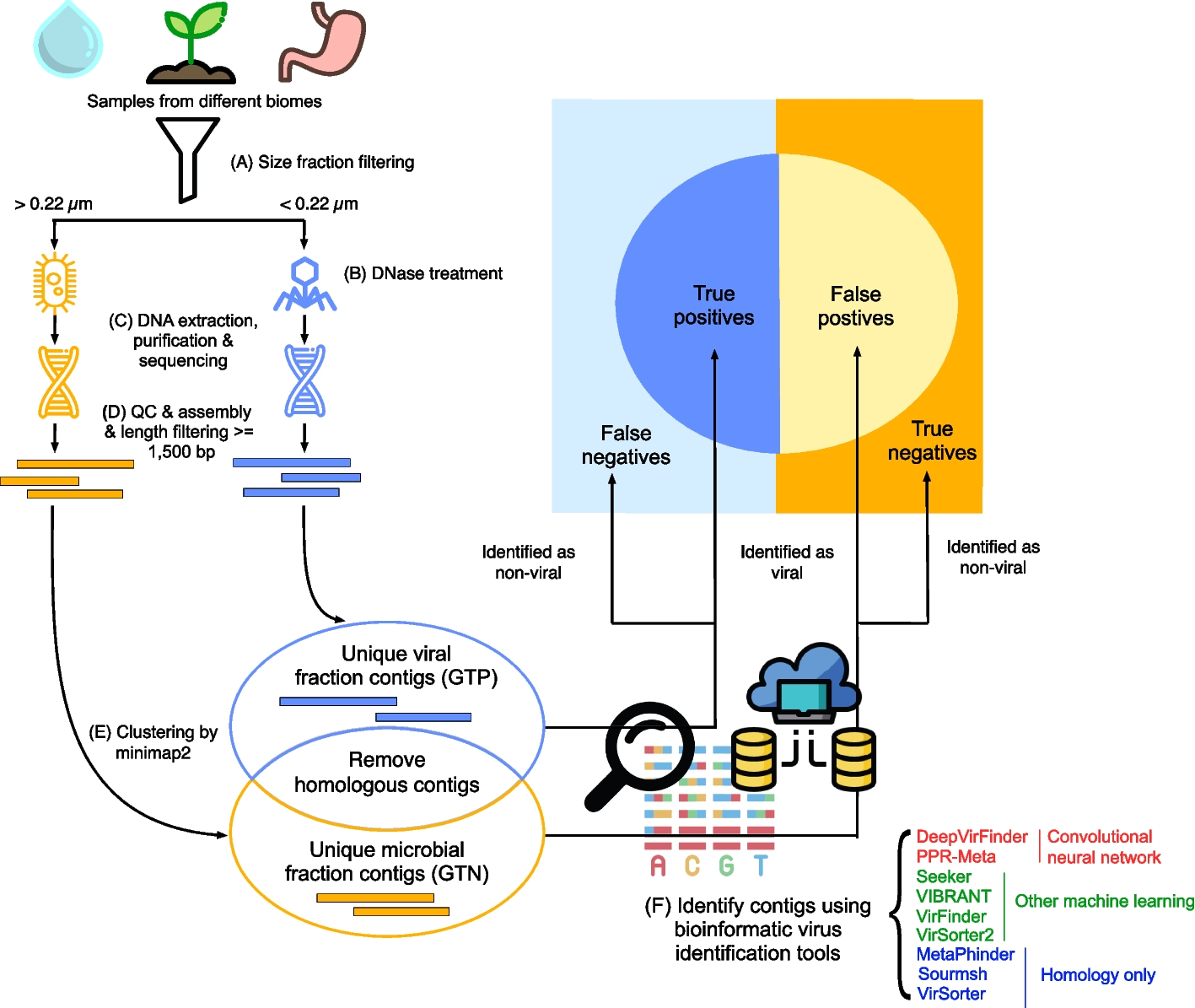




Previous studies have demonstrated that m6A reader proteins ECT2 and ECT3 bind largely overlapping targeted sites and ECT2/ECT3/ECT4 participate redundantly in certain plant developmental processes, including plant developmental timing, morphogenesis, and plant organogenesis [16, 28, 29]. To investigate the spatial aspect of their overlapping targets and redundant regulation, we first utilized the bimolecular fluorescence complementation (BiFC) system to co-express ECT2, ECT3, and ECT4 in pairs with split yellow fluorescent protein (YFP) in N. benthamiana leaves. All three protein pairs, ECT2-nYFP + ECT3-cYFP, ECT2-nYFP + ECT4-cYFP, and ECT3-nYFP + ECT4-cYFP, exhibited strong reconstituted YFP signal in the cytoplasm. In contrast, no fluorescence signal was observed with empty vector co-expression and other negative controls (Fig. 1a; Additional file 1: Fig. S1). To follow up on these results, we performed yeast two-hybrid (Y2H) assays to examine pairwise interactions among the full-length ECT2, ECT3, and ECT4 proteins. Yeast strains co-transformed with ECT2-BD + ECT3-AD, ECT2-BD + ECT4-AD, and ECT3-BD + ECT4-AD successfully grew on selective medium at all dilutions (Fig. 1b), demonstrating the occurrence of protein–protein interactions.
