
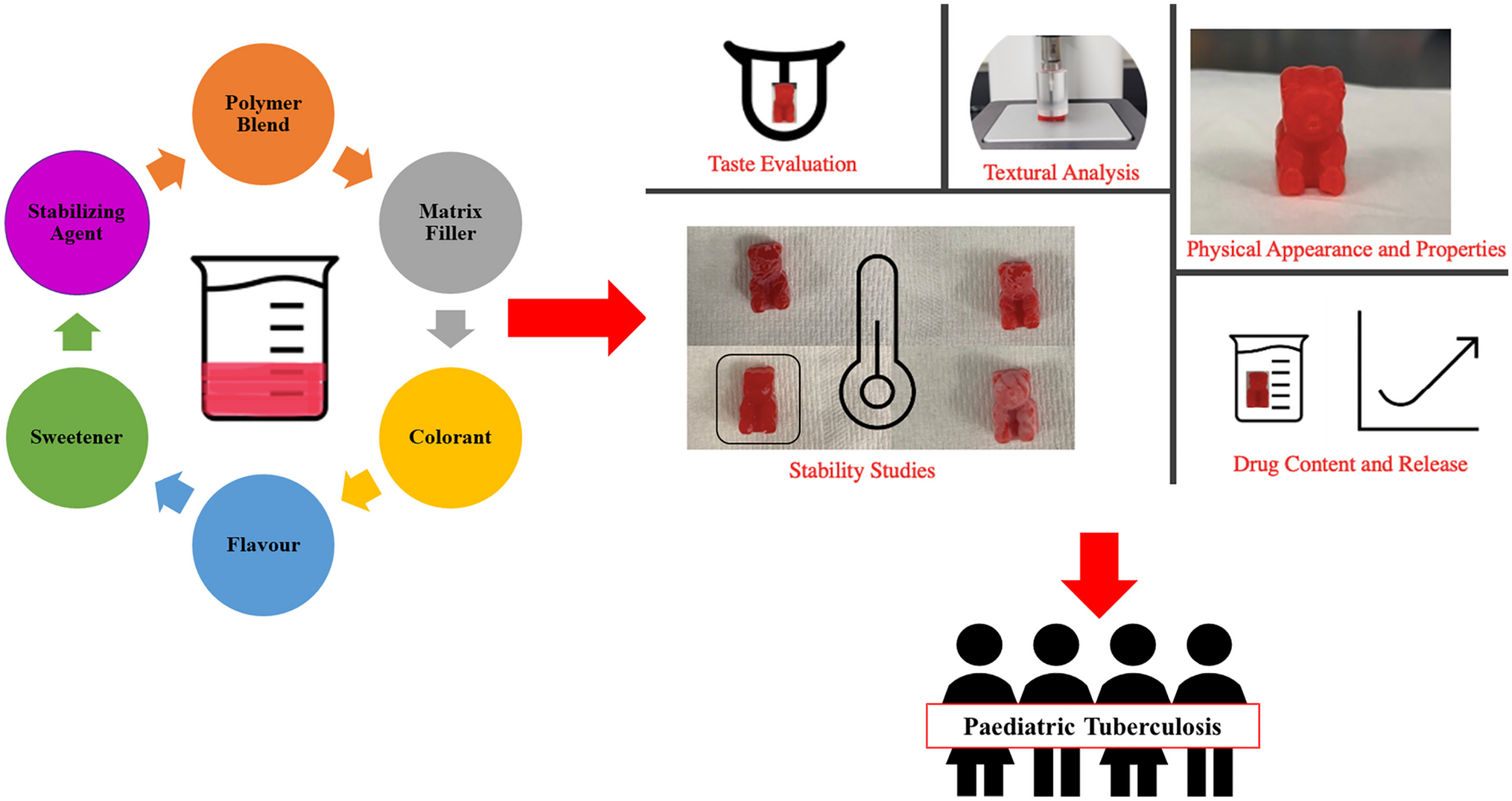
記住我
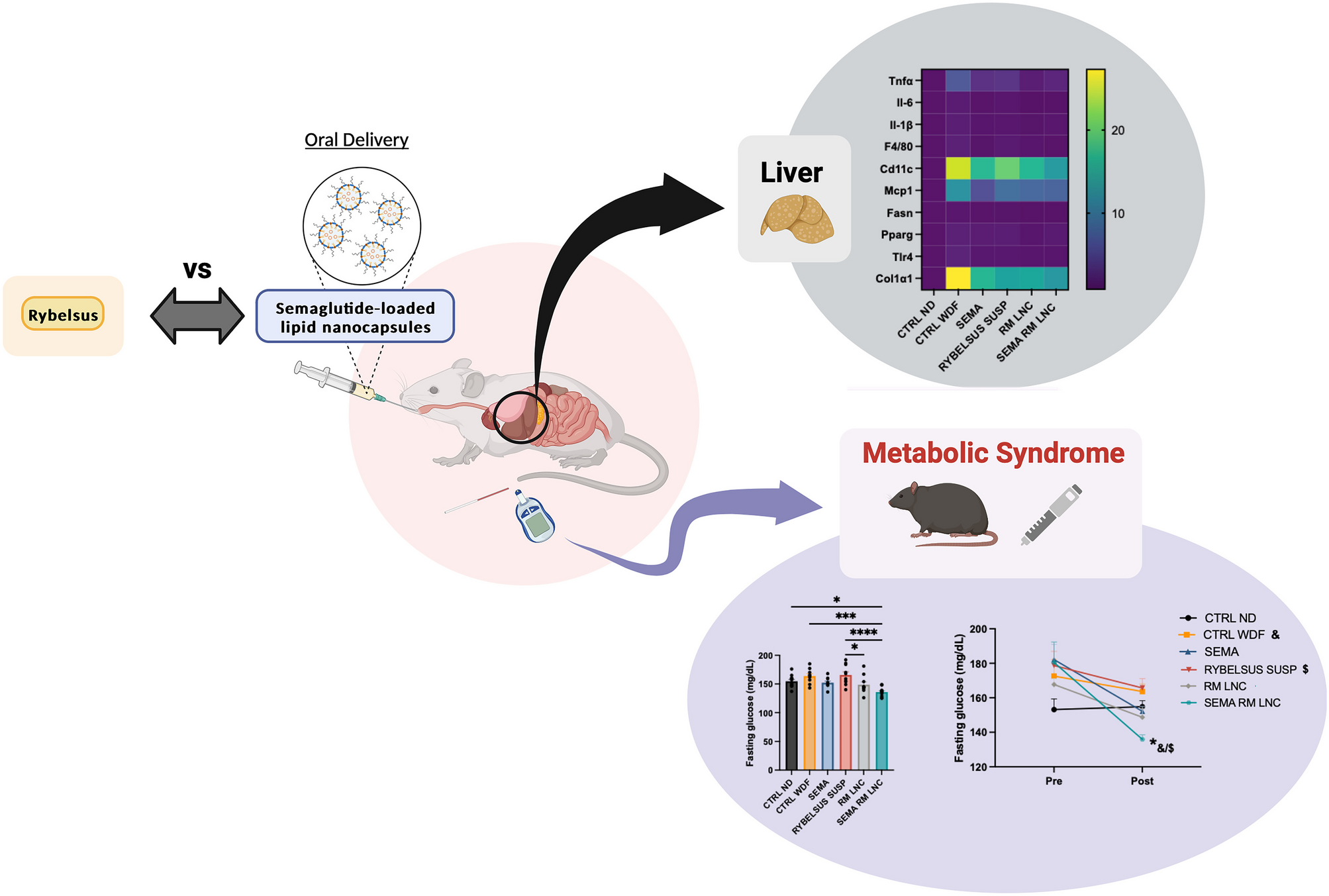
Rif-ILH microparticles of 3.11 ± 1.1 µm mean particle size were synthesized by spray drying rifampicin-encapsulated lipid nano-droplets (diameter = 160 ± 27 nm), comprised of medium chain length lipids dispersed within an inulin aqueous solution at a 1:1 lipid:inulin ratio (Fig. 1). Rif-ILH microparticles revealed a negative zeta potential of − 11.5 ± 0.53 mV, which was attributed to deprotonation of inulin hydroxyl groups when exposed to aqueous media [36]. Rifampicin-loaded lipid micro-droplets (Rif-lipid), 2.46 ± 0.8 µm in diameter with a zeta potential of − 4.18 ± 1.2 mV, were also synthesized to serve as a control system to Rif-ILH. Rifampicin loading within Rif-ILH and Rif-lipid was 5.32 ± 0.84% (w/w) and 9.12 ± 1.10% (w/w), respectively.
