
記住我





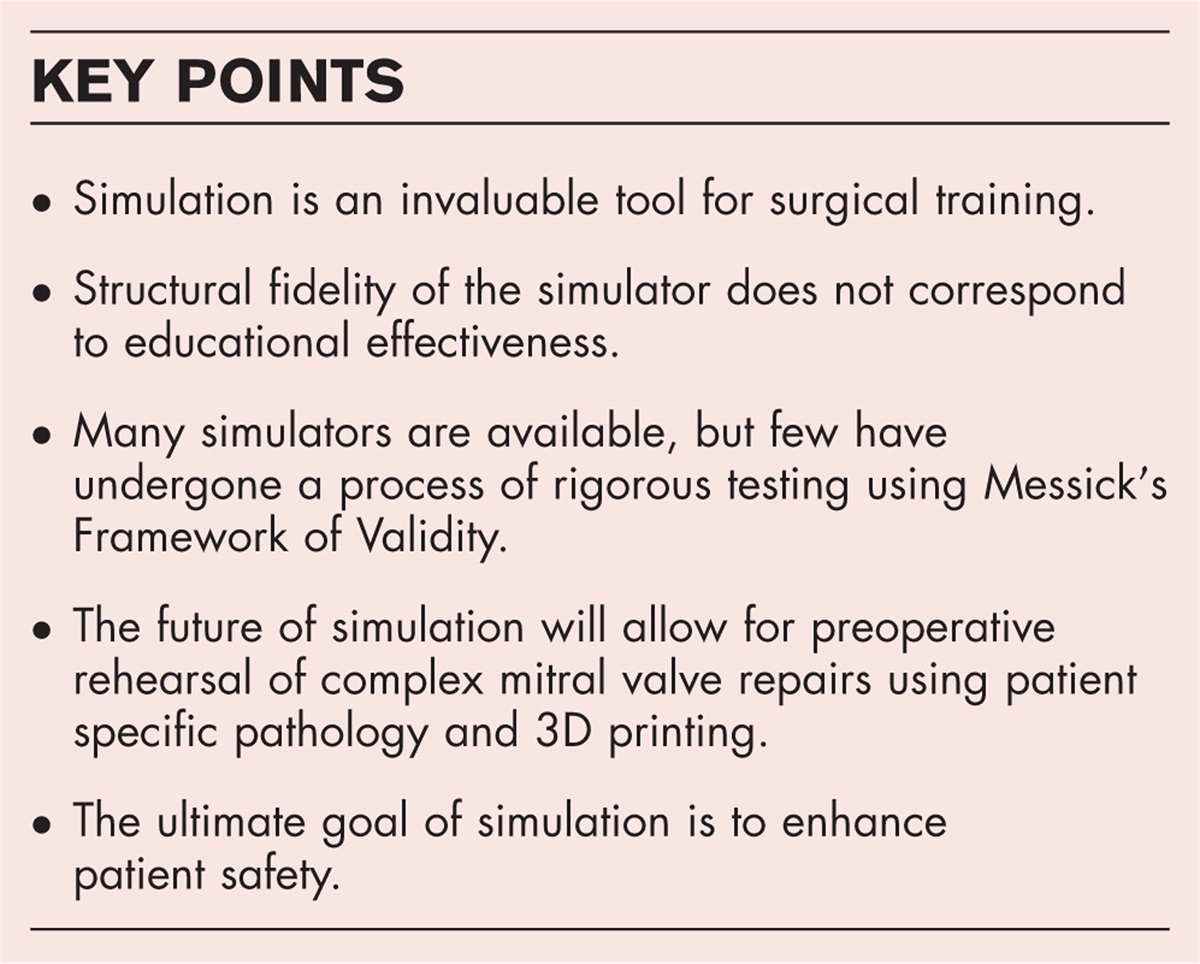

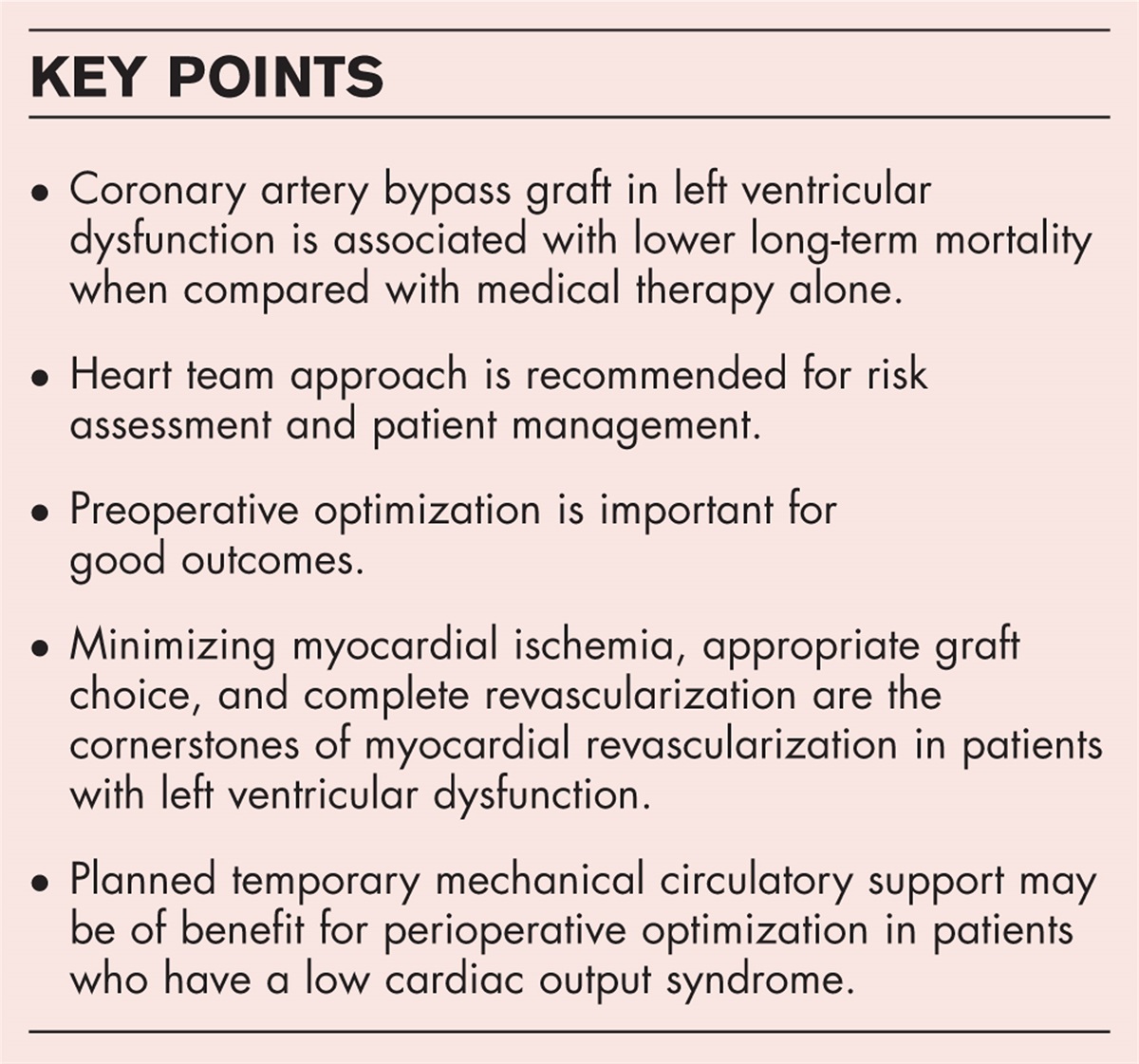
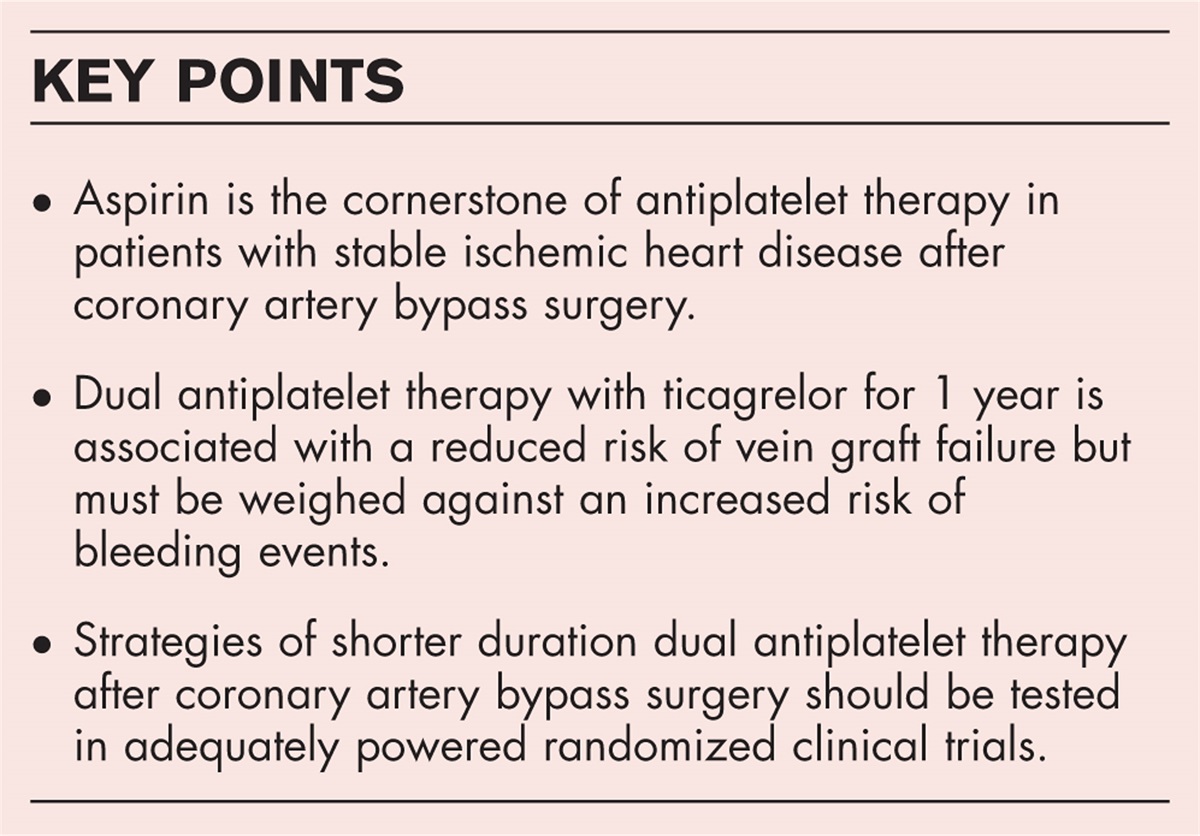
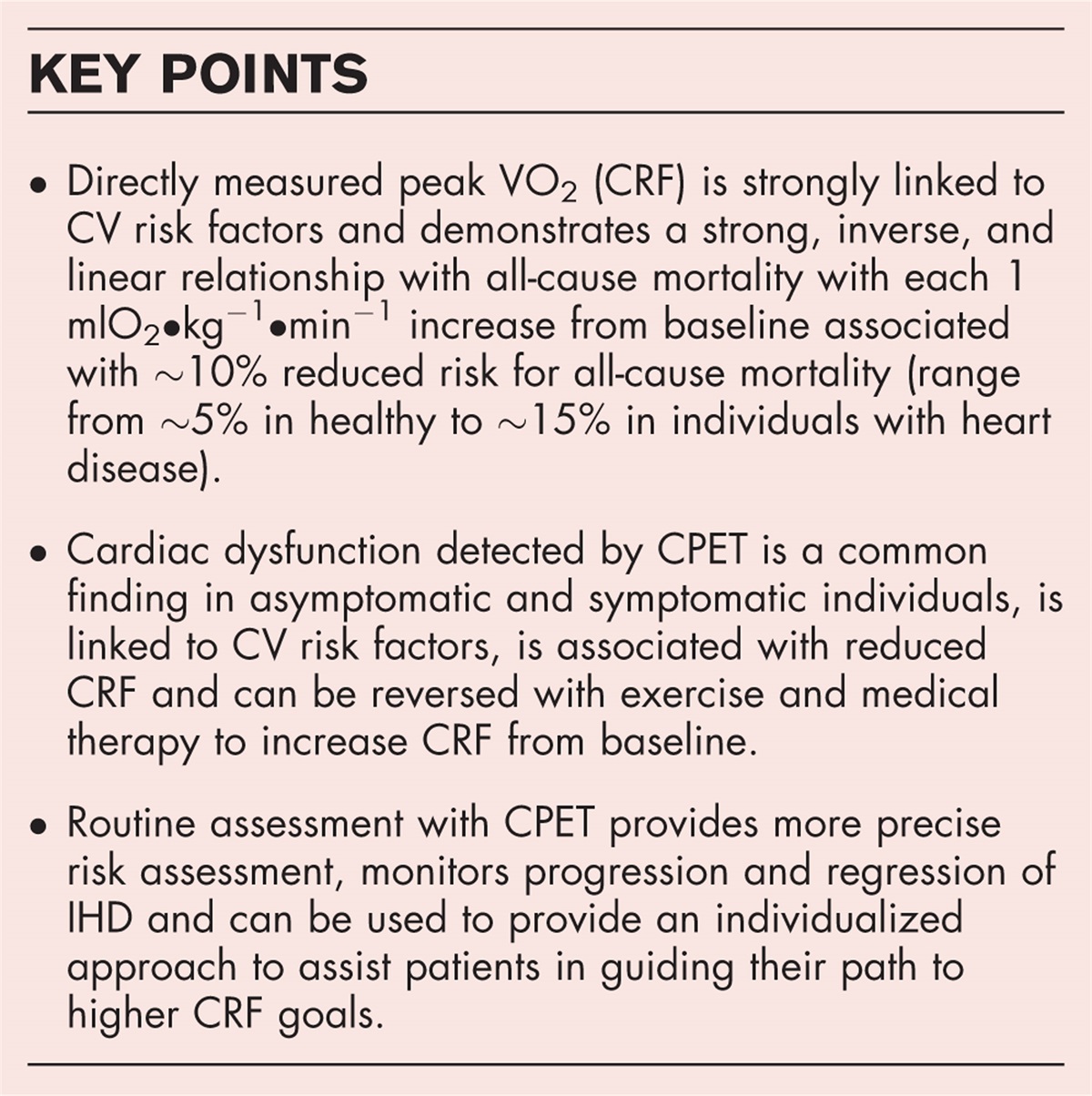
Structural variants are large genomic alterations of at least 50 bp in size. Over the last decades, they have emerged as a relevant genetic cause for several disorders, including intellectual disability, neuropsychiatric disorders, cancer and congenital heart disease [1▪]. This review specifically focusses on the contribution of structural variants in the development of aortic disease, including thoracic aortic aneurysm and dissection (TAAD), bicuspid aortic valve (BAV) related aortopathy and aortic stenosis.
 Box 1:
Box 1: no caption available
Types of structural variantsStructural variants can be subdivided into balanced and unbalanced rearrangements. Unbalanced structural variants include deletions