
記住我
Organ fibrosis is the final common pathological pathway of many chronic diseases and is characterized by various injuries and insults with maladaptive tissue repair, followed by excessive collagen deposition into the extracellular matrix (ECM). Structural alterations lead to tissue stiffness, vascular rarefaction, hypoxia, proliferation, differentiation and activation of fibroblasts and an inflammatory cascade that reinforces a vicious cycle of tissue destruction. Fibrosis can affect virtually every organ system, including the liver, heart, kidney, pulmonary system, and skin. The combined annual incidence of major fibrosis-related diseases is approximately 4968 per 100,000 person-years and has been estimated to contribute to 45% of annual deaths worldwide [1]. Despite imposing an enormous socioeconomic burden globally, there is a lack of available treatments to mitigate, halt or reverse fibrotic progression. Hence, it is imperative to explore the mechanisms underlying organ fibrosis and find potential new targets for antifibrotic therapies. Circular RNAs (circRNAs) are a large class of noncoding RNA molecules identified over the last decade that have recently been found to exert their pathophysiological roles through multidimensional mechanisms, including at the transcriptional, epigenetic, translational, and posttranslational levels [2]. In particular, circRNAs play critical roles in the development and progression of fibrosis [3], and manipulating circRNAs has shown promising prospects as ideal biomarkers of fibrosis and antagonists of fibrotic disorders in vivo. In this review, we provide an update on the versatile mechanisms linking circRNAs to fibrosis in different organs, and we uncover the potential of circRNAs as therapeutic targets and biomarkers.
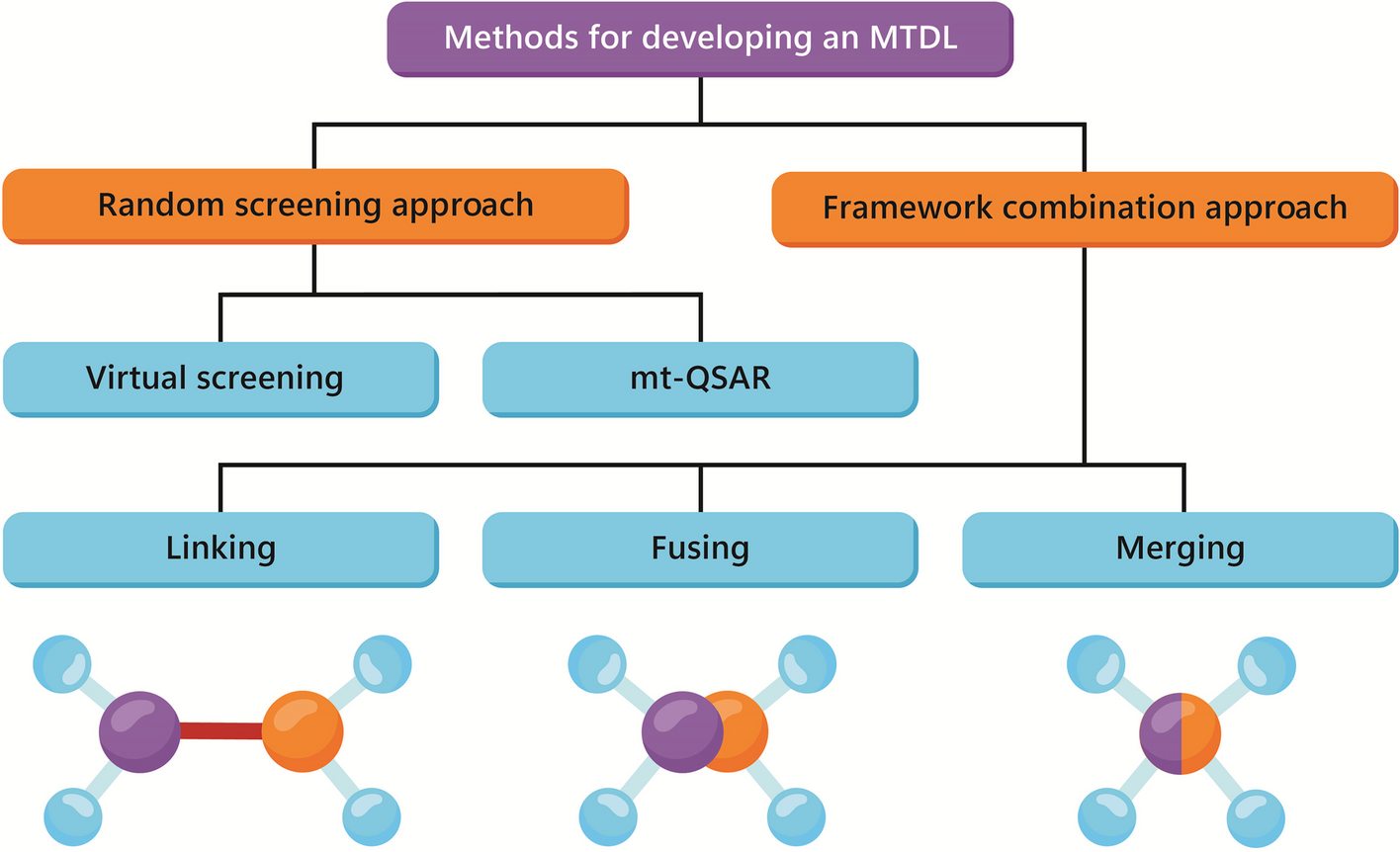
