
記住我



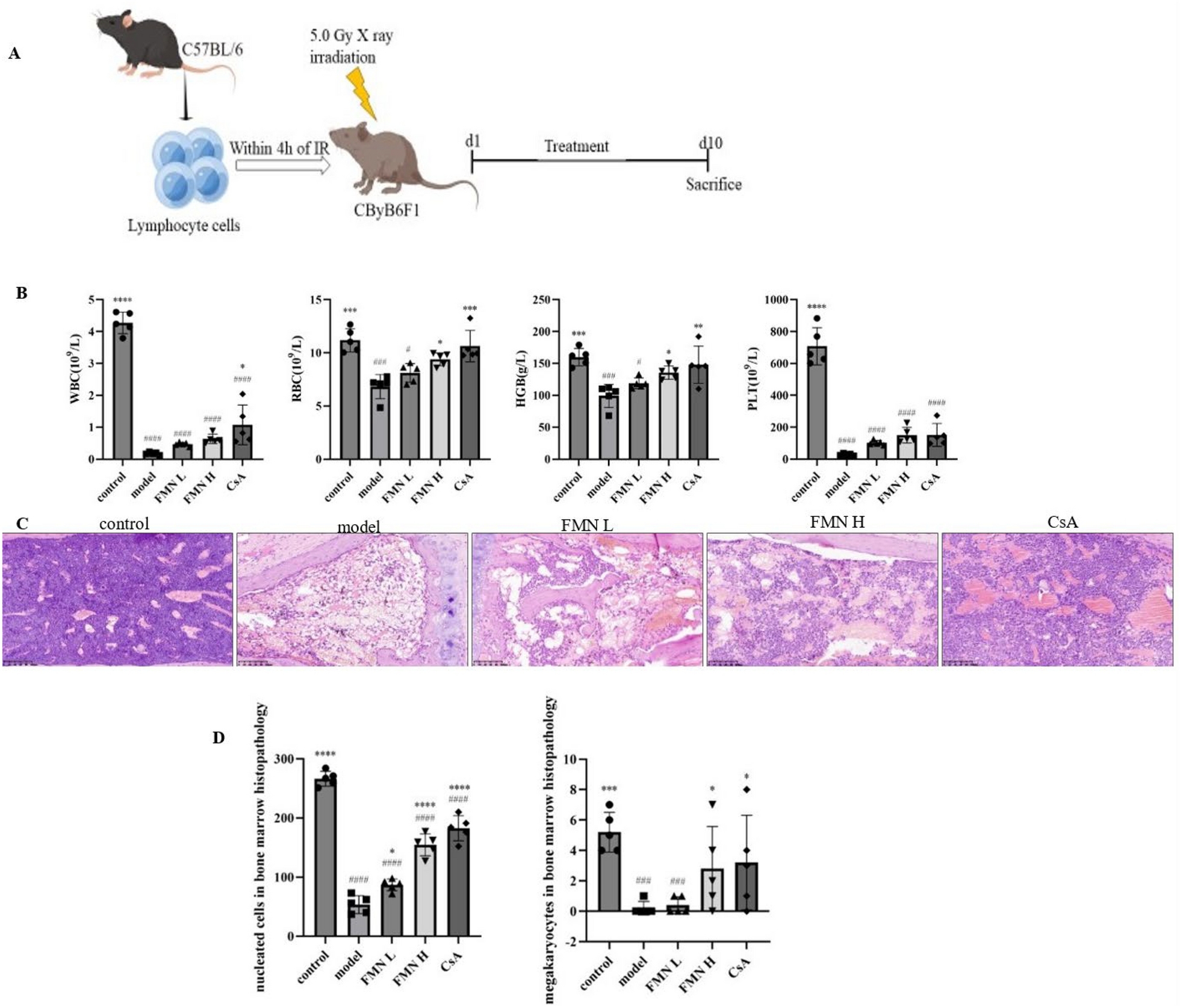
To find out the potential effective compounds of ASIV, we analyzed the microbial and hepatic biotransformation by retrieving literature [21, 22]. As shown in Fig. 2, a little of ASIV was absorbed into the portal vein. The rest was mixed with the gut microbiota. Then, the xylose or glucose moiety was removed to form 6-O-β-d-glucopyranosyl (Bra B) and 3-O-β-d-xylopyranosyl-cycloastragenol (Cyc B), respectively. Next, the microbiota sequentially degraded the products into CA, 20R, 24S-epoxy-6α, 16β, 25-tyihydroxy-9-cycloartan-3-one (CA-2H), and iso-CA. The six compounds mentioned above were first infused into the liver through the portal vein and then underwent intrahepatic conversion and biliary exertion. As a result, only ASIV, CA, and iso-CA were detected in the bloodstream.
