
記住我



Although prior studies have demonstrated that α-syn, TDP-43 and tau can be shuttled to the lysosome for degradation [24-26], the specific proteases that carry out their degradation once inside the lysosomal compartment have not been fully characterized. Thus, we set out to broadly identify which lysosomal proteases are able to cleave α-syn, TDP-43 and tau via in vitro cleavage assays. Different cathepsins have distinct pH optima. Thus, to better capture the full range at which proteases can and cannot cleave these proteins we performed individual reactions of full-length, soluble α-syn, TDP-43 or tau with each protease across a broad range of pH settings (i.e., pH 3.4, 4.5, 5.5, and 7.4) (Fig. 1, Supplementary Data 1).
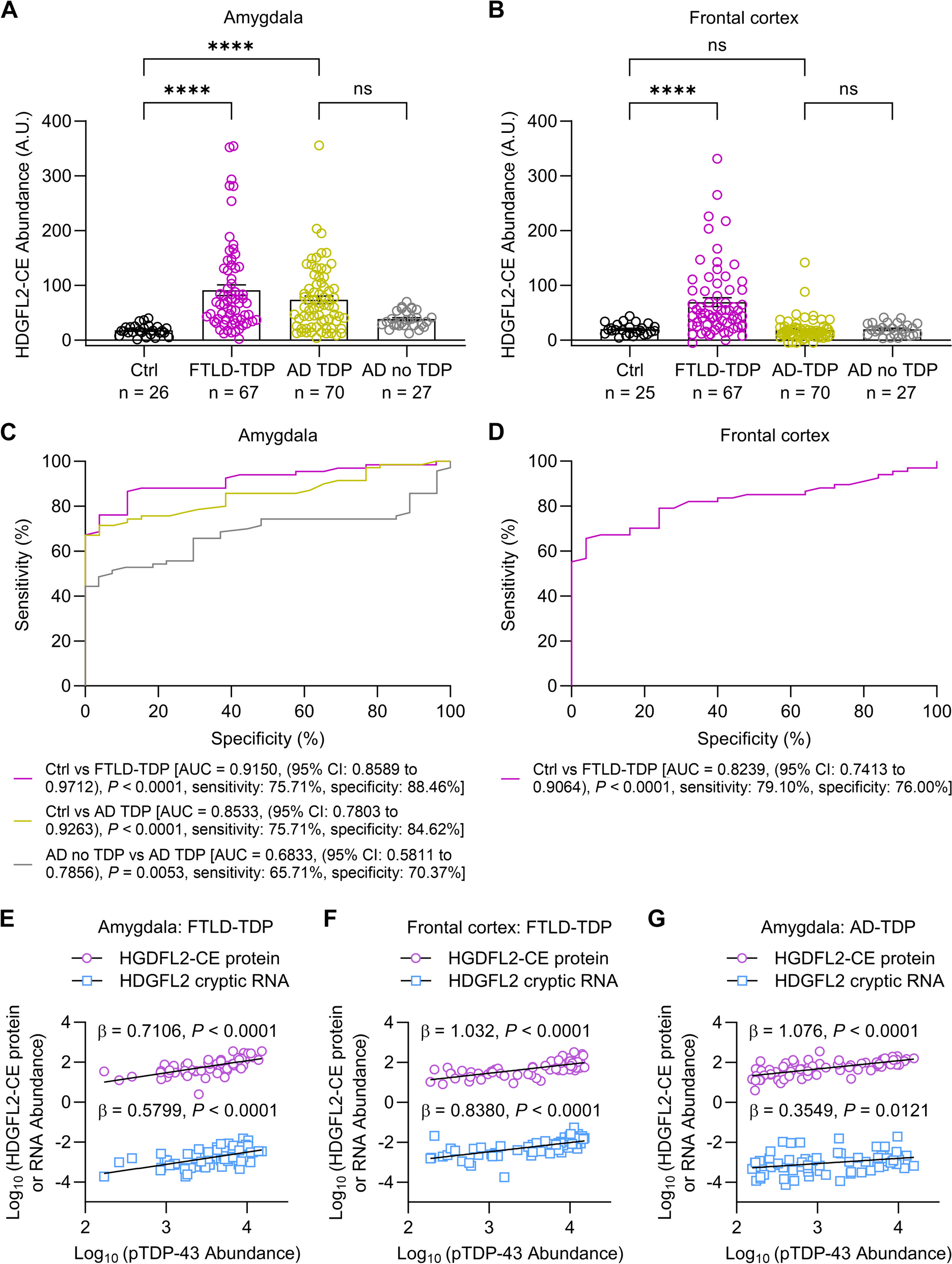
Fig. 1
Lysosomal proteases digest α-syn, TDP-43 and tau in a selective fashion. Recombinant, human, full-length α-syn, TDP-43 or 2N4R tau (1 µg) were incubated with 1 µM lysosomal protease for 1 h at the indicated pH. Reactions were subjected to SDS-PAGE and gels visualized by silver stain. Gels are representative of n = 3 independent replicates. Protein and protease combinations are organized according to those in which the protease can cleave (a) or cannot cleave (b) the protein. Full-length gels are shown in Supplementary Data 1 and control reactions for those proteases that cannot cleave any of the substrates are shown in Figure S1
We first assayed the largest class of lysosomal proteases, the cysteine protease family [39]. Under our experimental conditions, cathepsins B (CTSB), L (CTSL), K (CTSK), S (CTSS), and V (CTSV) were capable of cleaving all three full-length proteins at or near their optimal pH within the span of 1 h (Fig. 1a, Supplementary Data 1). Cathepsin C (CTSC) and asparagine endopeptidase (AEP) appeared to cleave α-syn and tau moderately but not TDP-43. Interestingly, cathepsin F (CTSF) could only cleave tau in this assay (Fig. 1a, b). Most cysteine proteases cleaved α-syn, TDP-43 and tau within their known pH optima (typically between pH 4.5 to 6.5). However, CTSV, an enzyme previously observed to cleave at lower pHs than other cysteine proteases [40], appeared most active against these full-length protein substrates at pH 3.4. Intriguingly, these results suggest that CTSV in vivo may only be active against these substrates in the context of mature, highly acidified lysosomes.
Next, we tested the serine proteases, cathepsin A (CTSA) and cathepsin G (CTSG). While CTSG was able to digest all three substrates at more neutral pHs, CTSA did not demonstrate any clear protease activity against any of our substrates in this assay (Fig. 1a, b, Supplementary Data 1). Lastly, we tested the lysosomal aspartyl proteases, cathepsin D (CTSD) and cathepsin E (CTSE). CTSD has been implicated in multiple neurodegenerative diseases, including a fatal congenital form of neuronal ceroid lipofuscinosis caused by CTSD mutations [41]. Consistent with their known pH optima, both CTSD and CTSE digested α-syn, TDP-43 and tau only at pH 3.4, the most acidic pH tested (Fig. 1a).