Properties of circRNAsSanger et al. first isolated and termed circRNAs in plant viruses and animal hepatitis almost half a century ago. However, these newly discovered circRNAs were simply associated with ‘scrambled exons’ or ‘mis-splicing’ until 2013, when two articles published in Nature [4, 5] identified the sponge-like function of microRNAs (miRNAs). Since then, circRNAs have become a research hotspot. CircRNAs are characterized as peculiar, single-stranded covalently closed RNA loops that spatially form 3D structures and interact with components, including RNA-binding proteins (RBPs), DNA or RNA. They are without 5’-caps and 3’-poly(A) tails and are thus resistant to exonuclease degradation and quite stable relative to their linear cognates. CircRNAs exert functions independently of their parental transcripts, and they are spatial-temporal, tissue- or cell-, and developmental stage-specific. Based on their origins and circularization patterns, circRNAs are categorized into at least seven types: intronic circRNAs (ciRNAs), exon‒intron circRNAs (EIciRNAs), exon circRNAs, tRNA intronic circular RNAs, mitochondrial circRNAs (mecciRNAs), read-through circRNAs (from exons between neighboring genes on the same strand), and fused circRNAs (fused exons between two remote genes) (Figs. 1) [2]. EIciRNAs and ciRNAs are sequestered in the nucleus, exon circRNAs are mostly exported to the cytoplasm, fused circRNAs are localized in the nucleus and cytoplasm, and mecciRNAs are located in the mitochondria and cytoplasm [6]. Localization is closely linked to function. ciRNAs and EIciRNAs regulate transcription via interaction with the U1 small nuclear ribonucleoprotein (snRNP). Fused circRNAs formed by cancer-associated chromosomal translocations are usually oncogenes involved in tumorigenesis. mecciRNAs regulate reactive oxygen species (ROS) in mitochondria. The function of read-through circRNAs is not yet well understood. Exon circRNAs act as sponges for proteins or miRNAs, scaffolds or templates for translation, thus playing more diverse roles than other types.