Fig. 1
Direct protein–protein interaction among ECT2, ECT3, and ECT4. a BiFC assay showing the physical associations among ECT2, ECT3, and ECT4 in Nicotiana benthamiana leaf cells. The association of paired proteins is indicated by YFP fluorescence in the cytoplasm. Scale bars = 40 μm. b Y2H assay showing the physical associations among ECT2, ECT3, and ECT4 in yeast cells. The full-length coding sequences of ECT2, ECT3, and ECT4 were fused with either the GAL4-AD or BD domain as indicated. YSD-Leu-Trp-Ade-His, selective medium without tryptophan, leucine, histidine, or adenine; YSD-Leu-Trp, medium without tryptophan or leucine (growth control). c Pull-down assay showing the interaction among GST-ECT2, MBP-ECT3, and SUMO-ECT4 in vitro. Purified MBP-ECT3 and SUMO-ECT4 proteins were incubated with GST-ECT2 or GST alone, and pull-down assays were performed using GST magnetic beads, followed by immunoblot analysis with anti-GST, anti-MBP, and anti-SUMO antibodies. d Correlation analysis of mRNA expression levels in Arabidopsis among ECT2, ECT3, and ECT4 in the ATTED-II database (n > 10,000 samples; ρ, Spearman’s correlation coefficient). P-values were calculated with Pearson’s correlation analysis. e The protein level of ECT2–FLAG in indicated samples, as determined by western blot. β-Actin protein was used as the loading control. f In vivo FA-RIP assay showing that ECT2-IP RNA amount and m6A level is enriched in ECT2:ECT2/ect2-1 compared with ECT2:ECT2/ect2/3/4 plant. IgG-IP was used as control. Data are presented as means ± SE, n = 3 biological replicates × 2 technical replicates. *P < 0.05, **P < 0.01 (two-sided t-test)
Since both the BiFC and Y2H assays confirmed that ECT2, ECT3, and ECT4 directly interact with each other, we asked whether ECT2, ECT3, and ECT4 could form a complex via pairwise interaction. To test this hypothesis, we performed an in vitro glutathione S-transferase (GST) pull-down assays with purified recombinant proteins from Escherichia coli to examine whether ECT2 could physically interact with ECT3 and ECT4. We found that maltose-binding protein (MBP)-tagged ECT3 and (small ubiquitin-like modifier) SUMO-tagged ECT4 interacted with GST-tagged ECT2, but not with GST alone (Fig. 1c), suggesting that ECT2, ECT3, and ECT4 constituted a complex through direct protein–protein interactions in vitro. Additionally, the mRNA expression level landscape of ECT2, ECT3, and ECT4 was strongly correlated in all three pairwise comparisons (Spearman’s ρ values between 0.69 and 0.76; Fig. 1d), indicating their largely undifferentiated functions in various plant biological and developmental processes.
