
記住我

The human brain is the most complex organ in our body, with many aspects of its development and human-specific pathology remaining unknown because of limited accessibility to living human brain tissue. As a result, many approaches and models have been generated in an attempt to recapitulate the human brain in an in vitro system.
Two-dimensional (2D) cell culture systems have historically been used to model various diseases and systems in vitro; however, they are quite limited when it comes to modeling complex systems such as the human brain. 2D-cell culture systems lack the cellular organization that is present in the brain, as they are grown in a monolayer format which limits cell interactions only to their periphery. This results in a lack of proper oxygen and nutrient diffusion as well as waste clearance (Antoni et al. 2015). Furthermore, 2D systems lack the tissue complexity that is present in the brain. As cells are often cultured as a single cell type, they are missing the cell-to-cell interaction between different cell types that is present in vivo. To create a 2D system that is more resemblant to cellular interactions in the human brain, co-cultures of cells can be created, such as co-culturing neurons with other neural cell types such as microglia (Haenseler et al. 2017; Vahsen et al. 2022). Despite these co-culturing techniques, the cells are still limited to peripheral contacts and lack the organization needed to recapitulate the human brain in vitro.
Various neuronal cells, such as primary neurons and neuroblastoma cells, have been used in 2D systems in neuroscience research (Liu et al. 2022). However, obtaining patient-derived brain tissue, or neural stem cells and embryonic stem cells that can differentiate into neuronal cells, is controversial and not always accessible (Gabriel and Gopalakrishnan 2017). The innovation of induced pluripotent stem cell (iPSC) technology eased this problem by opening the doors to being able to generate human neural cell types without the need to isolate cells directly from the human Central Nervous System (CNS) (Gabriel and Gopalakrishnan 2017). The somatic cell-derived iPSCs are a type of stem cell that can differentiate into other cell types in the body (Lyadova and Vasiliev 2022), which makes them an available source for researchers (Karagiannis and Kim 2021). iPSCs were developed by Takahashi and Yamanaka 2006 at Kyoto University in Japan, where they first induced pluripotency in mouse embryonic stem cells (ESCs) and then in adult human fibroblast cells a year later (Takahashi and Yamanaka 2006; Takahashi et al. 2007). Takahashi and Yamanaka generated their iPSCs from fibroblast culture by using retroviral vectors to introduce specific transcription factors such as Oct3/4, Sox2, Klf4, and c-Myc into the skin cells. They noted that their iPSCs display both the morphology and growth properties of ESCs and express ESC-specific marker genes (Takahashi and Yamanaka 2006; Takahashi et al. 2007). Researchers have created techniques for generating iPSCs without using viral vectors, like plasmid-based or episomal reprogramming, that can eliminate the dangers linked to mutations due to viral integration into the genome (Bang et al. 2018). This method involves electroporation, where the electrical shocks introduce plasmid into the genome of the cells. Additionally, CRISPR-based reprogramming allows for the precise and efficient editing of certain genes for successful reprogramming (Liu et al. 2018).
Human iPSCs (hiPSC) prove a powerful tool for easily generating human neural cell types in culture; however, the differentiated cells are still 2D and limited in the same way other 2D cell culture systems are, as listed previously. This led to the need to develop alternative models to study such systems in vitro while mimicking an in vivo environment, such as 3D culture systems. One of the first successful 3D neural systems was that of neurospheres. Neurospheres are 3D cell aggregates of multipotent neural stem cells (NSC) grown in culture, providing a good resource for studying NSCs in vitro (Soares et al. 2021). These clusters of NSCs can then be differentiated into varying cell types, such as neurons and glial cells, all within the same sphere, also known as a neural spheroid (Dingle et al. 2015; Zhou et al. 2016; Pamies et al. 2017). This further allows for a better representation of cell-to-cell interactions in vitro. Although these systems provide a better representation of the brain in vitro than traditional 2D cell culture, they lack complete cellular composition, organization, and complexity of the human brain (Reynolds et al. 1992; Pamies et al. 2017). The human brain has very specific region specificity and cellular organization that is crucial to its function, and this is lacking in neurospheres and neural spheroids as the cells do not organize (Dingle et al. 2015).