and duplications, which are also called copy number variants (CNVs), as they increase or decrease the copy number of the affected region. CNVs can range in size from sub-microscopic to complete chromosomal aneuploidies [2]. Balanced rearrangements, on the contrary, include inversions and translocations and, as the name implies, do not change the copy number. Instead, they affect the orientation or location of a chromosomal segment without changing the copy number [3].
Compared with single nucleotide variants (SNVs), structural variants, and especially balanced rearrangements, are largely understudied, as they are more difficult to identify and require dedicated techniques or bioinformatic tools and algorithms. Although balanced structural variants are occasionally described in aortopathy literature, most research has been focused on CNVs.
Methods to detect structural variantsStructural variants can be detected using different methods. In the early days, large CNVs were detected using karyotyping, but the introduction of fluorescence in-situ hybridization (FISH) increased the resolution significantly. Nowadays, a wider range of techniques is used, which are classified into amplification, hybridization or sequencing-based approaches. Amplification-based approaches rely on the use of PCR, and include, for example, multiplex ligation dependent probe amplification (MLPA). Although MLPA is effective in the identification of deletions and duplications, a priori knowledge of the potentially involved gene is required. Alternatively, hybridization-based approaches, such as FISH, comparative genomic hybridization (CGH) and single nucleotide polymorphism (SNP) array, are often used for CNV identification and provides a significantly higher resolution than karyotyping. Both CGH and SNP array have the advantage of being hypothesis-free and genome-wide methods. With the introduction of next-generation sequencing (NGS), CNVs can now be detected at a much higher resolution. Using panel-based NGS and whole exome sequencing (WES), intragenic exonic CNVs are now identified on a routine basis, using dedicated bioinformatic methods. Both short and long-read whole genome sequencing (WGS) data can be used to detect more complex structural variants with high (single base pair) resolution. However, short-read WGS-based analysis is associated with a relatively high rate of structural variant miscalling due to errors in base calling, alignment and de-novo assembly that is especially challenging in repetitive regions. These shortcomings can be overcome using long-read WGS [4]; however, this approach is more expensive and low-throughput.