Fig. 1
Schematic representation of the two-step fabrication process for Rif-ILH microparticles: A an inulin solution and B a rifampicin-loaded homogenized lipid emulsion are C combined with mixing and D spray dried to form Rif-ILH microparticles
The morphology of powdered inulin particles and spray-dried Rif-ILH microparticles was investigated using SEM. As shown in Fig. 2, raw inulin powder comprised 1–2 µm particles with a spherulite-like discoid shape, which consists of stacks of lamellar sheets, according to previous investigations [6]. In contrast, Rif-ILH microparticles were mainly between 2 and 5 µm in diameter, of spherical shape and a smooth surface morphology. It is anticipated that inulin accumulated at the surface of Rif-ILH particles, with lipid nano-droplets being encapsulated within a three-dimensional inulin matrix, comparable to alternate polymer-lipid hybrid microparticles prepared through an equivalent fabrication approach [37].
Fig. 2
SEM images of dry powder aggregates of raw inulin (A and B) and rifampicin-loaded inulin-lipid hybrid (Rif-ILH) microparticles (1:1 lipid:inulin ratio) (C and D)
In vitro rifampicin release studiesILH microparticles were strategically designed with pH-responsive properties, through the inclusion of acid-labile inulin within the shell of the hybrid particles, so that triggered release of the encapsulated cargo is prompted through particle internalization within the acidic environment of macrophage lysosomes. To assess the degradation behaviour of ILH microparticles in biologically relevant media simulating the neutral extracellular environment and acidic lysosomal environment, ILH particles were dispersed in PBS (pH 7.4) and artificial lysosomal fluid (ALF, pH 4.5) [10], respectively. The degree of polymerization (DPn) of the inulin chain and particle size was monitored in a time-dependent manner, which highlighted key differences in particle behaviour between the two media environments (Fig. 3). That is, the inulin fructooligosaccharide chain was stable under neutral conditions, with only a minor reduction in chain length from DPn = 35.0 to DPn = 34.3 ± 0.5 being observed over the 4-h dispersion period. In contrast, the inulin chain was hydrolyzed in acidic ALF media, as evidenced by a reduction in DPn from 35.0 to 28.0 ± 0.8 after 4 h. pH-mediated hydrolysis corresponded with a greater reduction in particle size for ILH when dispersed in ALF, compared to PBS, where the mean particle size decreased to 0.56 ± 0.2 µm after 4 h. The mean particle size of ILH also reduced in PBS, but only to 2.51 ± 0.9 µm, which suggests that the reduction observed was due to aggregates dispersing into individual particles throughout the dispersion period, rather than particle degradation. In contrast, the reduction in ILH particle size observed in ALF, along with the corresponding reduction in DPn, indicates that ILH microparticles are prone to pH-mediated degradation.
Fig. 3
The impact of media composition on inulin degradation within ILH microparticles, evidenced through time-dependent changes in A average degree of polymerization (DPn) of the inulin chain, and B particle size of ILH microparticles. Data is represented as mean ± SD (n = 3)
To validate the pH-responsive release behaviour of rifampicin when encapsulated within ILH microparticles, drug release studies were performed in PBS and ALF, and where release behaviour from Rif-ILH was contrasted against a Rif-Lipid microemulsion of a comparable particle size. In neutral media, only 22.5 ± 3.6% of rifampicin was released from Rif-ILH particles after 1 h, in contrast to 59.0 ± 7.1% for Rif-Lipid (Fig. 4A). Moreover, rifampicin release continued to increase for Rif-Lipid over the 4-h release period, leading to 73.4 ± 3.5% release. Restricted rifampicin release was observed for Rif-ILH microparticles throughout the entire 4-h period, whereby only 29.4 ± 2.2% of rifampicin partitioned towards the aqueous media (Fig. 4), emphasizing the capacity for the inulin three-dimensional matrix of Rif-ILH to protect against drug diffusion into the aqueous media.