Although most of the enzymes tested are endopeptidases (i.e., enzymes that can cleave peptide bonds within the middle portions of a protein substrate), cathepsins A, C, H, and X are primarily exopeptidases, hydrolyzing mainly N- or C-terminal peptide bonds and exhibiting limited endopeptidase activity. Within the context of our assay, we noted that cathepsins A, H, and X (exopeptidases) as well as the endopeptidase cathepsin O (CTSO) did not demonstrate any detectable activity against α-syn, TDP-43 or tau (Fig. 1b). To confirm the proteolytic activity of these enzymes, however, we performed fluorescence-based cleavage assays with fluorogenically-tagged casein, confirming that all enzymes are active and can cleave this universal substrate (Fig. S1).
Multiplexed substrate profiling by mass spectrometry (MSP-MS) reveals comprehensive lysosomal protease cleavage maps of α-syn, TDP-43 and tauTo map the specific locations at which lysosomal proteases cleave α-syn, TDP-43 and tau, we took advantage of MSP-MS, a rapid, quantitative, reproducible and unbiased method of direct substrate cleavage site identification [31]. To aid in the identification of all possible cleavage sites, we designed a custom library of seven to eighteen-residue, overlapping peptides covering the entire sequences of α-syn, TDP-43 and tau (Table S1). In contrast to previous studies which analyzed protease activity on either full-length protein or protein fibrils [42,43,44,45], we specifically chose linear peptides to avoid the possibility of steric hindrance from protein secondary structure and to better model the denatured and ultimately partially cleaved state that most soluble proteins attain within the complex, acidic, protease-filled environment of lysosomes. Moreover, each peptide was designed to generate unique fragments for tandem mass spectrometry (MS/MS) detection. The library was then incubated with individual proteases at one or more pH setpoints within their optimal range and cleavage fragments assessed at two time points (Table S2). Cathepsins G, C and H were excluded due to reportedly low neuronal expression and because the latter two require a pre-activation step that is technically incompatible with MSP-MS [31, 46]. In total, the peptide library was tested against twelve recombinant proteases which generated 920 total cleavages across the three proteins. The results of this extensive mapping campaign are shown in Figs. 2a, 3a and 4a as well as in Supplementary Data 2.