Unlike 2D cell culture systems and neurospheres, brain organoids are able to model the human brain at a cellular, structural, and developmental level, allowing researchers to model the human brain and its function in ways that were previously impossible. Brain organoids were first generated by Lancaster et al. in (2013) as a system to study microcephaly. They were able to successfully generate an iPSC-derived 3D cell system, which they dubbed “cerebral organoids” (COs), that displayed discrete brain regions, dorsal cortical organization, functional cortical neurons, and glial cell populations (Lancaster et al. 2013). The development of this system has been a major breakthrough in neural sciences research as it was the first time the human brain was able to be recapitulated in vitro with correct organization and patterning.
To generate organoids, specific conditions, like extracellular matrix (ECM), small molecules, and growth factors, are provided to iPSCs or tissue-derived cells (TDCs) (Zhao et al. 2022). Thus, this environment will differentiate iPSCs or TDCs into the tissue of interest, such as the lung, heart, and cerebral cortex (Zhao et al. 2022). Researchers use stem cells such as iPSCs to generate brain organoids due to their availability (Gabriel and Gopalakrishnan 2017). This method involves differentiating single-cell iPSCs into embryoid bodies (EBs) and then NSCs by using small molecules and growth factors (Hong et al. 2022). Neuroepithelium cells form during the induction phase of EBs (Hong et al. 2022). The expansion phase involves embedding the EBs in ECM such as Matrigel, which results in a budding morphology and promotes further differentiation into several cell types present in COs, such as NSCs, neurons, and glial cells (Agboola et al. 2021). The expanded EBs are cultured in suspension on an orbital shaker (Lancaster and Knoblich 2014) or in a spinning bioreactor (Qian et al. 2016) during and after the maturation phase, where they become self-organized COs.
The organoids generated using these protocols, known as “unguided organoids,” as they are allowed to freely organize themselves into forebrain, midbrain, and hindbrain regions (Lancaster et al. 2013; Qian et al. 2016). This allows for a recapitulation of the entire brain in vitro which is an extremely useful tool; however, some diseases affect specific regions of the brain. As a result, it is necessary to be able to model specifically the forebrain, midbrain, or hindbrain alone, as well as specific structures in the brain as organoids.
Many groups of researchers have worked to develop guided protocols using extrinsic factors to generate brain region-specific organoids, which contain more accurate cell populations and organization of specific brain regions and structures. A commonly used guided organoid is a cortical organoid, which is representative of the cerebral cortex. Cortical organoids have been used to study a variety of neural disorders, such as Zika virus (ZIKV) infection (Qian et al. 2016), Japanese encephalitis virus (Zhang et al. 2018), Alzheimer’s disease (AD) (Raja et al. 2016), and several other neural degenerative disorders. Beyond cortical organoids, many other brain regions and structures have successfully been generated using guide protocols. These include forebrain and midbrain organoids (Raja et al. 2016; Jo et al. 2016), thalamic and hypothalamic organoids (Xiang et al. 2020; Huang et al. 2021), pituitary organoids (Matsumoto et al. 2020), cerebellar organoids (Ballabio et al. 2020) and hindbrain/brainstem organoids (Eura et al. 2020) (Fig. 1).
Fig. 1
Differentiation of different types of human brain organoids from stem cell–derived embryoid bodies using either guided or unguided maturation protocols, as well as assembloid formation (created with biorender.com). Cerebral organoids were stained with H&E and neurons, microglia, astrocytes, and oligodendrocytes were visualized by immunohistochemistry (IHC) TUJ-1/MAP2, IBA1, GFAP, and Olig2 specific antibodies, respectively
These region-specific organoids can be studied alone, but they can also be combined into what is known as an “assembloid” and form an even more complex brain-like structure (Fig. 1). Bagley et al. fused dorsal and ventral forebrain organoids to generate the dorsal–ventral axis and were able to show that these fused organoids can model the complex interactions between different regions of the brain (Bagley et al. 2017). In another study, Xiang et al. fused thalamic organoids with cortical organoids to investigate the circuit organizations and related disorders between the thalamus and cortex (Xiang et al. 2019). Furthermore, Miura et al. generated cortico-striatal assembloids to model complex long-distance forebrain circuits (Miura et al. 2022).