Fig. 2
Microbial and hepatic biotransformation of ASIV. ASIV is transformed by gut microbiota to form Cyc B, Bra B, CA, CA-2H, and iso-CA. The microbiota-transformed products are infused into the liver. Then CA is partly converted into CA-2H and iso-CA. The bile duct system exerts Cyc B, Bra B, and ASIV. Only ASIV, CA, and iso-CA are detectable in the bloodstream. ASIV Astragaloside IV, CA cycloastragenol; iso-CA 3-epi-cycloastragenol, Bra B 6-O-β-d-glucopyranosyl, Cyc B 3-O-β-d-xylopyranosyl-cycloastragenol; CA-2H dehydrogenated to 20R, 24S-epoxy-6α, 16β, 25-tyihydroxy-9-cycloartan-3-one
The ADMET prediction indicated that the transformed metabolites of ASIV processed lower molecular weight (MW), easier synthetic accessibility, and higher GI absorption, bioavailability, and DL (Table 1). These results were in agreement with the previous study indicating much higher OB of CA (25.7%) than ASIV (2.2–3.7%) in rats [22, 38]. Therefore, CA and iso-CA detected in the blood may be the additional active compounds of ASIV. Moreover, while ASIV was difficult to permeate the BBB, its microbial-transformed products, CA and iso-CA, could enter the brain parenchyma (Table 1). Taken together, ASIV, CA, and iso-CA were the candidate effective compounds of ASIV. Moreover, CA and iso-CA, other than ASIV itself, seemed to be the major effectors of ASIV in targeting CNS where BBB exists. Therefore, it is necessary to add the biotransformation analysis before the network pharmacology scheme to explore the therapeutic mechanism of ASIV in treating ICH.
Table 1 The predicted ADME properties, drug-likeness, pharmacokinetic, and physicochemical properties of Astragaloside IV and its derivatesMicrobial and hepatic biotransformation increases the potential targets of ASIV in treating ICHTo evaluate the advantage of our new strategy in discovering new targets, we screened the potential targets of ASIV before and after biotransformation. SwissTargetPrediction, SEA, and PharmMapper databases were applied. These tools work differently to predict targets: SwissTargetPrediction and SEA prediction rely on the topologic similarity to the known ligands [30, 39], and PharmMapper is based on a pharmacophore mapping approach [32]. As a result, a total of 373, 339, and 335 proteins were targeted by ASIV, CA, and iso-CA, respectively (Fig. 3). Among them, as many as 215 targets were common for ASIV, CA, and iso-CA. It is probably because of their similar backbones and functional groups. The results suggest that biotransformation enhances the abilities of ASIV to affect its targets. Moreover, 150 proteins were additionally targeted by CA or (and) iso-CA but not by ASIV. It implies that microbial and hepatic biotransformation add new effectors to ASIV functions.