Fig. 4
Rifampicin release kinetics from Rif-Lipid (blue) and Rif-ILH microparticles (pink) in A neutral media (PBS, pH 7.4, 37 °C) and B artificial lysosomal fluid (ALF; pH 4.5, 37 °C). Data is represented as mean ± SD (n = 3). C Schematic representation illustrating the release behaviour of rifampicin from both Rif-Lipid and Rif-ILH microparticles in C neutral media (pH 7.4) and D acidic media (pH 4.5)
The pH-dependent release mechanism of Rif-ILH microparticles was exhibited in ALF at pH 4.5, where over three-quarters of the rifampicin was released within 1 h, with a total release of 79.1 ± 0.2 µg/mL being observed after 4 h. For Rif-Lipid, the change in pH did not induce any significant differences in rifampicin release kinetics (Fig. 4). These findings confirm that the inulin ‘coating’ protects the majority of lipid-encapsulated drug from premature release within plasma-simulating media, due to inulin forming a stable shell surrounding the encapsulated lipid nanoparticles and drug molecules. However, once exposed to acidic ALF media, inulin chains rapidly decompose into oligosaccharide and monosaccharide units, as evidenced through a decrease in DPn and particle size, thus promoting the diffusion of rifampicin from the exposed lipid nano-droplets [11,12,13,14,15,16]. This is in accordance with previous findings that have shown the pH-provoked release mechanism of drugs conjugated to inulin chains, whereby in neutral media, the drug-inulin conjugate is stable, but once exposed to an environment simulating lysosomal media, the drug is cleaved from the inulin chain due to inulin hydrolysis. One key example is where inulin was used as a carrier for the delivery of the anti-tuberculosis (TB) drug, isoniazid, to monocytes, with the inulin-conjugated isoniazid exhibiting a pH-dependent release with less than 10% release in pH 7.4 and > 40% drug release in pH 4.5 [17].
Cytotoxicity assessment of ILH microparticlesThe cytotoxicity of ILH microparticles and lipid micro-droplets was evaluated using the MTT assay in RAW 264.7 macrophages (Fig. 5). Triton-X was used as a positive control with no cellular viability being observed after 1-h incubation. Both ILH microparticles and lipid micro-droplets exhibited a dose-dependent effect on macrophage viability, whereby dose increases from 10 to 100 µg/mL led to a reduction in cellular viability, from 95.2 to 79.7% for ILH and from 96.3 to 52.9% for lipid micro-droplets. Importantly, the presence of the inulin coating for ILH microparticles reduced the cytotoxicity of the particles with the cellular viability remaining ≥ 80% even at 100 µg/mL particle concentration [9]. Subsequently, these findings validate the safety of ILH microparticles towards macrophages. Cellular uptake studies were performed at a particle concentration of 50 µg/mL for both ILH microparticles and lipid micro-droplets, since at this concentration, both systems exerted a cellular viability of > 80%.
Fig. 5
Cellular viability of inulin-lipid hybrid (ILH) microparticles (pink bars) and lipid micro-droplets (blue bars) in RAW 264.7 cells determined via MTT assay (mean ± SD, n = 3)
Cellular uptake studiesTo understand whether the inulin coating of ILH microparticles promotes cellular uptake compared to uncoated lipid micro-droplets, both formulations were fluorescently labelled with Nile red (lipophilic dye) and the intracellular localization of particles was determined based on differences in fluorescence intensities within the cells, using flow cytometry. Both the cells incubated with Nile red solution as well as control group (no treatment) exhibited no significant difference in fluorescence intensity, thus confirming the inability for the pure dye to promote uptake. Subsequently, any enhancement in Nile red intensity within the cells treated with Nile red particles can be attributed to particle uptake.
Incubation with both lipid micro-droplets and ILH microparticles triggered cellular uptake of Nile red, with 16.4 ± 4.0% and 93.4 ± 3.4% particles being internalized after 1 h, respectively (Fig. 6). Thus, the degree of particle uptake for ILH microparticles was 6- to 16-fold greater compared to the lipid micro-droplet and the pure Nile red solution after 1-h incubation. This was further evidenced using confocal microscopy, whereby there is enhanced dye internalization for ILH particles after 1-h incubation, compared to lipid micro-droplets and pure dye (Fig. 7). No additional uptake was observed for ILH microparticles between 1 and 4 h. In contrast, lipid micro-droplets continued being internalized within macrophages during this period, achieving a total of 88.3 ± 2.5% uptake after 4 h.