Although brain organoids prove to be a significantly better method for modeling the brain in vitro, they are not without limitations. Most notably, organoids have limited diffusion of nutrients and oxygen to their centers, and as a result, are often limited in size and are prone to cell death within their core (Lancaster et al. 2013). The limited diffusion is likely a result of the lack of a circulatory system within the organoid. In an attempt to overcome this challenge, researchers have attempted to create vascularized organoids though several different methods. One method used successfully by Pham et al. was to embed cerebral organoids with endothelial cells derived from the same line of iPSCs (Pham et al. 2018). Another approach used was to create assembloids between cerebral organoids and brain vascular organoids, which was able to recapitulate several aspects of the blood–brain-barrier (Sun et al. 2022). Additionally, the vascularization of organoids has been done using an in vivo model of transplantation. Mansour et al. transplanted human iPSC-derived brain organoids into adult mice brains and were able to see extensive vascularization, as well as neuronal integration between the organoid and the host brain, and microglial infiltration (Mansour et al. 2018). Vascularization of organoids is still a work in progress; however, there are many promising approaches to improving this system and improving organoids as in in vitro models of the human brain.
Cerebral organoids are often generated by pushing iPSCs into a neuroectoderm lineage by inhibiting the formation of the mesoderm and endoderm. As a result, many organoids used have reportedly lacked microglia, which are derived from the mesoderm and are critical to studying immune responses in the brain. A study done by Dos Reis et al. incorporated microglia into their organoids by integrating human immunodeficiency virus (HIV) infected microglia into 2-week-old organoids (dos Reis et al. 2020, 2023). However, Ormel et al. showed that microglia can innately develop within a CO (Ormel et al. 2018). This represents a valuable resource for studying interactions between microglia, neurons, and other glial cells as well as modeling impacts of immune responses on the brain, such as viral infections.
Neurotropic viral infections have been difficult to accurately study both in vitro and in vivo, as a result of the lack of a multicellular composition in vitro and differences between human and mice brains in vivo. Of importance is that viruses can either directly infect cells to cause an effect, or uninfected cells may be affected indirectly as a result of infected cells releasing cytokines, chemokines, viral proteins, and other toxic factors. This is especially important in looking at viruses such as HIV, which infects microglial cells and possess neurotoxic effects on neurons that are not prone to HIV infection (Kovalevich and Langford 2012). Furthermore, in vivo mice systems do not accurately represent the human brain, and certain viruses such as JC virus (JCV) and HIV-1 cannot infect mice, providing a need for a better human-based system. Therefore, 3D brain organoids may provide an incredibly powerful system for modeling human neurotropic viral infections in vitro.
NeuroHIV modeling in 3D brain organoidsHIV is a retrovirus that enters target cells through the interaction of viral proteins and host cell receptors CD4 and CC-chemokine receptor 5 (CCR5) or CXC-chemokine receptor 4 (CXCR4) (Deeks et al. 2015). Shortly after initial infection, HIV enters the CNS and establishes viral reservoirs leading to neuropathogenesis (Enting et al. 2001; Zayyad and Spudich 2015). In the brain, microglia are the major cell type infected by HIV, while infection of astrocytes remains controversial even though they may play a key role in neuropathogenesis (Brack-Werner 1999; Wallet et al. 2019). Neurons are typically not infected with HIV but can be injured by indirect mechanisms, such as toxic viral proteins and neurotoxicity from glial activation (Kovalevich and Langford 2012). Early in the HIV/AIDS pandemic, pre combination antiretroviral therapy (cART) era, an estimate of 20–30% of HIV patients developed HIV-associated dementia (HAD) (González-Scarano and Martín-García 2005). With the advent of cART, the incidence of HAD has significantly decreased, although a large percentage of people with HIV (PWH) still develop neurological disorders, grouped into the terms of HIV-associated neurocognitive disorders (HAND) (Antinori et al. 2007; Clifford and Ances 2013). HAND manifestation ranges from asymptomatic neurocognitive impairment (ANI) to mild neurocognitive disorder (MND) and HIV-associated dementia (HAD) (Clifford and Ances 2013).
Understanding the neuropathology of HIV is particularly important for elucidating mechanisms associated with cognitive impairment seen in PWH. One of the major obstacles in studying HIV neuropathogenesis is the lack of in vitro culture models which accurately recapitulate HAND, as multiple CNS cell types may contribute to the pathology. Additionally, animal models have been used in the past, such as nonhuman primates (NHP) or genetically modified mouse models. Although these animal models have helped researchers understand many aspects of HIV pathophysiology, there are still some limitations in the understanding of HIV-CNS interactions contributing to neurocognitive disorders (Mallard and Williams 2018). In humans, studies on HIV neuropathology have been limited to the collection and analysis of post-mortem brain tissues. The majority of the studies on neuroHIV in vitro have been performed in 2D culture models, using immortalized microglial cell lines, peripheral blood monocyte-derived microglia (MMG), or primary human microglia isolated from human tissues (Garcia-Mesa et al. 2017; Rawat and Spector 2017; Rai et al. 2020). The recent advances in iPSC culture research and the generation of brain organoids have allowed the creation of 2D and 3D in vitro models for studying neuroHIV in humans.
