
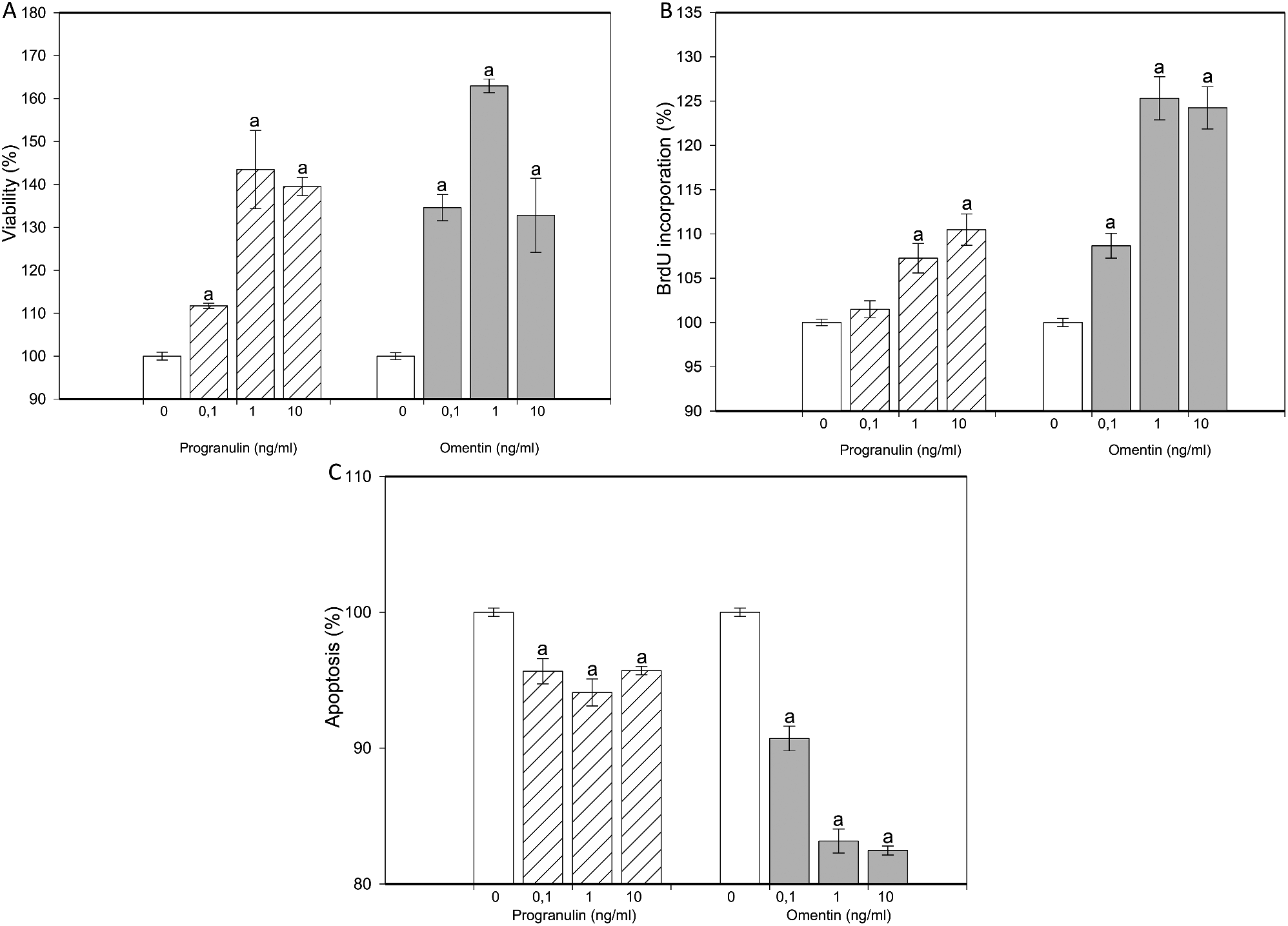
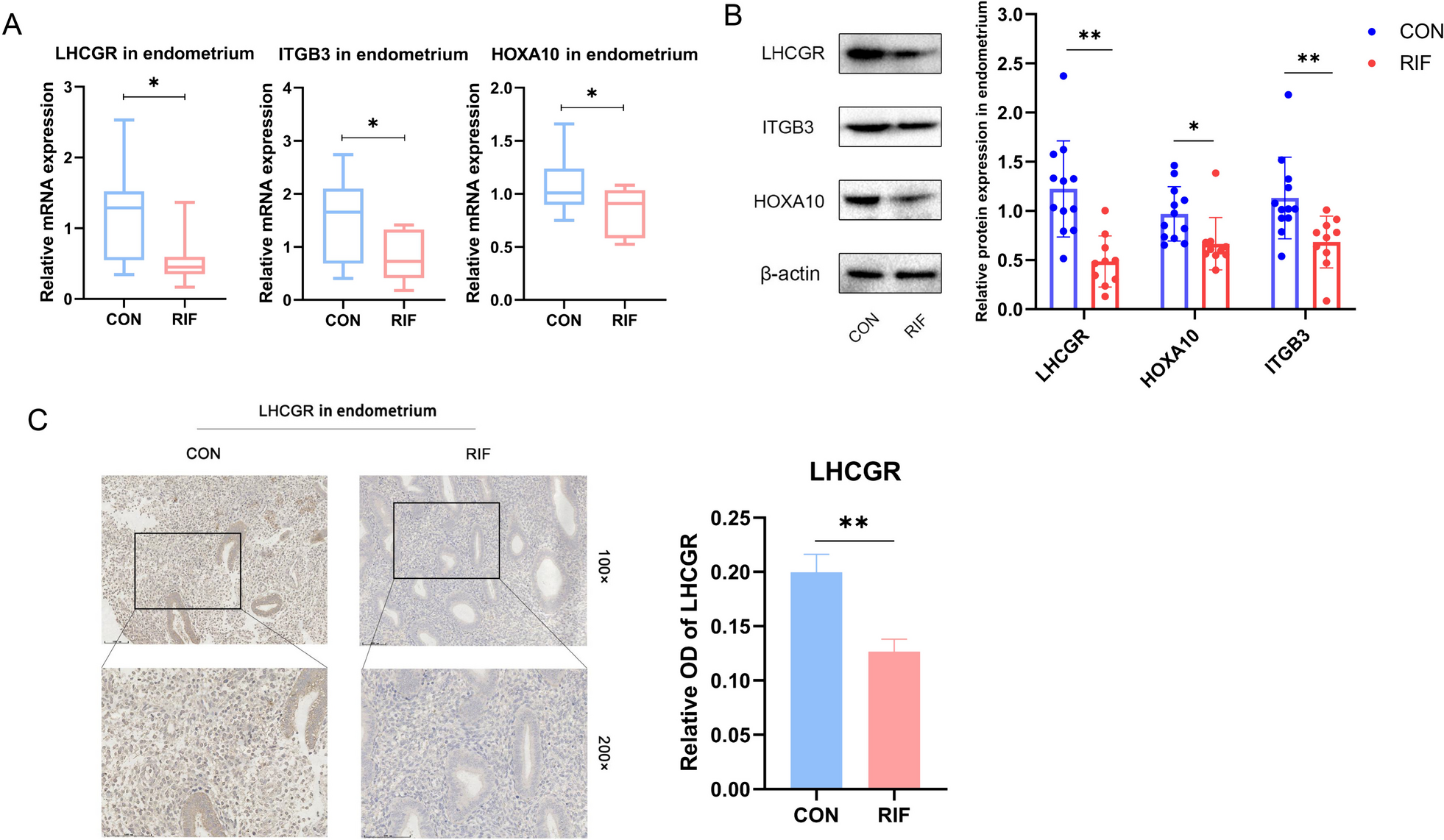
記住我
First, we investigated the mRNA levels of RA synthesizing enzymes (Raldh1, Raldh2, and Raldh3), RA receptors (Rarα, Rarβ, and Rarγ), retinoid X receptors (Rxrα, Rxrβ, and Rxrγ) and RA catabolizing enzymes (Cyp26a1, Cyp26b1, and Cyp26c1) by RT-qPCR in the mouse vaginas at 5 weeks, in which vaginal opening occurred. The result showed that Raldh2, Rarα, Rarγ, Rxrα, Rxrβ, Cyp26a1 and Cyp26b1 were highly expressed (Fig. 1A). In the following studies, we chose these highly expressed isoforms for localization analysis. Immunofluorescence staining revealed that RALDH2 was strongly stained in the vaginal epithelial cells of the cornified and the stromal region near basement membrane layers, and weakly stained in the vaginal epithelial cells of spinous layers (Fig. 1B). RARα, RARγ and CYP26A1 were stained in the vaginal epithelial cells of the spinous and cornified layers, and RARγ was also stained in the endothelial cells (Fig. 1B). CYP26B1 was strongly stained in the vaginal epithelial cells of the spinous layers, and weakly stained in the endothelial cells (Fig. 1B).