Fig. 3
The potential targets of ASIV before and after microbial and hepatic biotransformation. There are 373 and 524 targets of ASIV before and after biotransformation, respectively. Two hundred and fifteen targets are common for ASIV, CA, and iso-CA (purple circles). The biotransformation results in 150 additional targets. ASIV Astragaloside IV, CA cycloastragenol, iso-CA 3-epi-cycloastragenol
To address the importance of microbial and hepatic biotransformation in ASIV treatment for ICH. The ICH targets were screened out and intersected with the compound targets before and after transformation. As displayed in the Venn diagram, 64 ICH-related genes were also targeted by ASIV. After biotransformation, the ASIV-associated ICH targets increased by 27 (Fig. 4A). The pre-transformation ASIV targets-ICH PPI network contained 64 nodes and 663 edges (Additional file 1: Fig. S1A). And the active compound-targets-ICH network included 91 nodes and much more edges (1192) than ASIV alone (Additional file 1: Fig. S1B). The additional targets-ICH network comprised 27 nodes and 63 edges (Fig. 4B). The core clusters resulting from MCODE included 13 nodes. The associations of these targets with ICH and the corresponding ingredients are displayed in Table 2. Among them, the central nodes predominantly included genes associated with cell migration (PTK2, CDC42, CSF1R, HGF), proliferation (PIK3R1, HGF, CSF1R), and inflammation (STAT1, TNF). (Fig. 4C). Then, to uncover the targeted cell type of the additional genes after biotransformation, we mapped the 27 additional genes to Brain RNA-Seq, a brain transcriptome database. The transformation-added targets are primarily expressed in microglia/macrophage (Fig. 4D).
Fig. 4
The biotransformation-added targets of ASIV on ICH. A Twenty-seven additional genes overlap with ICH targets. B The PPI network of the 27 additional targets. C The distribution analysis of 27 additional targets suggests that the biotransformation products primarily act on microglia/macrophages. D The core subnetwork of 27 additional targets shows the central role of HRAS, PIK3R1, PTK2, CDC42, CSF1R, HGF, STAT1, and TNF
Table 2 The associations of the core genes to ICH and the herbal productsFurther functional enrichment analyses of the additional targets after ASIV biotransformation showed that most of the top enriched pathways were also relevant to cell migration (GO: positive regulation of cell migration; KEGG: Rap1 signaling pathway; KEGG: Focal adhesion), proliferation (GO: regulation of cell proliferation), and inflammation (GO: chemokine signaling pathway) (Fig. 5; Additional file 1: Table S1), which differed from the results of ASIV [17] (Additional file 1: Table S2).
Fig. 5
GO and KEGG analyses of the biotransformation-added targets. A The top 10 biological processes, cellular components, and molecular functions of ASIV enriched in GO analysis emphasize cell migration and proliferation. B The top 30 KEGG pathways indicate the essential roles of chemokines and focal adhesion on ASIV in treating ICH
Microglia is the resident macrophage in the central nervous system (CNS). It is the primary cell to regulate neuro-inflammation by producing cytokines like TNF-α. Besides, the activated microglia can proliferate and migrate. [33]. Though indispensable in CNS homeostasis, microglia and brain-infiltrating monocyte-derived macrophage are the instigators of secondary brain injury after ICH [34, 35]. Upon ICH, the perihematomal microglia/macrophage is activated as early as 1 h and 4 h in the collagenase- and auto-blood-induced ICH models, respectively [34]. In addition, microglia migrate to the lesions within 12 h [36]. Then, microglia/macrophage increases 8 folds in 3–5 days, exacerbating inflammation and secondary brain damage [34, 36]. Therefore, we focused on the biological processes directly related to microglia/macrophage activity. As a result, “GO: positive regulation of macrophage chemotaxis” and “GO: macrophage differentiation” were screened out, where CSF1R and PTK2, CDC42, and CSF1R hit (Additional file 1: Table S1). As predicted, CDC42, PTK2, and CSF1R were targeted by CA. In addition, CSF1R was also bound to iso-CA. In contrast, the biological processes “GO: positive regulation of macrophage chemotaxis” and “GO: macrophage differentiation” were not significantly enriched by the ASIV targets (Additional file 1: Table S2), nor were they reported by previous network pharmacology-based studies [17].