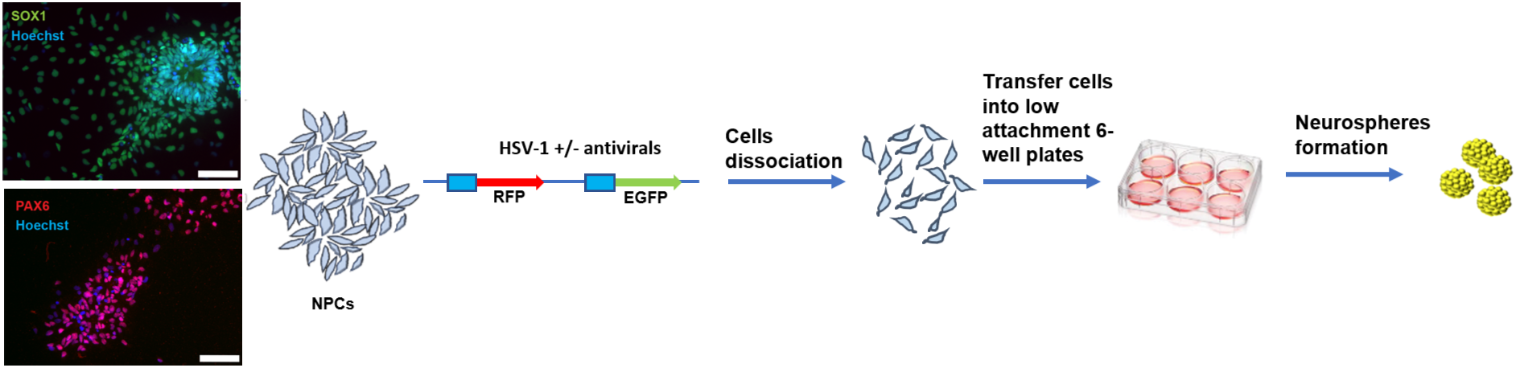
HIV modeling in human brain organoids has recently been described by two groups (dos Reis et al. 2020; Gumbs et al. 2022) with a potential to be adopted and further developed by the neuroHIV research community. These two groups used two different approaches to develop human brain organoids containing microglia, the major cell type along with macrophages responsible for HIV infection and replication in the brain (Wiley et al. 1999). Dos Reis and colleagues developed a model of 3D human brain organoids (hBORG) containing microglia (MG) which were infected with HIV prior to their integration into hBORGs. Neurospheres from neuronal progenitor cells (NPCs) were first generated using a media containing a combination of astrocyte differentiation media and neuronal media. The hBORGs generated contained only neurons and astrocytes. HMC3 cells, an immortalized human microglia cell line, were first infected with a neurotropic HIV-1 (NL(YU2-Env)-EGFP) reporter virus and then placed on top of the hBORGs. The authors showed that the majority of microglia were on the outer layer of the hBORGs, with some migrated and embedded into the organoids. In addition, HIV-infected or control (uninfected) primary human microglia from post-mortem adult human brain tissues were combined with hBORGs. Interestingly, HIV-infected human microglial cells showed some level of infiltration into the hBORGs and established productive infection. Moreover, both HMC3 and primary microglia containing hBORGs infected with HIV showed higher levels of TNF-α and IL-1β in culture supernatant compared to mock-infected MG-hBORGs. This suggested recapitulation of HIV-1 CNS pathology trademarks from post-mortem brain tissues from HIV-infected individuals (at some degree) in the MG-hBORG model. While evaluating cell toxicity in infected primary MG-hBORGs, an increase in neuronal cell death was also observed. In addition, a decrease in βIII-Tubulin (neurons) mRNA and an increase in GFAP (astrocytes) expression was observed, suggesting neuronal loss and astrocytosis, two of the characteristics of neuropathology associated with severe HAND (Huangui Xiong 2013). Although these MG-hBORG models provided advancement in the modeling of HIV infection and recapitulated some aspects of HIV CNS disease, they also possessed some limitations. Primarily, the limited incorporation of exogenously added microglial cells into organoids does not truly represents the in vivo conditions. The endogenous and natural distribution of microglia with properly developed interactions with other cell types including neurons and astrocytes within the organoids would provide a better representation of neuroHIV pathophysiology. This was recently achieved by Gumbs and colleagues (Gumbs et al. 2022) utilizing a microglia-containing 3D brain organoid model which was established earlier by the same group (Ormel et al. 2018). COs were generated from iPSCs with cells from the three germ layers at early stages during the induction and expansion phases. In particular, the formation of mesodermal progenitors with the potential to generate microglial cells was a great advancement for the development of COs with microglial incorporation. Indeed, analysis of cell-specific markers revealed that the COs contained neurons, astrocytes, and microglial cells along with CD4 and CCR5 receptor expressions in microglial cells. To evaluate the HIV-1 infectivity of these cells, authors isolated organoid-derived microglia (oMG) and infected them with CCR5 M-tropic HIV1_bal-luciferase and HIV-GFP reporter viruses along