Fig. 1
Illustrations of different categories of circRNA. ①EcircRNA is formed by the back-splicing of exons; ②EIciRNA is circularized with introns “retained” between exons; ③ciRNA is formed by the back-splicing of the intron; ④rt-circRNA is a hybrid circle that includes coding exons from two adjacent and similarly oriented genes. ⑤f-circRNA is generated from transcribed exons of distinct genes affected by translocations. ⑥TricRNA is derived from pretRNA, which is recognized and cleaved by TSEN and produces 5’-OH and a 2’,3’-cyclic phosphate at each site of cleavage. Ligation of these two cleavage sites generates a circle. ⑦mecci-RNA refers to mtDNA-encoded or mitochondria-located circRNAs. SA splicing acceptor, SD splicing donor, EcirRNA exon circRNA, EIciRNA exon‒intron circRNA, ciRNA intronic circRNA, rt-circRNA read-through circRNA, f-circRNA fused circRNA, tricRNA tRNA intronic circular RNA, mecciRNA mitochondrial circRNA
Biogenesis, transportation and degradationSpliceosomes, cis-complementary sequences, epigenetic modulators and proteins are all involved in the biogenesis of circRNAs. There are three proposed mechanisms, direct back-splicing or intronic complementary sequence-driven circularization, RBP-mediated circularization, or lariat-mediated back-splicing, to decipher how circRNAs are alternatively spliced (Fig. 2, left panel) [7]. The complementary base pairing of inverted repeats flanking the exons mediates direct back-splicing. ALU repeats are the main pattern of circularization. Reverse complementary matches enriched in introns promote the formation of hairpin structures in transcripts, thus facilitating direct joining of the 5’ and 3’ splice sites. RBPs can inhibit or promote the biogenesis of circRNAs by binding specific motifs in the flanking introns, and their regulation of circRNAs is closely related to the tissue, cellular or developmental specificities of circRNAs [8]. Quaking, muscleblind, heterogeneous nuclear ribonucleoprotein L, fused-in-sarcoma, RNA-binding motif protein 20, nuclear factor 90 and nuclear factor 110 are RBPs that commonly facilitate circRNA production. DEAH-box helicase 9 and adenosine deaminase acting on RNA 1 have been reported to repress circRNA biogenesis by binding inverted repeat Alus in flanking sequences or A-to-I RNA editing activity [9]. A lariat structure is produced by exon-skipping during pre-mRNA processing, which can then undergo internal splicing to form exon circRNAs or EIcircRNAs. In addition, increased N6-methyladenosine modification (m6A) levels are associated with the production of circRNAs [10]. Writers (methyltransferases), erasers (demethylases), or readers (RBPs) modulate the circularization of m6A exons [11, 12].
Fig. 2
The biogenesis, transportation, degradation and functions of circRNA. The left panel describes the three mechanisms of biogenesis of circRNAs, including complementary base-pairing driven, RBP-driven and lariat-driven circularization. The middle panel illustrates the transportation and degradation of circRNAs. The right panel briefly summarizes the modes of action of circRNAs. DDX39A DEAD-box protein 39 A, EPO4 Exportin 4, G3BP1 Ras-GTPase-activating SH3 domain-binding-proteins 1, UPF1 Upframeshift 1, RISC RNA-induced silencing complex, Ago Argonaute, TRBP Transactivation response (TAR) RNA-binding protein, YTHDF YTH domain family, HRSP12 heat responsive protein 12, RBP RNA binding protein
The specific mechanism of circRNA transportation from the nucleus into the cytoplasm is not completely understood (Fig. 2, middle panel). The ATP-dependent RNA helicase DEAD-box protein 39 A is required for efficient nuclear export of long circRNAs, whereas DEAD-box protein 39 B controls the localization of short circRNAs [13]. Recently, Chen et al. reported that exportin 4 deficiency leads to nuclear accumulation of exon circRNAs, deleterious formation of RNA:DNA hybrids and DNA damage [14]. It has been demonstrated that m6A modification also plays pivotal roles in the nucleus–cytoplasm output of circRNAs [15].
CircRNAs are highly stable due to their resistance to exonucleases; thus, the mechanisms for their degradation remain to be fully elucidated (Fig. 2, middle panel). The binding of miRNAs to circRNAs can initiate Argonaute 2 (Ago2)-mediated circRNA degradation executed by the RNA-induced silencing complex (RISC). m6A modifications have been shown to cause widespread and diverse modulations of circRNA turnover. A subset of circRNAs containing m6A was identified as degraded by the YTHDF2-HRSP12-RNase P/MRP axes [16]. Structure-mediated RNA decay models formed by base pairing in circRNAs require the RBP UPF1 and its associated protein G3BP1, and downregulation of either UPF1 or G3BP1 causes the accumulation of circRNAs [17]. Furthermore, circRNAs can also be globally degraded by activated RNase L upon viral infection, which is required for PKR activation [18].