Thoracic aortic aneurysm and dissectionA progressive dilatation of the thoracic aorta leads to the development of thoracic aortic aneurysms (TAAs), which are often asymptomatic, but predispose to dissection and rupture. The latter are associated with high mortality rates [5]. Prophylactic surgery of TAA patients reduces the mortality rate to approximately 5%, but comes along with a relatively high risk of complications [6]. To date, mutations in over 40 genes have been described to cause TAA [7▪▪].
Using SNP array, CNVs in several genes have been shown to predispose to aneurysm development. A large cohort study of TAAD cases (n = 418 sporadic cases) by Prakash et al.[8] identified rare CNVs in 13% of sporadic TAAD cases, which was replicated in an independent cohort (n = 387 sporadic cases). In addition, considerably more CNVs (23%) were observed in familial TAAD patients (n = 88) than in sporadic (older) TAAD cases (17%) [8]. These CNVs were enriched for genes involved in smooth muscle cell adhesion and contractility. In an early-onset TAAD cohort with an average age of 20 years, the frequency of rare CNVs was even higher (i.e. 32%). This cohort was also enriched for BAV (55%), which underscores the role of BAV as a risk factor for early development of TAA [9], or alternatively indicates a shared genetic cause for BAV and TAA. The most frequently reported regions involved are 16p11.3 and 7q11.23.
Duplications of the 16p13.1 locus are reported to increase the risk for TAAD at least 12-fold [10]. This locus includes MYH11, a gene that encodes a contractile smooth muscle protein, called myosin heavy chain 11. In this gene, pathogenic TAAD-causing variants have already been described in approximately 1% of TAAD patients, especially when associated with persistent ductus arteriosus (PDA) [11]. Interestingly, the duplications did not always segregate with the TAAD phenotype in the families, indicating that the 16p13.1 duplication is a risk factor, which requires an additional hit, such as hypertension, the presence of BAV or another rare genetic variant, to lead to a TAAD phenotype. Indeed, patients with this duplication were more likely to carry a second rare CNV. Strikingly, they were also statistically more likely to suffer from aortic dissections [10]. As anticipated, MYH11 gene expression in affected aortic tissue was increased in patients with the 16p13.1 duplication compared with control aortas. However, no corresponding increase in protein levels was observed. It is hypothesized that the overexpression of MYH11 causes an imbalance of MYH11 to its chaperone, leading to MYH11 concentration-dependent dysfunction of smooth muscle cell contractility [10]. These recurrent 16p13.1 duplications are likely the result of the high number of interspersed segmental repeats, which often lead to duplications and deletions. Deletions of the same region have been described in a variety of complex mental disorders [12–14].
Another well known example is the triplication of the 7p11.23 locus, which includes the known supravalvular aortic stenosis (SVAS) gene ELN that codes for the protein elastin [15]. Cardiovascular abnormalities have also been reported in patients with a duplication of the 7q11.23 locus, but these seem to be less severe. At least one case of 7q1.23 duplication with supravalvular aortic aneurysms has been described in literature [8]. It remains to be determined how duplications and triplications of ELN lead to aortic aneurysms, but one hypothesis states that the excess of elastin could be responsible for the loss of resistance of elastic fibres to haemodynamic stress. The fact that blood pressure is higher in the supravalvular aorta could explain why aortic aneurysms occur specifically at this location [15]. Interestingly, ELN deletions and pathogenic variants leading to haplo-insufficiency of ELN cause SVAS (see Williams-Beuren syndrome below).