To investigate the regulatory role of ECT2, ECT3, and ECT4 interaction in m6A-modified RNAs, we assessed whether the ECT2-ECT3-ECT4 interaction affects the m6A-binding activity of ECT2 by conducting a formaldehyde cross-linking and RNA immunoprecipitation (FA-RIP) assay using the generated ECT2 complementary transgenic plants in ect2-1 and ect2/3/4 background, respectively (ECT2:ECT2/ect2-1 and ECT2:ECT2/ect2/3/4). Both ECT2:ECT2/ect2-1 and ECT2:ECT2/ect2/3/4 plants expressed FLAG-tagged ECT2 proteins at similar levels (Fig. 1e). Our findings demonstrated a significant decrease in the amount of immunoprecipitated RNA and m6A level in ECT2:ECT2/ect2/3/4 compared to ECT2:ECT2/ect2-1 (Fig. 1f), indicating that ECT2, in collaboration with ECT3 and ECT4, can bind more m6A-modified RNA than ECT2 alone. These observations established that ECT2/ECT3/ECT4 can form a complex through direct protein–protein interactions, thereby enhancing m6A-binding capability and conferring fine regulation on their target RNAs.
ECT2/ECT3/ECT4 are required for seed germination and post-germination development under ABA treatmentAlthough the deficiency of ECT2/ECT3/ECT4 has been shown to delay development in early growth stages [16, 28], it is not yet known whether they play a role in abiotic stress responses. To investigate the biological functions of ECT2/ECT3/ECT4, we generated homozygous T-DNA insertion double mutants ect2-1/ect4-1 (referred to as ect2/4) and ect3-2/ect4-1 (ect3/4), as well as a triple mutant, ect2-1/ect3-2/ect4-1 (ect2/3/4), by crossing the ect2-1 (SALK_002225) mutant with the ect3-2 (GABIseq_487H12) and ect4-1 (SALK_151516) mutants (Additional file 1: Fig. S2a). Reverse transcription quantitative PCR (RT-qPCR) and detailed phenotypic analysis confirmed knockout of the target genes among ect2/4, ect3/4, and ect2/3/4 mutants and the ect2/3/4 mutant exhibited defective leaf morphology under normal growth conditions (Additional file 1: Fig. S2b, c), consistent with a previous report [16].
To investigate the roles of ECT2, ECT3, and ECT4 in abiotic stress responses, we initially assessed their ABA sensitivity by measuring the germination rate of single mutant seeds (ect2-1, ect3-2, and ect4-1). No obvious differences in germination rates were observed between the wild-type (WT) and mutant seeds under normal conditions (Mock) or with varying ABA concentrations (Additional file 1: Fig. S3a-c). As ABA is known to inhibit cotyledon greening more strongly than germination [30], we also evaluated the cotyledon greening rates and found ect2-1 exhibited a significant reduction in greening compared to WT in the presence of ABA. The ect3-2 and ect4-1 mutants showed only a slight reduction (not statistically significant) in cotyledon greening rates upon ABA treatment (Additional file 1: Fig. S3d). We next examined ABA sensitivity in double and triple mutant seeds (ect2/4, ect3/4, and ect2/3/4) and found that in the presence of different concentrations of ABA, all mutant seeds all exhibited enhanced ABA sensitivity compared to WT in a manner that demonstrated genetic redundancy; ABA hypersensitivity in ect3/4 was weaker than that of ect2/4, and ect2/3/4 seeds exhibited stronger ABA hypersensitivity than either ect2/4 or ect3/4 mutants (Fig. 2a–d). These results demonstrated that ECT2/ECT3/ECT4 redundantly and negatively regulate ABA signaling during seed germination and post-germination growth.
Fig. 2
ECT2/ECT3/ECT4 are required for seed germination and post-germination development under ABA treatment. a Phenotypic analysis of the ABA response in WT, ect2/3/4, ect3/4, and ect2/4 seeds grown on 1/2 MS-medium supplemented with 0 (Mock), 0.5, or 0.8 μM ABA under long-day conditions. Representative photographs were taken 8 days after cold stratification. b Statistical analysis of germination rates in WT, ect2/3/4, ect3/4, and ect2/4 plants under ABA treatment. Radicle emergence was used as the morphological marker for germination. At least 40 seeds per genotype were measured in each replicate. Biological triplicates were averaged. Data are presented as the mean ± SE. c–d Statistical analysis of germination rates 4 days after imbibition (c) and of cotyledon greening rates 8 days after imbibition (d) in WT, ect2/3/4, ect3/4, and ect2/4 plants under ABA treatment. Data are presented as the mean ± SE; n = 3 biological replicates. **P < 0.001, ***P < 0.001, ****P < 0.0001 (two-sided t-test). e Relative mRNA levels of COR47 and NCED3 in 7-day-old WT, ect2/3/4, ect3/4, and ect2/4 seedlings under Mock and ABA treatment. TUB8 was used as the internal control gene. Data are presented as the mean ± SE; n = 3 biological replicates × 2 technical replicates. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 (two-sided t-test)
In addition, we found that expression levels of some ABA-responsive genes were modulated by ECT2/ECT3/ECT4 activity. In the presence of exogenous ABA, the ect2/3/4, ect3/4, and ect2/4 mutants showed up-regulation of ABA-responsive genes such as COLD-REGULATED 47 (COR47) and NINE-CIS-EPOXYCAROTENOID DIOXYGENASE 3 (NCED3). Among the three double/triple mutants, these stress-responsive genes were most significantly up-regulated in ect2/3/4 compared to the WT (Fig. 2e). These results confirmed that ECT2/ECT3/ECT4 redundantly function in the expression of ABA-related transcripts during seed germination and post-germination growth.