Regulatory roles of circRNAscircRNAs play essential roles in various pathophysiological conditions via interactions with numerous molecules at different levels. Their modes of action have recently been systemically reviewed elsewhere [7, 19, 20] and can be briefly summarized into five categories (Fig. 2, right panel): (A) miRNA sponges or decoys; (B) interactions with proteins; (C) regulation of transcription; (D) translation into proteins and peptides; and (E) mediation of cell‒cell communications (exosomal circRNAs). The miRNA sponge role is the most extensively investigated mechanism, but major pitfalls and controversies exist in this research field [21]. Major issues include being based solely on bioinformatics, a lack of fundamental validation, expression at relatively low levels, mismatched stoichiometry between circRNAs and miRNAs, and the harboring of a limited binding region. In contrast to other action patterns, the interactions between circRNAs and proteins are much more intricate [22]. circRNAs can change protein interaction modes, tether or sequester proteins, recruit proteins to chromatin, form circRNA–protein‒mRNA ternary complexes, and translocate or redistribute proteins. Interactions with proteins are the chief executors of life processes, and these interactions endow circRNAs with flexible and crucial functions. circRNAs regulate transcription at the initiation and elongation steps. They can serve as cis- or trans-acting elements and promote RNA Pol II transcription by interacting with U1 snRNP. In addition, circRNAs can form R loops with their cognate gene [23] or coactivate transcription factors to modulate transcription [24]. CircRNAs were previously thought to be noncoding RNAs due to their lack of a 5’ cap and 3’ poly (A) tail structure, which are necessary for cap-dependent translation. The identification of cap-independent mechanisms has revolutionized the concept that circRNAs are actually able to function as mRNAs to direct protein synthesis into peptides or proteins. Cap-independent translation can be mediated by the internal ribosome entry site-initiated pattern, m6A internal ribosome entry site-initiated patterns, and rolling translation [6]. By adopting a cell-based reporter system and using mass spectrum techniques, Fan et al. identified hundreds of circRNA-coding peptides, providing robust evidence of the coding capacity of circRNAs [25]. The new isoforms formed by noncanonical translation of circRNAs are different from their parental genes, thus opening a broad array of actions that can potentially extend well beyond mRNA-coding proteins [26].
Finally, as discussed in more detail below, circRNAs are cargoes packaged into exosomes that can mediate crosstalk involving numerous cell and organ systems [27], and exosomal circRNAs are promising biomarkers and therapeutic targets [28].
CircRNAs are pivotal regulators involved in the molecular pathways of organ fibrosisFibrosis is a multistaged and elaborate process involving numerous cells and extra or intracellular or soluble signaling molecules (Fig. 3). The fibrotic signaling pathways are not independent of each other but communicate in an orchestrated fashion, jointly amplifying the fibrotic cascades. Recently, circRNAs have been demonstrated to be pivotal regulators involved in complicated fibrotic signaling pathways, highlighting their potential as therapeutic targets.
Fig. 3
Representative circRNAs involved in core and regulatory fibrotic signaling pathways. Fibrotic signaling includes the TGF-β1, Wnt, Notch and PDGF pathways. The blue dotted line and a plus sign represent promoted effects, whereas a red dotted line and a minus sign represent inhibited effects. EMT Epithelial-mesenchymal transition, ECM Extracellular matrix
Transforming growth factor β (TGF-β) signalingTGF-β, a pluripotent growth factor, has been demonstrated to be the master regulator of fibrosis and orchestrates core and regulatory fibrotic signaling pathways. Therapeutic agents, including oligonucleotides, peptides, receptor blockers, antibodies, and small-molecule inhibitors, target the TGF-β network by inhibiting TGF-β synthesis, activation, binding to TGF-β receptors, TGF-βR function, or downstream molecules. However, these existing strategies have little impact on halting or suppressing fibrosis. Upon activation from its latent form, TGF-β binds to TGF-βRII, which subsequently dimerizes with and phosphorylates TGF-βRI. The downstream pathways of TGF-β include canonical solvated metal atom dispersed (smad)-dependent signaling and smad-independent noncanonical pathways (mitogen-activated protein kinase/extracellular signal-regulated kinase, p38 and JUN amino-terminal kinase, or phosphatidylinositol 3-kinase (PI3K/AKT)) [29]. CircANAPC7 functions as an anti-oncogene in pancreatic cancer through the miR-373/PH domain leucine-rich repeat-containing protein phosphatase 2 axis, leading to AKT dephosphorylation and TGF-β downregulation [30]. circPTEN1 binds to the MH2 domain of Smad4 to disrupt its physical interaction with Smad2/3 and blocks TGF-β signaling activation [31]. Overexpression of circNCOR1 epigenetically promotes Smad7 transcription, thereby inhibiting the TGF-β-Smad signaling pathway [32].