Fig. 6
Cellular uptake of lipid micro-droplets (blue bars) and ILH microparticles (pink bars) loaded with Nile red in RAW 264.7 cells after A 1-h and B 4-h incubation. No significant difference was observed between cells treated with Nile red (yellow bars) compared to the control group (no treatment; green bars)
Fig. 7
Confocal microscopy images for the uptake of Nile red formulations by RAW 264.7 macrophages, after 1-h incubation with A Nile red solution, B Nile red–loaded lipid microparticles and C Nile red–loaded ILH microparticles. Magnified regions of D Nile red–loaded lipid micro-droplets and E Nile red–loaded ILH microparticles. All formulations were dosed at an equivalent dye concentration of 50 µg/mL. Nuclei were stained with DAPI dye (blue region); the cellular cytoskeleton was stained with Alexa-488 (green region); and the particles were stained with Nile red (red region)
Similar outcomes were reported in earlier studies where inulin-based drug delivery systems were used for targeted intracellular antibiotic delivery to cells infected with intracellular pathogens, e.g. Mycobacterium tuberculosis [29, 38]. The results suggested the ability of inulin to promote a rapid uptake of particles by infected cells through endocytosis pathways; however. the specific receptor(s) mediating this process are unknown [23, 39, 40]. Ultimately, it is suggested that incorporation of inulin facilitated the rapid endocytosis of ILH microparticles by phagocytic cells, i.e. RAW 264.7 cells, leading to higher uptake of ILH microparticles compared to lipid micro-droplets in a shorter period of time. For both lipid micro-droplets and ILH microparticles, the particle size could be considered ideal for cellular uptake, since previous studies have highlighted the enhanced capacity for particles with intermediate particles size (~ 2–3 μm in diameter) to be engulfed by macrophages, compared to submicron particles and particles exceeding 10 µm [41]. Subsequently, it was considered that the particle size of ILH microparticles synthesized in this study was ideal for promoting cellular uptake; however, further optimization may lead to production of a more uniform particle size distribution and therefore greater enhancements in particle internalization within macrophages in key target tissues associated with intracellular infection, such as the lungs.
Rifampicin delivery to macrophagesThe degree of rifampicin internalization within macrophages was examined by incubating RAW 264.7 cells with a rifampicin solution, rifampicin-loaded ILH (Rif-ILH) microparticles and lipid micro-droplets (Rif-Lipid) (Fig. 8). After 1-h incubation, 37.3 ± 4.3% and 69.5 ± 8.1% of the rifampicin dose was detected inside the macrophages when treated with Rif-Lipid and Rif-ILH microparticles, respectively, compared to 16.5 ± 3.3% for the cells treated with pure rifampicin solution. As the incubation time increased from 1 to 4 h, the degree of rifampicin uptake when encapsulated within Rif-Lipid and Rif-ILH microparticles did not increase, indicating that rifampicin was internalized within the first hour of incubation. Furthermore, there was no significant difference in intracellular rifampicin concentration for the cells treated with Rif-Lipid compared to the pure drug solution after 4-h incubation, which did not correlate with the cellular uptake data obtained for the Nile red lipid micro-droplets. This can be attributed to the inability for lipid micro-droplets to protect the encapsulated cargo (rapid partitioning into solution), specifically rifampicin, in neutral conditions equivalent to the extracellular environment. That is, drug release data indicated that over 75% of rifampicin is released from the lipid micro-droplet after 1 h at pH 7.4 (Fig. 4). Thus, after this period, lipid micro-droplets cannot facilitate uptake of rifampicin. In contrast, Rif-ILH microparticles protect the encapsulated rifampicin from premature release in the extracellular environment, until phagocytosed by the macrophages, where the acidic lysosomal media hydrolyses inulin (Fig. 3), triggering the release of rifampicin. This highlights the important role of the pH-triggered release mechanism of Rif-ILH microparticles in transporting rifampicin to macrophages.
Fig. 8
Internal rifampicin concentration within macrophages when incubated with a rifampicin solution (orange bars), Rif-Lipid (blue bars) and Rif-ILH microparticles (pink bars), when dosed at a rifampicin concentration 50 µg/mL after A 1-h and B 4-h incubation time (mean ± SD, n = 3)
Antibacterial efficacy of rifampicin formulations against SCV S. aureusRAW 264.7 cells were infected with SCV S. aureus as a model intracellular pathogen. This pathogen is responsible for unresolved clinical problems associated with chronic infections and owing to their slow metabolism, SCV S. aureus are phenotypically smaller compared to the parent strain of S. aureus [35]. Here, the intracellular localization of SCV S. aureus within RAW 264.7 cells was visually confirmed using confocal microscope. As illustrated in Fig. 9, the cytoskeleton of both non-infected and infected macrophages was stained green with deep purple nuclei, while the aggregation of dye (green spots) was associated with bacterial cell wall stained with Alexa 488 and highlighted the presence of SCV S. aureus within the cells.