Considering the observation that the absence of ECT2 protein exhibited the reduced cotyledon greening rate, and the presence of ECT2 protein could largely rescue the ABA sensitivity phenotype, we next assessed whether ECT2 predominately depends on its m6A-binding ability to function in the ABA response. We generated ECT2:ECT2/ect2/3/4 and ECT2:ECT2m/ect2/3/4 transgenic plants, which expressed the coding sequence of ECT2 and the m6A-binding abolished ECT2m with double mutations (W521A/W534A) [14] in the ect2/3/4 mutant background. The germination and cotyledon greening rates of the transgenic seeds were indistinguishable from WT seeds under Mock treatment (Additional file 1: Fig. S4a, b). However, in the presence of ABA, ECT2:ECT2/ect2/3/4 but not ECT2:ECT2m/ect2/3/4 can partially rescue the ABA hypersensitivity in the ect2/3/4 mutant (Additional file 1: Fig. S4). These results suggest that the m6A binding function plays a core regulatory role in ECT2/ECT3/ECT4-mediated ABA response.
ECT2/ECT3/ECT4 promote mRNA stabilizationOur previous studies have shown that ECT2 promotes mRNA stability in the cytoplasm [14] (Fig. 3a). However, it is unclear whether ECT2/ECT3/ECT4 cooperatively facilitate mRNA stabilization. To investigate this, we performed mRNA sequencing (mRNA-seq) in WT and ect2/3/4 seedlings. Correlation analysis between two biological replicates for each genotype confirmed that the replicability of the mRNA-seq (Additional file 1: Fig. S5). Transcripts with per million mapped fragments (FPKM) < 1 were excluded. Considering that more than 94% of ECT3 targets were overlapped with ECT2 targets [29], and the predominant phenotype-related regulatory role of ECT2 in ECT2/ECT3/ECT4 (Fig. 2a), we chose ECT2 targets from previous FA-CLIP-seq [14] that could largely cover the targets of ECT2, ECT3, and ECT4 to analyze the datasets. We divided the genes into three groups: ECT2 targets, ECT2 & m6A targets (ECT2-binding genes with m6A modification), and Non-targets (ECT2 unbound genes). Our results showed that the ect2/3/4 mutant had significantly lower accumulation of ECT2 targets and of ECT2 & m6A targets compared to non-targets (Fig. 3b). This trend of reduced transcript accumulation was stronger in the ect2/3/4 than in the ect2-1 mutant (Fig. 3a, b), consistent with the functional redundancy of ECT2/ECT3/ECT4.