Fig. 2
α-Syn proteolysis map and in vitro fluorescence protease assays. a A peptide library tiling across α-syn was incubated with individual lysosomal proteases (left). At various times and pHs, the reaction was subjected to MSP-MS to detect proteolytic cleavage sites in α-syn. The amino acid sequence of α-syn is in black letters at the top. Cleavage sites are indicated with the enzyme letter (e.g., B for CTSB) positioned at the P1 position (e.g. the B for CTSB is at amino acid position 7, so the cleavage occurs between positions 7 and 8). A total of 82 cleavages were found. Autosomal-dominant coding mutations associated with Parkinson’s Disease are noted in red above the α-syn sequence. Grey bars highlight amino acid mutations tested in in vitro fluorescent protease assays. b Pie chart demonstrating the number of cleavage sites within the α-syn sequence for each protease with the percentage of contributed cleavage sites in parentheses. c Table of maximal velocity (Vmax) ratios (mutant/WT), comparing protease cleavage of WT versus mutant α-syn peptides. A grey box denotes a mutation which was predicted to be "non-damaging." Mutations decreasing the Vmax of protease cleavage by 0–25% (1 point), 25–75% (2 points), and > 75% (3 points) are highlighted in light pink, dark pink, and dark red, respectively. Mutations augmenting the cleavage rate (-1 point) are highlighted in light green. Mutations with similar rate of cleavage compared to WT (0 points) are highlighted in yellow. Grey boxes denote no observed cleavage for either the WT or mutant peptide. Points were summed to derive a total “Damage Score”. d-i Representative curves of fluorescence generated from α-syn peptide cleavage, comparing WT and mutant peptides as labeled (n = 3 for all protease-substrate pairs). NAC, non-amyloid component domain; Æ, asparagine endopeptidase (AEP)
Fig. 3
TDP-43 proteolysis map and in vitro fluorescence protease assays. a A peptide library tiling across TDP-43 was incubated with individual lysosomal proteases (left). At various times and pHs, the reaction was subjected to MSP-MS to detect proteolytic cleavage sites in TDP-43. The amino acid sequence of TDP-43 is in black letters at the top. Cleavage sites are indicated with the enzyme letter (e.g., B for CTSB) positioned at the P1 position (e.g. the B for CTSB is at amino acid position 17, so the cleavage occurs between positions 17 and 18). A total of 553 cleavages were found. Autosomal-dominant coding mutations associated with amyotrophic lateral sclerosis are noted in red above the TDP-43 sequence. Grey bars highlight amino acid mutations tested in in vitro fluorescent protease assays. b Pie chart demonstrating the number of cleavages sites within the TDP-43 sequence for each protease with the percentage of contributed cleavage sites in parentheses. c Table of maximal velocity (Vmax) ratios (mutant/WT), comparing protease cleavage of WT versus mutant TDP-43 peptides. A grey box denotes a mutation which was predicted to be "non-damaging." Mutations decreasing the Vmax of protease cleavage by 0–25% (1 point), 25–75% (2 points), and > 75% (3 points) are highlighted in light pink, dark pink, and dark red, respectively. Mutations augmenting the cleavage rate (-1 point) are highlighted in light green. Mutations with similar rate of cleavage compared to WT (0 points) are highlighted in yellow. Grey boxes denote no observed cleavage for either the WT or mutant peptide. Points were summed to derive a total “damage score”. d-i Representative curves of fluorescence generated from TDP-43 peptide cleavage, comparing WT and mutant peptides as labeled (n = 3 for all protease-substrate pairs). NLS, nuclear localization sequence; RRM1 and RRM2, RNA recognition motifs 1 and 2; Æ, asparagine endopeptidase (AEP)
Fig. 4