Fig. 1
The expression pattern of RA signaling molecules in the vaginas. (A) The mRNA levels of Raldhs, Rars, Rxrs and Cyp26s in mouse vaginas at 5 weeks. The mRNA values of Raldh1, Rarα, Rxrα and Cyp26a1 were set as 1, and those of other gens were normalized accordingly. (n = 3 independent experiments). Bars indicate the mean ± SEM. (B) Immunofluorescence staining for RALDH2, RARα, RARγ, CYP26A1 and CYP26B1 (red) in the vaginas at 5 weeks. The small white boxes indicate the enlarged areas as shown in the following images. Downwards arrows indicate epithelial cells of spinous layers, while leftwards and rightwards arrows indicate the epithelial cells of cornified layers and the stromal region near basement membrane layers, respectively. Asterisks (*) indicate endothelial cells. The nuclei were counterstained by DAPI (blue). The cells in the dashed white line box are vaginal epithelial cells. Scale bar, 100 μm. VL, vaginal lumen; E, epithelium. (C) The mRNA levels of Raldh2, Raldh3, Rarα and Rarγ in the vaginas at 2, 15, 21 and 30 dpp. The mRNA values of 2 dpp group were set as 1, and those of other groups were normalized accordingly. (n = 3 independent experiments). Bars indicate the mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001 vs. 2 dpp group. (D) The protein levels of RARα and RARγ in the vaginas at 2, 15, 21 and 30 dpp. GAPDH was used as a loading control. (n = 3 independent experiments). Bars indicate the mean ± SEM. *P < 0.05, **P < 0.01 vs. 2 dpp group
Next, we compared the expression of RALDH2, RALDH3, RARα and RARγ in the mouse vaginas at 2–30 dpp. The mRNA and/or protein levels of RALDH2, RALDH3, RARα and RARγ were significantly increased from 2 to 30 dpp (Fig. 1C and D), and those of RARγ reached a peak at 21 dpp (Fig. 1C and D). These results suggest that the increases of RALDH2, RALDH3, RARα and RARγ expression levels in the mouse vaginas are positively related to the occurrence of the vaginal opening.
Rarα deletion causes growth delay and progressive lethalityTo identify whether RA-RAR signaling was involved in regulating vaginal opening, we obtained the homozygous knockout mice (Rarα−/−) by crossing of heterozygote mice (Rarα+/−) with disrupted exon 4 of the Rarα gene on one chromosome (Fig. 2A). Genotyping was detected by common PCR (Fig. S1A). The Rarα knockout efficiency was confirmed by immunofluorescence and western blotting (Fig. 2B, C and Fig. S1B). Body size was comparable between Rarα−/− and wild type (WT) mice (Fig. S3A). However, the body weight of Rarα−/− females is significantly lower than WT mice from 2 to 7 weeks, but this difference disappeared after 8 weeks (Fig. 2D). The birth rate of Rarα−/− mice was 9.9% (26/263), which was significantly lower than the expected Mendelian rate of 25.0% (Fig. 2E), suggesting Rarα deletion causes partly embryonic lethality. 57.7% of Rarα−/− mice (15/26) died within 1 month of birth (Fig. 2F). These data demonstrate that Rarα deletion results in growth delay and progressive lethality.