Fig. 3
ECT2/ECT3/ECT4 promote stabilization of target m6A-modified mRNA. a–b Cumulative distribution of relative mRNA expression in ect2-1 compared to WT (a) and ect2/3/4 compared to WT (b) for Non-targets (black), ECT2 targets (blue), and ECT2 & m6A targets (red). c Cumulative distribution of mRNA half-life in WT for Non-targets (black), ECT2 targets (blue), and ECT2 & m6A targets (red). d Cumulative distribution of mRNA half-life in WT, analyzed based on the number of ECT2 target sites in each transcript. e Cumulative distribution of relative mRNA half-life log2 fold changes between ect2/3/4 and WT for Non-targets (black), ECT2 targets (blue), and ECT2 & m6A targets (red). f Cumulative distribution of mRNA half-life log2 fold changes between ect2/3/4 and WT, analyzed based on the number of ECT2 target sites in each transcript. P-values were calculated using two-sided Mann–Whitney U test
We subsequently performed mRNA stability profiling by calibrating External RNA Controls Consortium (ERCC) spike-in controls in equal amounts of total RNA from 7-day-old WT and ect2/3/4 mutant plants to investigate the functional role of ECT2/ECT3/ECT4 in mRNA stabilization. Plants were collected over a series of time points after transcription inhibition with actinomycin D. Analysis results revealed that both ECT2 targets and ECT2 & m6A targets tended to have longer mRNA half-lives than Non-targets in WT (Fig. 3c; Additional file 2: Table S1). We also examined whether there was a correlation between mRNA stability and the number of ECT2-binding sites by dividing the ECT2 targets into three groups based on the number of ECT2-binding sites they had. A positive association was observed between the number of ECT2-binding sites and mRNA target stability, with targets having more than two ECT2-binding sites showing increased stability compared to targets with only one or two binding sites (Fig. 3d). Moreover, we found that compared with WT, the mRNA half-lives of ECT2 and ECT2 & m6A targets were significantly shortened in the ect2/3/4 mutants relative to Non-targets (Fig. 3e). This analysis also revealed that mRNA stabilization was again associated with the number of ECT2-binding sites (Fig. 3f; Additional file 2: Table S1). We further examined individual genes and found that some ABA-related transcripts bound by ECT2 and modified with m6A were rapidly degraded in ect2/3/4 mutant (Additional file 1: Fig. S6), suggesting that ECT2/ECT3/ECT4 function in ABA response via enhancing mRNA stabilization.
Additionally, we analyzed the previously identified ECT2/ECT3 common targets [29] with our mRNA stability profiling data. Consistently, both ECT2/ECT3 common targets had longer mRNA half-lives than their Non-targets in WT (Additional file 1: Fig. S7a). Disruption of ECT2/ECT3/ECT4 reduced mRNA half-lives for ECT2/ECT3 common targets compared with Non-targets (Additional file 1: Fig. S7b). These findings provide compelling evidence that the m6A reader proteins ECT2, ECT3, and ECT4 act in concert to promote mRNA stabilization in Arabidopsis, highlighting a previously unrecognized regulatory mechanism for m6A modification in plant RNA metabolism.
ECT2/ECT3/ECT4 have no effects on alternative polyadenylation and translationOur previous findings suggested that ECT2 may be involved in mRNA 3′ end processing due to its binding around the UGUA region [14]. However, a recent study using NanoPARE analysis revealed that ECT2/ECT3/ECT4 do not play a direct role in alternative polyadenylation [29]. We further investigate whether ECT2/ECT3/ECT4 affect alternative polyadenylation (APA). The subcellular localization of m6A readers is known to influence their regulatory roles in RNA processing and metabolic processes. Thus, ect2-1 mutant plants that expressed a transgene for ECT2 fused to green fluorescence protein (eGFP) were utilized to acquired high-resolution images of ECT2-eGFP in 7-day-old root tips. The results confirmed the cytoplasmic localization of ECT2 (Additional file 1: Fig. S8), suggesting that ECT2 would have negligible effects on RNA processing in the nucleus. To investigate further, we sequenced polyadenylation (poly(A)) sites using the A-seq2 method [31] in WT and ect2/3/4 plant samples. Microheterogeneity at the cleavage and poly(A) site often produces clusters of related poly(A) sites [32]. We therefore consolidated all sites ending within 25 nucleotides of one another into a single poly(A) cluster (PAC) for further analysis and identified over 19,000 high-confidence PACs per sample (tags per million (TPM) ≥ 3) after mapping ~ 10 million reads per sample to the Arabidopsis genome. More than 80% of PACs aligned to the terminal exons and 3′ untranslated regions (3′ UTRs) of protein-coding genes (Additional file 1: Fig. S9a) and correlation analysis between biological replicates confirmed the reproducibility of the poly(A) site profiling results (Additional file 1: Fig. S9b).