Tau proteolysis map and in vitro fluorescence protease assays. a A peptide library tiling across tau was incubated with individual lysosomal proteases (left). At various times and pHs, the reaction was subjected to MSP-MS to detect proteolytic cleavage sites in tau. The amino acid sequence of tau is in black letters at the top. Cleavage sites are indicated with the enzyme letter (e.g., B for CTSB) positioned at the P1 position (e.g. the B for CTSB is at amino acid position 7, so the cleavage occurs between positions 7 and 8). A total of 285 cleavages were found. Autosomal-dominant coding mutations associated with frontotemporal dementia are noted in red above the tau sequence. Grey bars highlight amino acid mutations tested in in vitro fluorescent protease assays. b Pie chart demonstrating the number of cleavages sites within the tau sequence for each protease with the percentage of contributed cleavage sites in parentheses. c Table of maximal velocity (Vmax) ratios (mutant/WT), comparing protease cleavage of WT versus mutant tau peptides. A grey box denotes a mutation which was predicted to be "non-damaging." Mutations disrupting protease cleave by 0–25% (1 point), 25–75% (2 points), and > 75% (3 points) are highlighted in light pink, dark pink, and dark red, respectively. Mutations augmenting the rate cleavage (-1 point) are highlighted in light green. Mutations with similar rate of cleavage compared to WT (0 points) are highlighted in yellow. Grey boxes denote no observed cleavage for either the WT or mutant peptide. Points were summed to derive a total “damage score”. d-i Representative curves of fluorescence generated from tau peptide cleavage, comparing WT and mutant peptides as labeled (n = 3 for all protease-substrate pairs). N1 and N2, N-terminal repeats; P1 and P2, proline-rich regions, R1-4, microtubule binding repeats 1–4; Æ, asparagine endopeptidase (AEP)
α-SynAt 140 amino acids, α-syn is the smallest of the three proteins assessed. In total, there were 82 cleavages in α-syn by 9 of the 12 proteases tested (Fig. 2a). In agreement with the recombinant, full-length α-syn in vitro protease assay results (Fig. 1b), cathepsins A, F and O did not cleave any of the peptides within the library. Remarkably, CTSX, a protease that could not cleave full-length α-syn, was able to digest several α-syn peptides. This highlights the advantages of utilizing short peptides in this assay, since under physiological conditions secondary structure will eventually be resolved as individual lysosomal cathepsins will ultimately see peptides cleaved by other proteases. Protease cleavage sites were evenly distributed throughout the α-syn sequence, including within the non-amyloid component (NAC) domain which forms the core of α-syn fibrils [2]. Of the proteases tested, CTSL and CTSS displayed the most cleavage sites in the α-syn peptides with CTSB and CTSV following (Fig. 2b). Of the proteases examined, CTSD has previously been shown to cleave soluble α-syn [42] while cathepsins B, K, and L are capable of cleaving α-syn fibrils [43,44,45]. Our results confirm CTSD as a soluble α-syn protease and substantially supplement the known lysosomal proteases that can process α-syn to include five additional: AEP as well as cathepsins E, S, V, and X.
TDP-43TDP-43 contains 414 amino acids. Each of the 12 proteases tested was able to cleave TDP-43 peptides in multiple locations, resulting in 553 total cleavages (Fig. 3a). Cathepsins S, L, V and E had the greatest number of proteolytic sites (Fig. 3b). Full-length, recombinant TDP-43 in solution likely retains some secondary structure [46]. As such, it was notable that cathepsins A, F, O, X and AEP, all of which did not appear to degrade full-length TDP-43 in Fig. 1, were able to cleave linear TDP-43 peptides under these MSP-MS conditions. Prior experimental data has demonstrated cleavage sites within TDP-43 for CTSL (positions 32, 341) and CTSS (position 341) [27]. Our results validated these sites and provided many more additional CTSL and CTSS cleavage sites. Notably, protease processing sites were not uniformly distributed across TDP-43. One region replete with protease sites, amino acids 306–378, forms the densely packed common core of TDP-43 fibrils [46, 47]. In contrast, other regions within TDP-43 exhibited a relative paucity of cleavage sites, including within the carboxy (C)-terminus. Regions with a relative paucity of protease processing sites, such as the C-terminal glycine rich domain (amino acids 351 to 414) did contain AEP sites. This is consistent with prior reports that AEP exhibits relatively distinctive, non-overlapping cleavage recognition sequences compared to other proteases [33, 48].