With the advent of NGS, CNVs can now be identified using a routine NGS panel, WES or WGS screening. In a large cohort of 810 TAAD patients, 8.1% received a genetic diagnosis using gene panel sequencing, of which 9.1% harboured a CNV in one of the coding regions of known TAA genes [MYLK (deletion, loss of initiation codon in smooth-muscle specific isoform), TGFB2 (in-frame deletion exon 4–7), SMAD3 (in-frame deletion exon 6), PRKG1 (in-frame deletion exon 3) and NOTCH1 (unbalanced translocation)]. In addition, a whole gene deletion of SCARF2 was detected. This gene was not associated with TAA before, but had previously been selected in view of a possible differential diagnosis of ‘congenital contractural arachnodactyly’ and ‘Van den Ende-Gupta syndrome’. The low diagnostic yield in this study is most likely attributed to increased offering of genetic testing at low thresholds in the Netherlands, where this study was executed [16]. Importantly, it should be noted that these CNVs can only be detected using dedicated bioinformatic tools and algorithms, which rapidly evolve over time. In a recent large genotype-phenotype study of paediatric Marfan syndrome (MFS; MIM #154700) patients (n = 373), which are often characterized by the presence of TAAD, three deletions of the entire FBN1 gene were identified, next to 13 deletions of one or more exons. In addition, two multiexon duplications were described [17–19]. In this paediatric cohort, 5% of patients harboured a pathogenic CNV in FBN1. FBN1 encodes fibrillin-1 that functions as a structural component in the extracellular matrix and regulates growth factor signalling pathways [20]. A study by Yamaguchi et al.[21] identified one multiexon deletion of COL3A1 (alpha 1 chain of type III collagen) in a patient with vascular Ehlers-Danlos syndrome (vEDS; MIM #130050). Although this specific patient did not present with aortic or arterial diseases (yet), these features are often present in vEDS [21]. Kempers et al.[22] described three patients with a deletion of varying size including both COL3A1 and COL5A2 (alpha 2 chain of type V collagen). These patients had a mild connective tissue phenotype without vascular abnormalities, which was hypothesized to corroborate the haploinsufficient disease mechanism [22]. In contrast, Meienberg et al.[23] identified an overlapping deletion in another patient with a complex phenotype with aortic rupture. Lastly, in 2017, we described two deletions of the X-linked gene BGN, in addition to three pathogenic SNVs, in men with a severe syndromic form of TAAD, termed Meester-Loeys syndrome (MRLS, MIM #300989). BGN encodes biglycan, a class I small leucine-rich proteoglycan involved in structural integrity and regulation of growth factor and cytokine signalling. The identified BGN deletions and pathogenic variants segregated with the aortic disease in the families [24]. These studies underscore the importance of routine CNV investigation in NGS data, as it significantly increases the genetic uptake.
More complex structural variants, such as inversions and translocations, are more difficult to identify, because it requires a combination of WGS and multiple algorithms and tools to accurately detect these types of structural variants. In 2022, more than 30 years after the identification of FBN1 as a causative gene for Marfan syndrome, the first inversion disrupting FBN1 has been described in a patient with classic MFS features. This discovery underscores the importance of routine structural variant investigation in TAAD patients [25▪].
Bicuspid aortic valveBicuspid aortic valve (BAV, MIM #109730) is the most common congenital heart defect (CHD), affecting 1–2% of the population. It is characterized by the presence of two aortic valve leaflets instead of three [26]. BAV patients have a greatly increased lifetime risk for the development of TAAD (i.e. 70–80%). Interestingly, BAV and TAAD segregate in pedigrees as different manifestations of a single underlying disorder. As already mentioned above, deleterious CNVs are highly enriched in early-onset TAAD patients, particularly in BAV patients, and affect genes that are known to cause aortic valve defects, aortic developmental abnormalities or TAA in humans or mice. These include deletions and duplications of ARSB, BGN, CXADR, EPHA3, HIC2, HOXA3 and SUMF1[9]. Of note, deletions and duplications of equal regions are associated with TAAD, suggesting that the development of the aorta is very sensitive to perturbations in the expression of implicated genes in either direction [9]. A truly largescale causative role for CNVs, however, has not been established for BAV. A study of Luyckx et al.[27] identified a potentially causative CNV involving TBX20, which encodes an important transcription factor for cardiovascular development. It was concluded that only 1.05% of BAV/TAA is caused by a potentially pathogenic CNVs. Instead, it has been hypothesized that certain CNVs could act as genetic risk factors for the development of BAV/TAA, rather than causing monogenic BAV/TAA disease.
Turner syndromeTurner syndrome (45,X) is a condition caused by the (partial) absence of one X-chromosome and occurs in approximately 1 : 2500 live births. It is characterized by congenital lymphedema, short stature, gonadal dysgenesis and cardiovascular malformations such as BAV, aortic coarctation, elongation of the transverse aortic arch and bovine aortic arch wherein the brachiocephalic artery shares a common origin with the left common carotid artery. Turner syndrome patients also have a high incidence of aortic root dilatation (10–32%) and aortic dissection [28–30]. It has been proposed that the aortic phenotype could be related to the absence of one copy of the X-linked BGN gene, an established X-linked TAAD gene, however, this still remains to be investigated [31].