WNT signalingWNT signaling is another core fibrotic pathway, and inhibition of Wnt signaling has antifibrotic effects on different organs. WNT pathways can be subdivided into β-catenin-dependent (also referred to as “canonical” WNT signaling) and β-catenin-independent cascades (“noncanonical” WNT signaling, including Wnt/planar cell polarity and the Wnt/calcium pathway). circFBXO7 represses Wnt/β-catenin signaling through the miR-96-5p/MTSS1 axis [33]. circMTCL1 acts as an oncogene by promoting complement C1q-binding protein-dependent ubiquitin degradation and subsequently activating Wnt/β-catenin signaling [34]. circStag1 is an osteoporosis-related circRNA that mediates the cytoplasmic translocation of human antigen R and then activates Wnt signaling pathways [35].
Notch signalingNotch signaling relies on direct cell‒cell contact and is evolutionarily conserved. Membrane-bound ligands bind to Notch receptors on the target cell and activate the Notch signaling cascade. Four paralogs of Notch receptors (Notch1-4) and five different ligands (Delta-like 1 (DLL1), DLL3 and DLL4) and Jagged protein families (JAG1 and JAG2) have been described in mammals. Aberrant Notch signaling activation has been closely associated with fibrotic diseases [36]. circ_0111277 activates HTRA1/Notch-1 signaling by sponging miR-494-3p [37], whereas circ-ASH2L upregulates Notch 1 expression by serving as a sponge for miR-34a [38].
Platelet-derived growth factor (PDGF) signalingPDGF has four isoforms (A, B, C and D) and forms five dimeric proteins (AA, BB, AB, CC and DD). The engagement of two PDGF receptor tyrosine kinases, PDGFRα and PDGFRβ, activates mitogen-activated protein kinase and PI3K, as well as the small RHO family GTPases involved in cell motility, SRC and other nonreceptor tyrosine kinases, and phospholipase Cγ. PDGF signaling promotes fibrosis by acting as a mesenchymal cell mitogen and by stimulating collagen contraction. circRNA_000543 [39] and CircCDK14 [40] increase PDGFRβ and PDGFRα by sponging miR-9 and miR-3938, respectively.
The emerging roles of circRNAs in fibrotic diseasesCommon diseases associated with fibrosis include chronic kidney disease (CKD), cirrhosis, hepatitis, nonalcoholic steatohepatitis, myocardial infarction, heart failure, diabetes, idiopathic pulmonary fibrosis, and scleroderma. Aberrant expression of circRNAs has been implicated in a wide variety of diseases, especially cancers, neurological disease, and metabolic disorders. Mounting evidence has outlined functional roles for circRNAs in promoting fibrotic diseases or exerting protective effects against them [41]. Here, we provide a comprehensive update of circRNA functions in different organ systems (Table 1).