TauThe longest adult isoform of tau (2N4R) contains 441 amino acids. Other than previously reported cleavage by CTSD and CTSS, little is known regarding lysosomal proteases that can digest tau [49, 50]. All twelve of the proteases tested cleaved tau peptides for a total of 285 cleavages (Fig. 4a). Similar to TDP-43, cathepsins A, O and X, which did not cleave full-length recombinant tau, exhibited a few sparse cleavages of tau peptides. CTSS and CTSL displayed the highest numbers of tau cleavage sites with cathepsins B, V, E and AEP following (Fig. 4b). Also like TDP-43, protease cleavage sites within tau clustered together. Portions of tau, encompassing the proline rich P1 and P2 domains, contained relative few proteolytic sites, with residues 131 to 170 demonstrating no cleavage sites in this assay. The region from residues 306 to 355 in tau makes up the structural core of paired helical and straight filaments found in AD [4]. Again in alignment with TDP-43, this region was relatively rich in cathepsin cleavage sites.
Proteases exhibit distinctive abilities to process α-syn, TDP-43 and tauHaving mapped the lysosomal proteolytic cleavage sites in α-syn, TDP-43 and tau, we next wondered if any of the proteases exhibited preferences for one of these substrates over the others. This prospect was of significant interest as cathepsins exhibit age-dependent, brain-region and cell-type specific variability in expression and activity [51,52,53]. Thus, preferential behavior of certain lysosomal proteases towards α-syn, TDP-43 or tau could contribute to selective neuronal vulnerability observed in different pathologies (e.g., PD versus ALS). Therefore, we performed comparative analysis of the proteases in regards to proteolysis of α-syn, TDP-43 and tau. To do so in proteins of different sizes, we normalized the number of cathepsin cleavages for each substrate relative to their amino acid number. Among these proteases, we found that two cysteine cathepsins, CTSL and CTSS, demonstrated the highest proportion of cleavage sites in α-syn, TDP-43 and tau (Fig. 5a). Cathepsins B, E, V and AEP also exhibited significant activity with numerous sites of cleavage identified. Comparing the aspartyl proteases, CTSE exhibited significantly more cleavage sites than CTSD. Notably, cathepsins A, F and O demonstrated relatively few sites of cleavage within tau and TDP-43 and no cleavage sites within α-syn.
Fig. 5
Lysosomal proteases exhibit distinctive abilities to process α-syn, TDP-43 and tau. a Comparison of the number of cleavage sites within α-syn, TDP-43 and tau for each lysosomal protease relative to total number of cleavage sites. b Hierarchical clustering analysis demonstrating the relative affinity of proteases for α-syn, TDP-43 or tau. Clusters identified are designated to the right. c-e Pairwise correlation analyses with significance values (p-values) of unique versus redundant activity between lysosomal proteases for α-syn (C), TDP-43 (D) and tau (E). A positive score suggests more correlation and a negative score lower correlation between protease cleavage sites. f-q The iceLogo output for each of the serine (F), aspartyl (G-H) and cysteine (I-Q) proteases demonstrating the frequency of particular amino acids at the P4 – P4’ positions of each protease recognition motif within α-syn, TDP-43 and tau. Amino acids that were more frequently seen are above the horizontal axis and those that were less frequently seen are below the horizontal axis. The cleavage site is indicated with a vertical hatched line. *p < 0.05, **p < 0.01, or ***p < 0.001
Co-pathologies are often found in neurodegenerative diseases, especially with increasing age, thus similarities or differences between cathepsin substrate preferences may be important in this process [54]. To assess which proteases are more or less alike in their ability to cleave α-syn, TDP-43 and tau, we performed hierarchical clustering [55]. This clustering analysis revealed up to six distinct clusters of cathepsins based on their relative preferences towards the three substrates (Fig. 5b). The first two clusters consisted of cathepsins A, F and O, which could not cleave α-syn. Cathepsins X, E and V formed a cluster that exhibit relatively even distributions of cleavages within α-syn, TDP-43 and tau. A cluster including cathepsins L, B and S had a slight preference for α-syn cleavage while in contrast, CTSK and AEP favored TDP-43 and tau over α-syn. Lastly, cathepsins A and D rather heavily favored TDP-43, and CTSF preferred tau. This knowledge would be important if these proteins were to be targeted for enhanced degradation through cathepsin upregulation.