with human primary microglia (pMG) cells. As anticipated, both oMG and pMG cells supported HIV-1 infection. Once the viral infection was confirmed in CO-derived microglial cells, HIV-1 infection in COs was characterized. Interestingly, one interesting observation was the Matrigel interference of HIV infections in COs, suggesting that modeling of HIV in COs may need further improvements for reliable and reproducible infection outcomes. In addition, COs from different iPSC lines showed variations in susceptibility to HIV infection. Expression of HIV receptors, such as CD4, CXCR4, and CCR5, were also variable between different organoids. Nonetheless, the CO model of HIV infection described by Gumbs et al. provides a unique platform (although further improvements are needed) to better understand cellular and viral infections in a 3D culture system in vitro.
Human-derived brain organoid models of Herpesvirus type 1Herpesvirus type 1 (HSV-1) belongs to the Herpesviridae family (Alphaherpesvirinae subfamily) and consists of a double-stranded DNA genome of ~ 152 kbp, which encodes for ~ 80 viral proteins (Denes et al. 2020). The worldwide prevalence of HSV-1 infection in adolescents and adults is between 45 and 90%, and the primary infection usually occurs during childhood with the establishment of latent infection in sensory neurons. The primary infection sites for HSV-1 are mostly the skin and mucous membranes of the lips and less frequently the genital mucosa. After infection, viral particles retrogradely move through the axons of ganglionic neurons to where the viral DNA genome is assembled into a heterochromatin-like structure. As neurons are non-dividing and they are not susceptible to immunological surveillance, HSV-1 infection in neurons results in a lifelong lasting infection. HSV-1 can also cause herpetic keratitis, which results in corneal scarring and impairment of sight. The most detrimental HSV-associated disease is herpes encephalitis, a rare disease that affects 2–4 out of 1,000,000 people annually with high mortality rates, that occurs when the virus reaches the CNS (Bradshaw and Venkatesan 2016). Furthermore, pregnant women are at risk of mother-to-child transmission of genital herpes, which may lead to fetal encephalitis and perinatal mortality if an emergency cesarean delivery is not performed (Kimberlin 2004; Messacar et al. 2018). Pathophysiological features of HSV-1-associated neurological disorders remain unclear, impelling researchers to establish relevant in vitro models of human CNS to understand the molecular mechanism of viral latency, reactivation, and neuropathology.
HSV-1 infection has been primarily studied in either human- or non-human- derived primary or immortalized cell cultures and cell lines. However, the obtained results in many cases possess reliability and reproducibility issues due to the variations in the genetic assets of the cell lines employed in different experiments. Moreover, testing antiviral drug candidates on cell lines has often been inconsistent. Therefore, in most pre-clinical studies, the efficacy of HSV-1 vaccines has been evaluated in mice models. These in vivo models have provided useful information on immune response against primary HSV-1 infection at the expense of the vaccine’s efficacy against recurrent viral shedding and disease events, which are both extremely rare in mice (Gebhardt and Halford 2005). The lack of an animal model that translates human immune responses to viral infection still represents a major limitation to develop valid therapeutic vaccines (Dasgupta and BenMohamed
留言 (0)