Fig. 2
The effects of Rarα deletion on growth and survival. (A) The strategy of Rarα gene knock out (KO) by disrupting its exon 4 using CRISPR/Cas9 gene editing technology. (B, C) The detection of Rarα knockout efficiency in adult mouse vaginas by immunofluorescence (B) and western blotting (C) The nuclei were counterstained by DAPI (blue). Scale bar, 100 μm. GAPDH was used as a loading control. (n = 3 independent experiments.). Bars indicate the mean ± SEM. ***P < 0.001 vs. the WT group. (D) Body weight of Rarα−/− female mice (n = 6) and their WT littermates (n = 32) from 2 to 24 weeks. Bars indicate the mean ± SEM. ***P < 0.001 vs. the WT group. (E) Frequencies of Rarα genotypes in offspring from 30 Rarα+/− mating pairs were determined. Columns represent the percentage of the genotypes Rarα+/+ (black), Rarα+/− (green), and Rarα−/−(red), and two dashed red lines indicate expected frequencies based on Mendelian inheritance. The pup number for each genotype is shown in the columns. (F) Survival curve of Rarα+/+ (black), Rarα+/− (green), and Rarα−/− (red) from the newborn to adulthood
Rarα deletion could cause a closed vaginal phenotypeIn WT and Rarα+/− females, vaginal opening occurred around 30 dpp (Fig. S2). However, 25.0% of Rarα−/− females appeared with a closed vaginal phenotype with the swelling of the genital area (Fig. 3A, B and Fig. S3B), which prevented natural mating and caused infertility. In the remaining Rarα−/− female mice, the vaginal opening is normal (Fig. S2). Compared with WT females, the protein levels of RARγ had no change in the vagina of Rarα−/− females with vaginal closure, but significantly increased in those of Rarα−/− females with vaginal opening (Fig. S4). The Rarα−/− females at 3 weeks exhibited small vagina (Fig. S3A) and the wet weight of the Rarα−/− females (7.23 ± 1.67 mg; n = 3) was significantly lower than that of the WT mice (14.05 ± 0.91 mg; n = 4; P = 0.012). The Rarα−/− females with vaginal closure exhibited enlarged vagina and uterus (Fig. 3C). Compared to WT, the vaginal length of the Rarα−/− females with vaginal closure significantly reduced at 6 and 24 weeks (Fig. 3D), and the vagina usually had two lumens filled with fluid and the vaginal wall was thinner (Fig. 3E). In the Rarα−/− females with vaginal closure, the uterine horn of was significantly lengthened (Fig. 3D), the endometrial and myometrial layers were obviously thin, and the endometrial gland was absent (Fig. 3E), which is likely to be secondary to the mechanical pressure exerted by fluid accumulation [30].