Because little is known about redundant versus non-redundant activity among these proteases, we also searched for similarities among cathepsins within the cleavage site profile of each substrate. We performed this survey since it would be highly relevant if upregulating protease activity were to become potential treatments in neurodegeneration. To this end, we performed pairwise correlational analyses. This analysis revealed that across the three substrates, the cleavage profiles of the aspartyl protease cathepsin E and the cysteine proteases cathepsins L and V were positively correlated with one another (Fig. 5c-e). Cathepsin X was also positively correlated with CTSA and CTSO for TDP-43 and tau. Somewhat surprisingly, CTSB showed either minimal or a negative correlation with most other cathepsins in α-syn and tau. This is fascinating as CTSB has previously been shown to cleave similar sequence motifs that of other cysteine proteases, such as CTSL and CTSS, when profiled with more generalized, screening peptide libraries [29]. In contrast to CTSB, however, AEP demonstrated an expectedly negative correlation pattern with the other tested cathepsins, as AEP is known to be highly specific for the endoproteolytic cleavage of asparagine while most other proteases do not favor asparagines at the P2 through P2' positions [26]. Overall, this analysis suggests that certain cathepsins could potentially work redundantly to degrade α-syn, TDP-43 or tau. In contrast, other cathepsins, such as CTSB and AEP, cleave relatively unique regions in these proteins. Thus, if CTSB or AEP activity were lost, proteolytic cleavage in α-syn, TDP-43 and tau may be affected in a manner that is both similar and not easily compensated by other proteases. Intriguingly, this finding raises the possibility that the molecular basis for disease co-pathology (e.g., findings of synucleinopathy plus tauopathy in the same brain regions) could be due to the early or preferential loss of CTSB or AEP activity.”
A subset of neurodegenerative disease-associated mutations is predicted to disrupt lysosomal protease cleavageWhile individual proteases have preferred amino acid recognition sequences, certain positions within their recognition sequence tend to exert outsized importance. Protease cleavage sequences are expressed based on the positions relative to the cleavage site, with (from left to right) P4 to P1 found on the amino (N)-terminal side of the cleavage site and P1’ to P4’ found on the C-terminal side. When expressed in this fashion, many of the cysteine proteases (e.g., cathepsins L and V) prefer hydrophobic residues at P2 [56] while AEP prefers asparagine at P1 [57]. Serine proteases rely heavily upon the P1 position for recognition [58]. Aspartyl proteases like CTSD and CTSE strongly prefer hydrophobic residues at P1 and P1’ [27].
Having generated cathepsin cleavage maps for α-syn, TDP-43 and tau, we overlaid the known disease-associated variants onto the maps (Figs. 2a, 3a and 4a). In several cases, disease-associated mutations appeared in close proximity to protease cleavage sites and were predicted to alter amino acids important for protease recognition [59]. For example, the TDP-43 A315T mutation is positioned at P1’ for a CTSE cleavage site and is predicted to make cleavage less favorable, since threonine is polar and not hydrophobic like alanine is at this position (Fig. 3a). Similarly, the N279K mutation on tau removes a critical asparagine from a P1 cleavage site position for AEP (Fig. 4a). We thus asked if neurodegenerative disease-associated mutations could directly alter the efficiency of proteolytic cleavage by lysosomal proteases.