Table 1 The summaries of representative circRNAs in different organ fibrosisCardiac fibrosisCardiovascular diseases remain the leading cause of death worldwide, causing nearly 18 million deaths annually [42]. Cardiac fibrosis represents a unifying theme across various etiologies, including coronary artery disease, myocardial infarction, and diabetes mellitus. Myocardial fibrosis can be divided into reactive or replacement fibrosis based on the pathological manifestations or interstitial, perivascular and subendocardial fibrosis, according to the locations of ECM deposition [43]. Accumulating evidence has shown that circRNAs are involved in cardiac fibrosis [44]. Doxorubicin, an extensively used chemotherapeutic drug, induces cardiac remodeling manifesting as cardiofibrosis, hypertrophy, dilation, cardiomyopathy, and decreased contractile strength due to its pronounced cardiotoxicity. circNlgn is highly abundant in the heart, whereas its parental gene Nlgn is hardly detectable. The translated product of circNIgn–Nlgn173 mediates the side effect of doxorubicin-induced cardiac fibrosis by binding to and activating H2AX, subsequently upregulating inflammatory cytokines, including IL-1b, IL-2Rb, and IL-6, and early growth response molecules [45]. The increased nuclear localization of circNIgn is mediated by interacting with the structural protein LaminB1 through its unique 9-amino-acid motif, which ultimately leads to aberrant collagen deposition, cardiac fibroblast proliferation, and reduced cardiomyocyte viability [46]. Another highly conserved circRNA insulin receptor (circ-INSR) protects against doxorubicin-mediated cardiotoxicity and represses cardiac remodeling by cooperating with single-stranded DNA-binding protein 1, which improves mitochondrial function by stabilizing mtDNA [47]. A 208-amino acid protein derived from circ_0036176 (back-splicing from exon 2 to exon 4 of myosin IXA) inhibits the proliferation of cardiac fibroblasts by suppressing the cyclin/retinoblastoma tumor-suppressor gene pathway, whereas miR-218-5p can bind to circ_0036176 and inhibit its translation, thus abolishing the effect of circ_0036176 on inactivating cardiac fibroblasts [48]. Together, circNlgn, circ-INSR and circ_0036176 are representative circRNAs that can be translated into novel isoforms and confer unique functions beyond their parental genes, suggesting that these coding circRNAs and derived peptides are potential therapeutic targets for cardiac injury and fibrosis.
CircRNAs also modulate cardiac fibrogenesis by interacting with key proteins, affecting m6A modification, or via exosomes. circYap is downregulated in cardiac diseases, and ectopic expression of circYap significantly improves cardiac function and mitigates cardiac fibrosis. Mechanistically, circYap binds to gamma-actin and tropomyosin-4 (TPM4) and increases the inhibitory effect of TPM4 on actin polymerization, thus exerting an antifibrotic role in cardiac tissue [49]. circZNF609 regulates the expression of yes-associated protein (YAP) by balancing the recognition of the m6A reader protein YTHDF by Yap mRNA, thus affecting Hippo-YAP and Akt signaling. Downregulation of circZNF609 exhibits a cardioprotective effect and combats myocardial remodeling [50]. Extracellular circRNAs can also serve as messengers mediating crosstalk between different cell types in myocardial remodeling [51]. CircUbe3a from M2 macrophage-derived small extracellular vesicles promotes the activation, migration, and phenotypic transformation of cardiac fibroblasts via the miR-138-5p/RhoC signaling pathway, which ultimately aggravates cardiac fibrosis after acute myocardial infarction [52]. These studies highlight the diverse functions of circRNAs in cardiac remodeling and provide potential targets for antifibrosis therapy.
Recently, Yang et al. attempted to explore the effect of circRNA-based therapy in a mouse model by using circularized antisense RNA to block sense circslc8a1. circslc8a1 is the most abundant circular RNA in the mouse heart and is significantly decreased 8 to 12 weeks after transverse aortic constriction surgery. Silencing cardiac-specific circslc8a1 by its antisense induces cardiac hypertrophy, fibrosis and hepatic steatosis, and overexpressing circslc8a1 in vivo improves heart function and alleviates hepatic steatosis [53]. This study provided the foundation for potential clinical applications in circular RNA therapy.