Molecular docking and molecular dynamics simulation of ASIV derivatives and the additional targets.We employed molecular docking to validate the additional effects of ASIV derivatives on ICH. As shown in Additional file 1: Fig. S2, CA formed 5 hydrogen bonds (H-bonds, blue dashed line) with CDC42, 5 H-bonds and 4 hydrophobic interactions (gray dashed line) with CSF1R, and 5 H-bonds and 6 hydrophobic interactions with PTK2, respectively. Besides, iso-CA (Blue-green sticks) was anchored into the active pocket of CSF1R via 4 H-bonds and 4 hydrophobic interactions. The lowest binding energy of CA-CDC42, CA-PTK2, CA-CSF1R, and iso-CA-CSF1R were − 7.5, − 7.3, − 7.9, and − 8.0 kcal/mol, respectively (Additional file 1: Fig. S2A–D; Table 3), indicating considerable binding affinities of the complexes.
Table 3 Results of molecular dockingMolecular docking calculates the most stable conformations of the ligand-target complexes statically. However, whether the putative interactions are dynamically stable in a virtual environment remains unknown. To address this, a molecular dynamics simulation was performed. During the simulation, the atmosphere was set at physiological conditions, including ionic strength, temperature, and pressure [40]. In this method, root mean square deviation (RMSD) reflects the dynamic variations in the conformational stability of backbone molecules to the initial state. Root mean square fluctuation (RMSF) represents the stability of specific residues in the receptors. And the radius of gyration (Rg) evaluates the structural compactness of complex biological systems [41]. The results indicated that during the 80-ns simulation. The RMSD and Rg of the CA-CDC42, CA-PTK2, and iso-CA-CSF1R complexes were relatively smooth with limited fluctuation (Fig. 6A–D). Moreover, the RMSFs were low, especially for CA-CDC42, which suggested satisfactory stability of the above complexes. In these complexes, the ligands bound to the active pockets of their corresponding receptors and formed 0–4 hydrogen bonds during the 80-ns simulations (Fig. 6E). The estimated lowest binding free energy of CA-CDC42, CA-PTK2, and iso-CA-CSF1R were − 87.675 kJ/mol, − 64.234 kJ/mol, and − 84.796 kJ/mol, respectively. Although the lowest binding free energy of CA-CSF1R was relatively low (− 67.632 kJ/mol), the overall dynamic performance of the CA-CSF1R complex was poor, as suggested by the sizeable RMSD fluctuation (Fig. 7A), unfixed binding pocket (Additional file 1: Fig. S3B), and limited H-bonds (Fig. 6E). The free energy landscape and the conformations with the lowest energy were displayed (Fig. 7A–M; Additional file 1: Fig. S3A–B). Taken together, CA binds CDC42 and PTK2, and iso-CA binds CSF1R firmly. The results indicate that the transformed products may target CDC42, PTK2, and CSF1R directly.