We identified a total of 128 genes with high-confidence PAC shifts (P-value < 0.05, Fisher’s exact test) in the ect2/3/4 mutant, which only accounted for 1.34% of all genes with a detected PAC (Additional file 1: Fig. S10a, b). To examine whether ECT2/ECT3/ECT4 were associated with the 128 PAC-shifted genes, we calculated the percentage of PAC-shifted genes in the three groups described above: ECT2 targets, ECT2 & m6A targets, and Non-targets. The results showed that the PAC shifting rate was comparable between ECT2 targets, ECT2 & m6A targets, and Non-targets (Additional file 1: Fig. S10a, c). Because PAC shifting would affect 3′ UTR length, we compared 3′ UTR length between ect2/3/4 mutant and WT plants using the same mRNA groupings. The results showed that there were no significant differences in 3′ UTR length in ECT2 targets and ECT2 & m6A targets compared to Non-targets (two-sided t-test; Additional file 1: Fig. S10d). Thus, we conclude that ECT2/ECT3/ECT4 are not responsible for the APA processing of m6A-modified genes.
Considering the cytoplasmic localization of ECT2, we evaluated their impact on translation efficiency in WT seedlings where mRNAs were recognized by ECT2 or ECT2/ECT3/ECT4 and in ect2-1 or ect2/3/4 seedlings where mRNAs were not recognized by these m6A reader proteins. Ribosome profiling (ribo-seq) was performed to measure the translation efficiency (ribosome-bound fragments/mRNA input) of targeted mRNAs (Additional file 3: Table S2). Ribosome-bound fragments were generated with high reproducibility between biological replicates through nuclease digestion of polysomes into monosomes (Additional file 1: Fig. S11a-d). Our analysis showed no significant differences in translation efficiency between ECT2 or ECT2 & m6A targets and Non-targets in ect2-1, ect2/3/4, or WT plants (Additional file 1: Fig. S11e, f). This suggests that ECT2/ECT3/ECT4 have no function in protein translation.
The PrLDs of ECT2 directly interacts with PAB2 and PAB4The mechanism by which human m6A reader proteins guide and decide RNA fate is determined by their interacting proteins [33]. Thus, to investigate the molecular mechanism of ECT2/ECT3/ECT4-mediated mRNA stabilization, the priority task is to identify their binding proteins. Since ECT2 is the dominant functional protein among ECT2, ECT3, and ECT4, we perform formaldehyde cross-linking and ECT2 immunoprecipitation combined with mass spectroscopy (FA-IP/MS) analysis with and without RNase T1 treatment in ECT2:ECT2/ect2-1 plants. Note that RNase T1 was used to avoid the RNA-induced protein–protein interaction due to the RNA binding ability of ECT2 (Fig. 4a; Additional file 1: Fig. S12a; Additional file 4: Table S3). Our analysis revealed that poly(A) binding protein 2 (PAB2) and PAB4 were potential interacting proteins of ECT2, regardless of RNase T1 treatment (Fig. 4a; Additional file 1: Fig. S11a;). Additionally, ECT3 was also co-immunoprecipitated with ECT2, providing further evidence for their interaction (Fig. 4a; Additional file 1: Fig. S12a). As PAB family proteins have been shown to promote mRNA stabilization through binding to poly(A) tails in mammals [34,35,36], we hypothesized that PAB2 and PAB4 could be ECT2’s interacting proteins to facilitate the function of ECT2/ECT3/ECT4-mediated mRNA stabilization. To confirm this, we conducted BiFC and in vitro Y2H assays (Fig. 4b, c; Additional file 1: Fig. S12b, c) which demonstrated direct interactions between PAB proteins and ECT2. Moreover, a correlation analysis of mRNA expression levels revealed that ECT2 had strong co-expression with PAB2 and PAB4 in Arabidopsis (Spearman’s ρ values between 0.67 and 0.83; Fig. 4d). The plant.MAP database (http://plants.proteincomplexes.org) [37] also supports the stable interaction between ECT2 and PAB family proteins.