Williams-Beuren syndromeWilliams-Beuren syndrome (WBS, MIM #194050) is a common microdeletion syndrome with an estimated prevalence of 1 : 7500 individuals [32]. It is caused by mispairing of low-copy DNA repeats at meiosis, resulting in a 1.5 Mb deletion of the 7q11.23 locus, which includes 25–28 genes [33,34▪▪]. WBS is characterized by cognitive impairment, dysmorphic facial features and SVAS [35,36]. Cardiovascular abnormalities are present in 50–88% of WBS patients [33], with the most common being SVAS, aortic arch hypoplasia and peripheral pulmonary stenosis [37]. A deletion of the ELN gene, located within the 7q11.23 locus, is responsible for the vascular and connective tissue features of WBS [32]. Of note, isolated SVAS is caused by translocations [38], gross deletions [39] and point mutations [40,41] that disrupt the elastin gene [42]. Interestingly, an inversion of the 7q11.23 locus is present in 6–7% of the general population and does not cause any symptoms [43].
Noncoding structural variantsThe involvement of coding deletions or duplications of known TAAD genes has already been characterized and these variants are now most often pursued in a routine NGS-based diagnostic setting. However, apart from coding CNVs, also noncoding CNVs affecting regulatory regions and more complex, balanced structural variants, could be involved in TAA development. Noncoding structural variants have been implicated in several diseases, but their role in the development of (syndromic) TAA remains largely elusive.
CONCLUSIONAlthough CNV identification in aortic (valve) disease has gained significant attraction over the past 15 years, there is still a lot to uncover. More complex structural variants, such as inversions, of known disease-causing genes are not yet routinely identified. This will most likely change in the near future, as WGS has become significantly cheaper and bioinformatic tools are being developed at a rapid pace. These developments allow an accurate identification of all types of structural variants. In addition, the use of WGS also facilitates the identification of structural variants in noncoding regions, which may affect the expression of relevant aortic (valve) genes. Although no noncoding CNVs have been described for aortic (valve) disease to date, these will undoubtedly be identified in the upcoming years.
AcknowledgementsNone.
Financial support and sponsorshipThis research was supported by funding from the Marfan Foundation (USA), the University of Antwerp (GOA, Methusalem-OEC grant ‘GENOMED’ 40709), the Research Foundation Flanders (FWO, Belgium 12X8520N). Dr Loeys holds a consolidator grant from the European Research Council (Genomia – ERC-CoG-2017–771945). Dr Loeys is a member of European Reference Network on rare vascular disorders (VASCERN project ID 769036).
Conflicts of interestNo conflict of interest is declared.
REFERENCES AND RECOMMENDED READINGPapers of particular interest, published within the annual period of review, have been highlighted as:
▪ of special interest
▪▪ of outstanding interest
REFERENCES 1▪. Audain E, Wilsdon A, Breckpot J, et al. Integrative analysis of genomic variants reveals new associations of candidate haploinsufficient genes with congenital heart disease. PLoS Genet 2021; 17:e1009679. 2. Mace A, Kutalik Z, Valsesia A. Copy number variation. Methods Mol Biol 2018; 1793:231–258. 3. Kosugi S, Momozawa Y, Liu X, et al. Comprehensive evaluation of structural variation detection algorithms for whole genome sequencing. Genome Biol 2019; 20:117. 4. Mahmoud M, Gobet N, Cruz-Davalos DI, et al. Structural variant calling: the long and the short of it. Genome Biol 2019; 20:246. 5. Hiratzka LF, Bakris GL, Beckman JA, et al. 2010 ACCF/AHA/AATS/ACR/ASA/SCA/SCAI/SIR/STS/SVM guidelines for the diagnosis and management of patients with Thoracic Aortic Disease: a report of the American College of Cardiology Foundation/American Heart Association Task Force on Practice Guidelines, American Association for Thoracic Surgery, American College of Radiology, American Stroke Association, Society of Cardiovascular Anesthesiologists, Society for Cardiovascular Angiography and Interventions, Society of Interventional Radiology, Society of Thoracic Surgeons, and Society for Vascular Medicine. Circulation 2010; 121:e266–e369. 