Fig. 6
The dynamic stability of the ligand-target complexes. A The RMSD of ligands in compound-targets complexes show limited fluctuations in CA-CDC42, CA-PTK2, and iso-CA-CSF1R complexes but relatively large volatility in CA-CSF1R. B The RMSDs of receptors in compound-targets complexes show limited fluctuations in CA-CDC42, CA-PTK2, CA-CSF1R, and iso-CA-CSF1R complexes. C The RMSFs indicate that the residue-specific fluctuations of receptors are also stable, especially for the CA-CDC42 complex. D The Rgs of the four complexes are less fluctuated
Fig. 7
Exploration of the conformations with the lowest free binding energy. A The free energy landscape of CA-CDC42 during the 80-ns molecular dynamics simulation. 2D graph projects on the first two principal components (PC1 + PC2). Blue spots indicate the energy minima. B The overlapped graph of CA-CDC42 before (green) and after (blue) molecular dynamics simulation. C The low-energy conformation of CA-CDC42 is selected according to the free energy landscape. D The binding model of CA-CDC42 complex. Light green represents van der Waals, dark green represents hydrogen bonds, and pink represents hydrophobic interactions. E The free energy landscape of CA-PTK2 during the 80-ns molecular dynamics simulation. F The overlapped graph of CA-CDC42 before (green) and after (blue) molecular dynamics simulation. G The low-energy conformation of CA-PTK2 is selected according to the free energy landscape. H The binding model of CA-PTK2 complex. I The free energy landscape of iso-CA-CSF1R during the 80-ns molecular dynamics simulation. J The overlapped graph of iso-CA-CSF1R before (green) and after (blue) molecular dynamics simulation. K The low-energy conformation of iso-CA-CSF1R is selected according to the free energy landscape. L The binding model of the iso-CA-CSF1R complex. M The free energy landscape of CA-CSF1R
CSF1R is the receptor of macrophage-CSF (M-CSF) and interleukin (IL)-34, whose activation contributes to microglia/macrophage survival, proliferation, chemotaxis, and proinflammation [42,43,44,45]. Once phosphorylated, CSF1R activates downstream factors, CDC42 included, to rapidly reorganize the actin cytoskeleton and focal adhesions, resulting in microglia/macrophage movement [44, 46]. In addition, CSF1R also triggers proliferating signaling pathways, like PI3K/Akt, JNK, and ERK1/2 pathways [44]. CSF1R blockage reduces microglia number and inflammation [45, 47, 48]. PTK2, or focal adhesion kinase (FAK), is one of the primary controllers of cell mobility by regulating cytoskeletal or cell adhesion site dynamics [49, 50]. Moreover, it can also activate CDC42 [51]. A previous study suggests that PTK2 inhibition impairs microglia migration [52]. Thus, we speculate that forming CA-CDC42, CA-PTK2, and iso-CA-CSF1R complexes can inhibit microglia/macrophage migration, proliferation, and chemokine secretion after ICH. ASIV may not fulfill these effects alone, because CDC42, PTK2, and CSF1R are not its targets.
The orally administrated ASIV inhibits microglia/macrophage migration, proliferation, and chemokine secretionTo further verify the putative interactions, we validated the additional targets and biological processes of transformed ASIV derivatives on ICH. Thus, we orally administrated ASIV to mice after ICH induction (Fig. 8A). As previously reported, ASIV delivered in this way can be transformed by gut microbiota and liver sequentially, resulting in considerable concentrations of CA and iso-CA in the bloodstream [21, 22]. Then, microglia/macrophage migration and proliferation were evaluated. As predicted by the bioinformatic analyses above, these processes might be promoted by CDC42, PTK2, and CSF1R while suppressed by the transformed products, CA or iso-CA.
Fig. 8
The effects of orally administrated ASIV on microglia/macrophage proliferation, migration, and chemokine secretion after ICH. A Flow chart of the animal experiments. B H&E staining suggests that ASIV alleviates brain disorganization and inflammatory cell infiltration. C The statistic graph of Iba1 immunofluorescent suggests that ASIV dose-dependently reduces the number of perihematomal microglia/macrophages. D The statistic graph of Iba1 and PCNA double staining suggests that ASIV reduces the number of proliferating microglia/macrophage (green cells encircles red nuclei). E The representative images of Iba1 (green) and PCNA (red) immunofluorescent. F The representative images of Iba1 (green) and TNF-α (red) double staining indicate that TNF-α is predominantly expressed by microglia/macrophage. G The statistical graph of TNF-α immunofluorescent suggests that a high dose of ASIV declines TNF-α production. Dashed line: hematoma (B, E, F). Data are expressed as mean ± SD, n = 5. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. Scale bar = 50 μm
H&E staining showed that in the ICH group, the cells were disorganized with a looser extracellular matrix. Inflammatory cells were under infiltration (Fig. 8B, black circles). After treatment, the perihematomal disorganization and inflammation were reduced (Fig. 8B). Immunofluorescent staining suggested that the total microglia/macrophage and proliferating microglia/macrophage were significantly diminished in the ASIV-treated groups, especially for the high dose. The results implied that ASIV alleviates microglia/macrophage migration and proliferation (Fig. 8C–E). Moreover, a high dose of ASIV decreased the perihematomal TNF-α, the pro-inflammatory factor mainly secreted by microglia/macrophages in the brain (Fig. 8F–G).