Fig. 4
ECT2/ECT3/ECT4 promote mRNA stability by recruiting PAB proteins. a Scatterplot showing the proteins bound to endogenous Arabidopsis ECT2 after RNase T1 treatment. The plot is based on the enrichment level (IP/control) and P-value. b BiFC assay showing the physical interaction between ECT2 and PAB2/PAB4 in Nicotiana benthamiana leaf cells. Scale bars = 20 μm. c Y2H assay showing the interaction of ECT2 with PAB2 and with PAB4 in yeast cells. d Correlation analysis of mRNA expression levels in Arabidopsis among ECT2 and PAB2, and PAB4 in the ATTED-II database (n > 10,000 samples; ρ, Spearman’s correlation coefficient). P-values were calculated with Pearson’s correlation analysis. e Top: predictions of PrLDs made by the “prion-like amino acid composition” (PLAAC; http://plaac.wi.mit.edu/); bottom: schematic diagram of ECT2 and its fragments. f Pull-down assay showing a direct interaction between PAB2 and PrLD domain of ECT2 in vitro. Purified MBP-PAB2 was incubated with GST-ECT2 fragments or GST alone, and pull-down assays were performed using GST magnetic beads, followed by immunoblot analysis with anti-GST and anti-MBP antibodies. g Overlapping of ECT2- and PAB2-binding targets in Arabidopsis. h The spatial distance distribution between the PAB2- and ECT2-binding sites. P-values were calculated using two-sided Mann–Whitney U test. i Cumulative distribution of relative mRNA half-life between ect2/3/4 and WT for Non-targets (black), ECT2 targets (blue), and ECT2 & PAB2 common targets (red). P-values were calculated using two-sided Mann–Whitney U test
To investigate the interaction domain of ECT2 with PAB2 and PAB4, we purified full-length ECT2, as well as four fragments (F1 to F4) containing one or two PrLD domains or YTH domain alone (Fig. 4e), each tagged with GST, and MBP-tagged PAB2 and PAB4 proteins, and conducted in vitro GST pull-down assays to assess the binding of each fragment to PAB2 or PAB4. We found that both PAB2 and PAB4 interacted with full-length ECT2 and each PrLD of ECT2, but not with YTH domain of ECT2 and GST alone (Fig. 4f; Additional file 1: Fig. S12d), suggesting that the PrLDs mediate the physical interaction of ECT2 with PAB proteins.
PAB2 and PAB4 promote mRNA stabilization in ArabidopsisAlthough PAB family proteins have been demonstrated to facilitate mRNA stabilization and translational efficiency through binding to poly(A) tails in mammalian [34, 36, 37], whether PAB2 and PAB4 stabilize mRNA has not been validated in plants [38]. Therefore, we analyzed published CLIP-seq data for PAB2 and PAB4 [38] and our mRNA stability profiling data in WT. We revealed that both PAB2 targets and PAB4 targets (thresholds: IP/Control ≥ 1, P-value < 0.05, FPKM value > 1) tended to have longer mRNA half-lives than their Non-targets, confirming the role of PAB2 and PAB4 in mRNA stabilization (Additional file 1: Fig. S13).
ECT2/ECT3/ECT4 coordinately enhance mRNA stability through recruitment of PAB2 and PAB4We identified overlapping binding targets of ECT2 and PAB proteins and have found that 61.6% (2268) of ECT2 targets are bound by PAB2 (Fig. 4g) and 50% of ECT2 targets are bound by PAB4 (Additional file 1: Fig. S14a). Further analysis of the spatial distance between their binding regions revealed that the binding sites of PAB2 and PAB4 were in the same region as the ECT2 binding positions (Fig. 4h; Additional file 1: Fig. S14b). By analyzing the mRNA lifetime accumulation between WT and ect2/3/4 plants, we found that in ect2/3/4 mutants, the mRNA half-lives of ECT2 & PAB2 common targets were significantly decreased compared to Non-targets (genes not targeted by either ECT2 or PAB2) (Fig. 4i), revealing a co-regulatory function of ECT2 and PAB2 in mRNA stabilization. A similar trend was also observed for ECT2 & PAB4 common targets (Additional file 1: Fig. S14c). Taken together, these results demonstrate the molecular mechanism by which ECT2 binds to m6A-modified mRNAs and promotes their stability by directly interacting with PAB2 and PAB4 proteins.
ECT2/ECT3/ECT4 function in multiple important biological pathwaysTo further investigate the functions of ECT2/ECT3/ECT4, we analyzed differentially expressed genes in the ect2/3/4 triple mutant compared with WT using our mRNA-seq data. There were 278 down-regulated and 186 up-regulated genes identified in ect2/3/4 (FPKM fold change ≥ 2 and P-value < 0.05; Additional file 5: Table S4). Gene Ontology (GO) analysis of the 464 differentially expressed genes revealed enrichment in biological processes including response to chitin, cold, wounding, bacterium and fungus, salt and oxidative stresses, abscisic acid, salicylic acid, auxin, and water deprivation (Additional file 1: Fig. S15), suggesting that ECT2/ECT3/ECT4 play regulatory roles in abiotic and biotic stress responses.