Fig. 9
The intracellular presence of SCV S. aureus as observed using confocal microscopy. The nuclei and the cell membrane of macrophages were shown as purple and green, respectively. The localized aggregation of dye associated with the bacterial cell wall within the cells (as indicated by white arrows) demonstrates the presence of SCV S. aureus within cells
The selection of rifampicin concentrations to evaluate the efficacy of formulations was based on the Minimum Inhibitory Concentration (MIC) against SCV S. aureus [34], which was previously determined to be 0.125 µg/mL [16]. Subsequently, a rifampicin concentration equivalent to 4 × MIC (0.5 µg/mL) was initially selected to ascertain the efficacy of each rifampicin formulation in triggering a reduction in intracellular bacteria. Following the 4-h treatment with rifampicin solution and Rif-Lipid, SCV S. aureus–infected macrophages exhibited a negligible reduction in CFU of SCV S. aureus, which was shown to be statistically comparable to the control group (i.e. no treatment) (Fig. 10). The poor ability of unformulated rifampicin to significantly reduce the abundance of viable bacteria was anticipated since uptake studies revealed the inability for rifampicin to be internalized within macrophages. For Rif-Lipid formulation, premature drug release in neutral media reduced the exposure of intracellular pathogens to the antibiotic drug, and thus, Rif-Lipid were unable to significantly improve the overall efficacy of rifampicin at 0.5 µg/mL. In contrast, treatment of infected macrophages with Rif-ILH resulted in a significant improvement in the antibacterial activity of rifampicin against SCV S. aureus. Rif-ILH demonstrated a ~ 4-log greater reduction in CFU compared to the rifampicin solution and Rif-Lipid. This correlates well with the macrophage uptake findings, where the intracellular concentration of rifampicin when hosted within Rif-ILH was over 2-log greater, compared to that within pure rifampicin and Rif-lipid. This further confirmed the increased intracellular concentration of the drug within the vicinity of the target site (i.e. site of pathogen confinement) and therefore facilitated more efficient bacterial killing.
Fig. 10
Efficacy of rifampicin-loaded formulations on the quantitative reduction of intracellular SCV S. aureus
To evaluate a dose-dependent effect of rifampicin-loaded formulations on antibacterial activity, infected macrophages were treated with a rifampicin dose 20-fold greater than its MIC (i.e. 2.5 μg/mL) (Fig. 10). Increasing the rifampicin dose from 4 × MIC to 20 × MIC did not provide any further improvement in bacterial reduction for Rif-Lipid (i.e. the CFU reduced from 6.5 × 105 CFU/mL to 2.7 × 105 CFU/mL), while Rif-ILH microparticles showed the greatest antibacterial activity, with a ~ 2-log reduction in CFU (i.e. 2 × 105 CFU/mL to ~ 2.2 × 103 CFU/mL). This is in agreement with the previous study where the maximum efficacy was achieved when SCV S. aureus–infected macrophages were treatment with rifampicin-loaded formulations equivalent to 2.50 μg/mL rifampicin dose [34].
Subsequently, this study reveals a new approach for enhancing the antibacterial efficacy of rifampicin against pathogens that shield themselves in intracellular environment of the host, through encapsulation within an inulin-based drug delivery system. Moreover, for the first time, utilizing the ILH carrier as a micro-encapsulation device enabled the advantageous effects of inulin as a bioactive material and delivery vehicle to be harnessed, while eliminating the need for chemical modifications to the drug via a covalent attachment mechanism. To facilitate clinical translation of this new drug delivery approach, further preclinical investigation and validation are necessary. Specifically, the efficacy of this treatment must be demonstrated across broader S. aureus strains with greater genetic diversity and in host cells that are not naturally phagocytic but are still capable of harbouring pathogens (e.g. epithelial cells). Furthermore, treatment efficacy must be clearly demonstrated within in vivo studies using validated intracellular models of infection. It is anticipated that the clinical application of micro-encapsulated antibiotics, using ILH, will reduce the high clinical dose required for effective bacterial killing and subsequently reduce severe side effects and toxicity associated with antibiotics’ over-consumption. Owing to the small particle size and low toxicity of ILH, it is anticipated that clinical administration through systemic (e.g. intravenous injection) or pulmonary (e.g. inhalation) routes will provide a novel approach for more effectively and safely treating systemic and lung infections. Previous studies have demonstrated the clinical translatability of biocompatible microparticles in treating such infections, including tuberculosis, and therefore, ILH is well-positioned for further preclinical investigation and optimization to promote translation to the clinic.
留言 (0)