As indicated by double immunofluorescent staining, the proliferation- and migration-promoting targets, CDC42 and CSF1R, were primarily expressed on Iba1-positive microglia/macrophage (Fig. 9B, E). They were significantly upregulated in the peri-hematoma brains after ICH. CDC42 and CSF1R were compromised after ASIV treatment (Fig. 9). It suggested that the binding of the transformed products on CDC42 and CSF1R potentially leads to their dysfunction and degradation. Moreover, the ASIV-reduced CDC42 and CSF1R might be responsible for the inhibited microglia/macrophage migration.
Fig. 9
The effects of orally administrated ASIV on CDC42 and CSF1R after ICH. A The representative double-staining images of Iba1 (green) and CSF1R (red) show that ASIV decreases CSF1R expression. B The enlarged images show that CSF1R is mainly expressed in microglia/macrophage. C The statistical graph indicates that ASIV suppresses the expression of CSF1R. D The statistical graph indicates that ASIV suppresses the expression of CDC42. E The enlarged images show that CDC42 (red) is highly expressed on Iba1-positive microglia/macrophage (green). F The representative double-staining images of Iba1 and CDC42 show ASIV decreases CDC42 expression. Data are expressed as mean ± SD, n = 5. *P < 0.05, ***P < 0.001, ****P < 0.0001. Scale bar = 50 μm
Our present results are supported by previous research, which demonstrates that intraperitoneally injection of ASIV or CA shifts microglia from a proinflammatory type to an anti-inflammatory one after brain ischemia [20, 23, 24]. In addition, a previous study applies a classic network pharmacology scheme to investigate the therapeutic mechanism of ASIV on ICH. It reports that orally administrated ASIV downregulates the levels of the master pro-inflammatory transcript factor, nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), in the post-ICH brain [17]. In comparison, our new strategy additionally suggests that ASIV can inhibit microglia/macrophage proliferation and migration after microbial and hepatic biotransformation.
CA but not ASIV suppresses microglia proliferation and migration in vitroThe in vivo study indicated that ASIV inhibited microglia/macrophage proliferation and migration after gut microbiota and liver biotransformation. However, the results could not distinguish the effects of the absorbed ASIV from those of the transformed products. Thus, we treated the microglia cell line (BV2) with ASIV or CA, one of the commercially available transformed products. The concentrations of ASIV and CA were determined according to the in vitro toxicity assay (Additional file 1: Fig. S4). Under LPS stimulation, CA but not ASIV suppressed BV2 proliferation (Fig. 10A–B) and migration (Fig. 10C–F). In addition, CA also reduced the expression of CDC42 and FAK in a dose-dependent manner (Fig. 10G–K).
Fig. 10
CA suppresses BV2 proliferation and migration, but ASIV does not. A CCK-8 shows that CA inhibits BV2 proliferation. B CCK-8 shows that ASIV doesn’t affect BV2 proliferation. C Scratch assay suggests that CA inhibits BV2 migration. D Scratch assay suggests that CA doesn’t inhibit BV2 migration. E Representative image of the scratch assay for CA. F Representative image of the scratch assay for ASIV. G Representative images of WB show that CA decreases FAK and CDC42 expressions. H The statistical graph shows that ASIV doesn’t affect the FAK level. I The statistical graph shows that CA decreases FAK expression. J The statistical graph shows that ASIV doesn’t affect CDC42 expression. K The statistical graph shows that CA suppresses the CDC42 level. Data are expressed as mean ± SD, n = 3. *P < 0.05, **P < 0.01, ***P < 0.001. Scale bar = 100 μm (E–F)
留言 (0)