Fig. 3
The effect of Rarα deletion on vaginal opening. (A, B) Representative pictures of the normal vaginal opening (white circle) in WT females and the vaginal closure (black circle) in Rarα−/− females at 6 weeks (A), and the frequency of the closed vagina in Rarα−/− females (B). The absolute numbers of females are indicated in the columns. (C, D) Representative female reproductive tracts (C) and the length of vagina and uterine horn (D) in Rarα−/− females with vaginal open, Rarα−/− females with vaginal closure and WT females at 6 and 24 weeks. Scale bar, 10 mm. (n = 3 independent experiments). Bars indicate the mean ± SEM. *P < 0.05, ***P < 0.001 vs. the WT group. (E) PAS-stained cross-sections for vaginas and uterus of Rarα−/− females with vaginal open, Rarα−/− females with vaginal closure and WT females. The small black boxes indicate the enlarged areas as shown in the following images. The yellow, black and white brackets indicate the thickness of vaginal epithelium (E), endometrium (EN) and myometrium (MY), respectively. Arrow, endometrial glands (Engl). Scale bar, 200 μm. VL, vaginal lumen; UL, uterine lumen
Rarα deletion blocks vaginal epithelial cell apoptosis and decreases Ctnnb1 expressionThe vaginal opening process is largely dependent on massive vaginal mucosal apoptosis, which is initiated by rapidly elevated levels of estrogen at puberty [7]. The deletion of Rarα significantly decreased the serum estradiol (E2) levels (Fig. 4A) and the ovarian mRNA levels of Cyp11a1, Cyp51, Ebp, Fdps, Msmo1, Mvk and Sqlen (Fig. S5). Compared with the WT females, the vagina mRNA (Raldh2, Raldh3, Ctnnb1, Bak and Bax ) and protein (Cleaved Caspase-3) levels were significantly decreased (Fig. 4B and C), but the mRNA levels of the anti-apoptosis gene Bcl-2 was significantly increased (Fig. 4C) in the Rarα−/− females with vaginal closure. Consistent with these, the percentage of vaginal epithelial cells with TUNEL- and Cleaved Caspase-3-positive staining was significantly decreased in the Rarα−/− females with vaginal closure. These results indicate that Rarα deletion decreases E2 levels and blocks vaginal epithelial cell apoptosis.
Fig. 4
The effect of Rarα deletion on vaginal epithelial cell apoptosis and Ctnnb1 mRNA levels. (A) The levels of serum estradiol (E2) in WT and Rarα−/− females at 30 dpp. (n = 3 independent experiments). Data are presented as mean ± SEM. ***P < 0.001 vs. the WT group. (B, C, D) The mRNA levels of Raldh2, Raldh3 (B), Bak, Bax and Bcl-2 (C) and the protein levels of Cleaved Caspase-3 (D) in the vaginas of Rarα−/− females with vaginal closure and WT females (non-estrus stage). The mRNA values of WT group were set as 1, and those of Rarα−/− group were normalized accordingly. GAPDH was used as a loading control. (n = 3 independent experiments). Data are presented as mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001 vs. the WT group. (E, F) Immunofluorescence stain of TUNEL and Cleaved Caspase-3 (green) (E), and the percentage of vaginal epithelial cells with TUNEL- and Cleaved Caspase-3-positive signals (green) (F) in each section in the vaginas of Rarα−/− females with vaginal closure and WT females (non-estrus stage). The nuclei were counterstained by DAPI (blue). Yellow arrows, apoptotic cells. Scale bar, 100 μm. (n = 3 independent experiments. The representative images are shown). Data are presented as mean ± SEM. ***P < 0.001 vs. the WT group. (G) The mRNA levels of Adamts18, Bac3, Bid, Bim, Ctnnb1, Epha1, Gpc3, Lhfpl2, Map3k1, Pax8 and Sema4d in the vaginas of Rarα−/− females with vaginal closure and WT females. The mRNA values of WT group were set as 1, and those of Rarα−/− group were normalized accordingly. (n = 3 independent experiments). Data are presented as mean ± SEM. ***P < 0.001 vs. the WT group
We next examined the mRNA levels of vaginal opening-related genes including Adamts18, Bbc3, Bid, Bim, Ctnnb1, Epha1, Gpc3, Lhfpl2, Map3k1, Pax8 and Sema4d. Compared with WT females, only the mRNA levels of Ctnnb1 were significantly decreased in the vaginas of Rarα−/− females with vaginal closure (Fig. 4G). These results indicate that the decrease of β-catenin in the vaginas of Rarα−/− females is involved in the closed vaginal phenotype.