Liver fibrosisChronic liver diseases, including viral infections (such as hepatitis B and C), autoimmune hepatitis, alcoholic steatohepatitis, nonalcoholic steatohepatitis, and progressive metabolic diseases, gradually result in liver dysfunction and fibrosis. Eventually, fibrosis progresses into advanced stage cirrhosis. Patients with decompensated cirrhosis develop life-threatening complications, and most die within a median time of approximately two years [54]. Despite increasing mortality and morbidity due to hepatic fibrosis, there are no approved antifibrotic therapies, and liver transplantation is currently the only treatment option [55]. Therefore, there is an exigent need to clarify the cellular and molecular mechanisms underlying hepatic fibrosis. Recent studies have revealed functional roles for circRNAs in liver fibrosis [56], most of which are focused on their role as miRNA sponges in hepatic stellate cells (HSCs). For example, circular RNA RSF1 promotes HSC proliferation and proinflammatory and profibrotic phenotype changes by targeting the miR-146a-5p/RAC1 axis [57]. Hsa_circ_0070963 [58] and circFBXW4 [59] inhibit liver fibrosis by sponging miR-223-3p and miR-18b-3p, thus upregulating LEMD3 and FBXW7, respectively. CircRNA derived from the Pleckstrin and Sect. 7 domain-containing 3 (PSD3) gene (circPSD3) is significantly downregulated in HSCs and liver tissues of mice in the CCl4-induced mouse model of liver fibrosis. Loss-of-function and gain-of-function analyses suggest that circPSD3 inhibits the activation and proliferation of HSCs by acting as a sponge for miR-92b-3p and subsequently promotes the expression of Smad7 [60]. Another circRNA derived from the MTO1 gene, namely, cMTO1, sponges miR-181-5p and enhances the expression of PTEN in HSCs, thereby exerting an antifibrotic role [61]. Additionally, hsa_circ_0004018 [62], circ608 [63], circCREBBP [64], and circUbe2k [65] suppress or promote hepatic fibrosis in the same manner as miRNA sponges.
Exosomal circRNAs could serve as messengers, disease biomarkers or therapeutic targets in liver fibrosis. circCDK13 packaged in human bone marrow mesenchymal stem cell-derived exosomes inhibits the activation and proliferation of HSCs and attenuates liver fibrosis in vivo. Mechanistically, exo-circCKD13 promotes the transcription of MFGE8 by targeting miR-17-5p/lysine acetyltransferase 2B and inhibits the PI3K/AKT and NF-κB signaling pathways [66]. mmu_circ_0000623 expression is downregulated in CCl4-induced fibrotic mouse livers, and mmu_circ_0000623-modified adipose-derived mesenchymal stem cell exosomes suppress CCl4-induced liver fibrosis by promoting autophagy through the miR-125/ATG4D axis [67]. Transfer of mesenchymal stem cell-derived exosomal circDIDO1 suppresses HSC activation through the miR-141-3p/PTEN/AKT pathway, providing a potential therapeutic target [68].
Groundbreaking research by Zhao et al. showed that although circRNAs transcribed by the mitochondrial genome account for less than 0.1% of overall circRNomics, they contribute nearly 40% of the downregulated circRNAs in fibroblasts from livers with nonalcoholic steatohepatitis [69]. Mitochondrial-localized steatohepatitis-associated circRNA ATP5B Regulator (SCAR), which is regulated by PGC-1α, binds to ATP5B. SCAR suppresses mitochondrial permeability transition pore opening and inhibits the release of reactive oxygen species and collagen contraction in HSCs. Interfering with this mito-circRNA mitigates high-fat diet-induced cirrhosis and insulin resistance in mice, thus serving as a therapeutic target.
Pulmonary fibrosisPulmonary fibrosis is an age-related, progressive and fatal disease with a median survival time of 3–5 years after diagnosis [70]. Chronic obstructive pulmonary disease, idiopathic pulmonary fibrosis, asthma, cystic fibrosis, and silicosis are common etiologies of pulmonary fibrosis. Various types of circRNAs have been discovered to be differentially expressed and involved in the pathogenesis of lung fibrosis. circANKRD42 promotes the nuclear translocation and translation of YAP1 through the miR-324-5p/AJUBA and miR-136-5p axes, mediating the crosstalk between mechanical stiffness and biochemical signals in lung fibrosis [71]. CircRNA TADA2A retards pulmonary fibrosis by targeting miR-526b and miR-203 and upregulates Caveolin (Cav)-1 and Cav-2, thereby inhibi
留言 (0)