ECT2/ECT3/ECT4 stabilize ABA response-related genesWe then investigated the molecular mechanism underlying ABA hypersensitivity in the ect2/3/4 mutant. ECT2 can aggregate in cytoplasmic foci in response to heat and drought stresses [15, 16], which may influence its function. Therefore, we first characterized the subcellular localization of ECT2 under 50 μM ABA treatment using ECT2:ECT2-eGFP/ect2-1 transgenic plants. Confocal images of ECT2-eGFP in 7-day-old ECT2:ECT2-eGFP/ect2-1 root tips showed that ECT2 was still localized in the cytoplasm and did not aggregate in response to 50 μM ABA treatment (Additional file 1: Fig. S16). This indicated that the ECT2-mediated m6A-modified mRNA stabilization regulatory mechanism would not be altered upon ABA stimulation in this experiment. We speculated that ABA signaling-related genes could be modified with m6A modification and regulated by the ECT2/ECT3/ECT4-PAB2/PAB4-mediated mRNA stabilization pathway. To test this hypothesis, we selected four ABA signaling-related genes, namely DWD HYPERSENSITIVE TO ABA (DWA) 1, DWA2, SDIR1-INTERACTING PROTEIN1 (SDIRIP1), and CHAPERONIN 20 (CPN20), from the m6A-seq, ECT2-CLIP, PAB2-CLIP, and PAB4-CLIP sequencing results for subsequent mechanistic study. All of these genes are known negative regulators of ABA signaling, and mutants for the genes exhibit enhanced ABA responses such as delayed germination and post-germination development [39,40,41]. The ECT2-targeted sites at the 3′ UTR of these four genes were highly overlapping with m6A sites, PAB2 binding sites, and PAB4 binding sites (Fig. 5a).
Fig. 5
ECT2/ECT3/ECT4 stabilize the ABA response-related genes. a Integrative genomics viewer showing the indicated sequencing results on DWA1, DWA2, SDIRIP1, and CPN20 transcripts. The light blue box labeled in each sequencing result indicated the position of the m6A site, the binding sites of ECT2, PAB2, and PAB4. b m6A-IP-qPCR validation of the m6A peaks in DWA1, DWA2, SDIRIP1, and CPN20 under Mock and ABA treatment. Data are presented as means ± SE, n = 2 biological replicates × 2 technical replicates. c FA-RIP-qPCR validation of the binding ability of ECT2 towards DWA1, DWA2, SDIRIP1, and CPN20 in 12-day-old ECT2:ECT2/ect2-1 seedlings under Mock and ABA treatment. Data are presented as means ± SE, n = 3 biological replicates × 2 technical replicates. **P < 0.01, ***P < 0.001, ****P < 0.0001 (two-sided t-test). d Relative mRNA levels of DWA1, DWA2, SDIRIP1, and CPN20 in 7-day-old WT and ect2/3/4 seedlings under Mock and ABA treatment. TUB8 was used as the internal control gene. Data are presented as means ± SE, n = 3 biological replicates × 2 technical replicates. *P < 0.05, **P < 0.01 (two-sided t-test). e The mRNA lifetime of DWA1, DWA2, SDIRIP1, and CPN20 in 7-day-old WT and ect2/3/4 seedlings. TI, transcription inhibition. 18S was used as the internal control gene. Data are presented as means ± SE, n = 3 biological replicates × 2 technical replicates
We further performed m6A-IP-qPCR and FA-RIP-qPCR assays to examine whether DWA1, DWA2, SDIRIP1, or CPN20 transcripts were modified with m6A and whether they were bound by ECT2 under ABA treatment. The m6A-IP-qPCR results showed that compared with negative control, DWA1, DWA2, SDIRIP1, and CPN20 transcripts were consistently modified with m6A in 12-day-old control (Mock treatment) or ABA-treated WT seedlings (Fig. 5b; Additional file 1: Fig. S17a), consistent with previously published m6A sequencing results from ABA-treated plants [19] (Additional file 1: Fig. S17b). The FA-RIP-qPCR analysis in 12-day-old ECT2:ECT2/ect2
留言 (0)