Since it was not feasible to test every mutation against every protease, we utilized two approaches to identify a subset of mutations mostly likely to abrogate cathepsin cleavage. First, we performed in silico comparisons of WT versus mutant protein sequences in the PROSPER database (https://prosper.erc.monash.edu.au/) [28]. This analysis generated potential cleavage sites as well as a rank order set of mutations predicted to change proteolytic cleavage (Fig. S2, Table S3). Second, we used the MSP-MS data to assess whether specific amino acids are preferentially recognized by certain proteases, specifically within the sequences of α-syn, TDP-43 and tau. To do so, we input the MSP-MS cleavage sites into the iceLogo algorithm (iomics.ugent.be/icelogoserver), which uses probability theory to identify conserved patterns in proteins [32]. The iceLogo output allowed visualization of favorable and unfavorable amino acids at each of the protease recognition motif positions (P4 to P4’) (Fig. 5f-q). Results of this analysis suggested that the amino acid preferences in α-syn, TDP-43 and tau for several of the proteases, specifically cathepsins E, L, S and AEP, were similar to previously published iceLogo maps [29, 31, 60] (Fig. 5i, l, n and q). However, CTSB showed sequence preferences within α-syn, TDP-43 and tau that diverged from current notations [29] (Fig. 5g).
From the PROSPER and iceLogo analyses, we curated a set of eleven potentially “damaging” disease mutations to test (three in α-syn, four in TDP-43 and three in tau) to test for altered proteolytic cleavage activity as well as non-damaging “control” mutations with lower probability of altering protease cleavage activity (Table S3). We then generated WT and mutant peptides encompassing these regions (Table S4). Each peptide was labelled with a coumarin-based (MCA) fluorophore and a fluorescence quencher on the N- and C-termini, respectively. The peptide substrates were thus quenched at baseline and only revealed fluorescence upon cleavage by a protease. We incubated the peptides with individual proteases and monitored the fluorescence generated over time. Given the nature of the assay, we limited our testing to the primary endopeptidases, cathepsins B, D, E, F, K, L, S and V as well as AEP.
Using this activity assay, the maximal velocities (Vmax) at which each enzyme cleaved the WT and mutant peptides were calculated (Fig. S3). Using these values, the rates of WT versus mutant cleavage were compared and ratios (mutant over WT) were generated. In this type of in vitro protease approach, every cathepsin would not be expected to cleave every peptide. Indeed, we found that each WT peptide was cleaved by three to seven of the nine proteases tested (Figs. 2c, 3c, 4c and Fig. S3). Mutant peptides were oftentimes cleaved similarly to wild-type peptides. However, in other cases they were cleaved more slowly or not at all. Rarely, mutant peptides were also cleaved more quickly than wild type. Examples for α-syn, TDP-43 and tau are described below and shown in Figs. 2c, 3c and 4c.
To better quantify the summed impact of these changes over many different proteases, we assigned “damage points” for each instance in which a mutation decreased a protease’s Vmax for peptide cleavage by 0–25% (1 point), 25–75% (2 points) or greater than 75% (3 points). In contrast, mutations that increased the rate of cleavage of mutant peptides were given a score of -1 point. By summing the point values, we were able to calculate a “Damage Score” for each mutation (Figs. 2c, 3c and 4c). Fascinatingly, each of the mutations predicted to be disruptive demonstrated positive scores of 3 to 6 while the control mutations (those disease mutations not predicted to be damaging to proteolytic cleavage) exhibited neutral or negative scores.
For α-syn, in silico predictions suggested that G51D and A53T would be damaging to proteolytic cleavage while A30P and E46K would not be. When α-syn G51D was tested via this in vitro activity assay, the cysteine proteases CTSB, CTSL, CTSK and CTSV cleaved the mutant α-syn G51D peptide less efficiently than the WT peptide for a total Damage Score of + 7 (Fig. 2c-e). The A53T mutation also diminished lysosomal protease cleavage by CTSK, CTSL and CTSE, resulting in a Damage Score of + 3 (Fig. 2c, f and g). The α-syn A30P and E46K mutations, which in silico analysis suggested would not be damaging, were also tested in vitro. Every protease capable of cleaving the wild-type peptides were also able to cleave the mutant peptides at similar or faster rates, with the single exception of CTSB cleavage of the A30P peptide (Fig. 2c, h and i). Overall, this analysis revealed that the A30P and E46K α-syn mutations had negative Damage Scores (of -3 and -2, respectively).
Disease-associated mutations in TDP-43 also affected proteolytic cleavage. TDP-43 A315T is in proximity to many protease cleavage sites (i.e., cathepsins A, B, L, S and V
留言 (0)