RA treatment promotes β-catenin expression and apoptosis progression in the vaginasRARs form heterodimers with RXRs and act as ligand-regulated transcription factors through binding specific RA response element sequences (RAREs, (A/G)G(G/T)TCA), which are usually located in the promoters of target genes [31]. RA promotes β-catenin expression in hippocampal neural stem cells [32]. Therefore, we analyzed the 1-2000 bp (bp) of the Ctnnb1 promoter region and found three potential RARE-binding sites (Fig. S6A). The female mice at 12 dpp were initially subjected to ovariectomy, and then these females at 14 dpp were injected with RA (OVX-RA group) or with vehicle only (OVX-oil group. Figure 5 A). The mRNA levels of Ctnnb1, Bak and Bax were significantly increased, and those of Bcl2 were significantly decreased in the vaginas of OVX-RA group contrast to OVX-oil group (Fig. 5B). Consistent with this, the protein levels of β-catenin and BAX were significantly increased, and BCL2 was significantly decreased in OVX-RA group (Fig. 5C and D). The protein levels of active β-catenin were also significantly increased (Fig. 5E). These findings indicate that RA signaling promotes β-catenin expression and activity and the apoptosis of the vaginal epithelium, ultimately triggering the vaginal opening.
Fig. 5
The effect of E2 and RA treatment on vaginal epithelial cell apoptosis and Ctnnb1 mRNA levels in WT females. (A) The strategy of RA and E2 treatment in WT ovariectomized females. After ovariectomy (OVX), the females at 14 dpp received daily subcutaneous injections (s.c.) of E2 (0.1 µg/kg body weight. OVX-E2 group), RA (2.5 mg/kg body weight. OVX-RA group) or vehicle oil (OVX-oil group) until 17 dpp, and the females were sacrificed for experiments on vaginal tissue 24 h later. (B) The mRNA levels of Raldh2, Raldh3, Rarα, Rarγ, Rxrα, Rxrβ, Ctnnb1, Bak, Bax, and Bcl2 in the vaginas of OVX-E2 and OVX-oil groups, and the mRNA levels of Ctnnb1, Bak, Bax, and Bcl2 in the vaginas of OVX-RA and OVX-oil groups. The mRNA values of OVX-oil groups were set as 1, and those of other gens were normalized accordingly. (n = 3 independent experiments). Data are presented as mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001 vs. the OVX-oil group. (C, D, E) The protein levels of RARα, RARγ, β-catenin, active β-catenin, BAK, BAX and BCL2 in the vaginas of OVX-E2 and OVX-oil groups, and the protein levels of β-catenin, active β-catenin, BAK, BAX and BCL2 in the vaginas of OVX-RA and OVX-oil groups. GAPDH was used as a loading control. (n = 3 independent experiments). Data are presented as mean ± SEM. *P < 0.05, **P < 0.01 vs. the OVX-oil group. (F) The proposed model for RA-RAR signaling in vaginal opening
It is reported that RA biosynthetic enzymes and receptors are up-regulated by E2 treatment in rat prostates [33]. We analyzed the 1-2000 bp (bp) in the promoter regions of Raldh2, Raldh3 and Rarα, and found a potential estrogen response element sequence (EREs, AGGTCA) [34] in each of the promoter regions of these genes (Fig. S6B, C and D). Thus, we investigated the effect of E2 treatment on the expression of RA signaling molecules in the vaginas of ovariectomized females (OVX-E2 group. Figure 5 A). Compared with the OVX-oil group, the mRNA levels of Raldh2, Raldh3, Rarα, Rarγ, Rxrα and Rxrβ were significantly increased in the vaginas of OVX-E2 group (Fig. 5B). Consistent with this, the protein levels of RARα and RARγ were significantly increased in the vaginas of OVX-E2 group (Fig. 5C and D). Interestingly, the mRNA and protein levels of β-catenin, active β-catenin, BAK and/or BAX were also significantly increased, and those of BCL2 were significantly decreased in the vaginas of the OVX-E2 group in contrast to the OVX-oil group (Fig. 5B, C, D and E). These results suggest that E2 could promote vaginal opening by up-regulating RA signaling molecule expression in the vaginas.
留言 (0)