6. Wong DR, Parenti JL, Green SY, et al. Open repair of thoracoabdominal aortic aneurysm in the modern surgical era: contemporary outcomes in 509 patients. J Am Coll Surg 2011; 212:569–579. discussion 79-81. 7▪▪. Rodrigues Bento J, Meester J, Luyckx I, et al. The genetics and typical traits of thoracic aortic aneurysm and dissection. Annu Rev Genomics Hum Genet 2022; 23:223–253. 8. Prakash SK, LeMaire SA, Guo DC, et al. Rare copy number variants disrupt genes regulating vascular smooth muscle cell adhesion and contractility in sporadic thoracic aortic aneurysms and dissections. Am J Hum Genet 2010; 87:743–756. 9. Prakash S, Kuang SQ, Regalado E, et al. Recurrent rare genomic copy number variants and bicuspid aortic valve are enriched in early onset thoracic aortic aneurysms and dissections. PloS one 2016; 11:e0153543. 10. Kuang SQ, Guo DC, Prakash SK, et al. Recurrent chromosome 16p13.1 duplications are a risk factor for aortic dissections. PLoS Genet 2011; 7:e1002118. 11. Zhu L, Vranckx R, Khau Van Kien P, et al. Mutations in myosin heavy chain 11 cause a syndrome associating thoracic aortic aneurysm/aortic dissection and patent ductus arteriosus. Nature genetics 2006; 38:343–349. 12. Ingason A, Rujescu D, Cichon S, et al. Copy number variations of chromosome 16p13.1 region associated with schizophrenia. Mol Psychiatry 2011; 16:17–25. 13. Ullmann R, Turner G, Kirchhoff M, et al. Array CGH identifies reciprocal 16p13.1 duplications and deletions that predispose to autism and/or mental retardation. Hum Mutat 2007; 28:674–682. 14. Jones I, Hamshere M, Nangle JM, et al. Bipolar affective puerperal psychosis: genome-wide significant evidence for linkage to chromosome 16. Am J Psychiatry 2007; 164:1099–1104. 15. Guemann AS, Andrieux J, Petit F, et al. ELN gene triplication responsible for familial supravalvular aortic aneurysm. Cardiol Young 2014; 25:712–717. 16. Overwater E, Marsili L, Baars MJH, et al. Results of next-generation sequencing gene panel diagnostics including copy-number variation analysis in 810 patients suspected of heritable thoracic aortic disorders. Hum Mutat 2018; 39:1173–1192. 17. Meester JAN, Peeters S, Van Den Heuvel L, et al. Molecular characterization and investigation of the role of genetic variation in phenotypic variability and response to treatment in a large pediatric Marfan syndrome cohort. Genet Med 2022; 24:1045–1053. 18. Baetens M, Van Laer L, De Leeneer K, et al. Applying massive parallel sequencing to molecular diagnosis of Marfan and Loeys-Dietz syndromes. Hum Mutat 2011; 32:1053–1062. 19. Proost D, Vandeweyer G, Meester JA, et al. Performant mutation identification using targeted next-generation sequencing of 14 thoracic aortic aneurysm genes. Hum Mutat 2015; 36:808–814. 20. Peeters S, De Kinderen P, Meester JAN, et al. The fibrillinopathies: new insights with focus on the paradigm of opposing phenotypes for both FBN1 and FBN2. Hum Mutat 2022; 43:815–831. 21. Yamaguchi T, Hayashi S, Hayashi D, et al. Comprehensive genetic screening for vascular Ehlers-Danlos syndrome through an amplification-based next-generation sequencing system. Am J Med Genet A 2023; 191:37–51. 22. Kempers MJ, Wessels M, Van Berendoncks A, et al. Phenotype of COL3A1/COL5A2 deletion patients. Eur J Med Genet 2022; 65:104593. 23. Meienberg J, Rohrbach M, Neuenschwander S, et al. Hemizygous deletion of COL3A1, COL5A2, and MSTN causes a complex phenotype with aortic dissection: a lesson for and from true haploinsufficiency. Eur J Hum Genet 2010; 18:1315–1321. 24. Meester JA, Vandeweyer G, Pintelon I, et al. Loss-of-function mutations in the X-linked biglycan gene cause a severe syndromic form of thoracic aortic aneurysms and dissections. Genet Med 2017; 19:386–395. 25▪. Pagnamenta AT, Yu J, Evans J, et al. Genomics England Research C. Conclusion of diagnostic odysseys due to inversions disrupting GLI3 and FBN1. J Med Genet 2022; 0:1–6. 26. Braverman AC, Guven H, Beardslee MA, et al. The bicuspid aortic valve. Curr Probl Cardiol 2005; 30:470–522. 27. Luyckx I, Kumar AA, Reyniers E, et al. Copy number variation analysis in bicuspid aortic valve-related aortopathy identifies TBX20 as a contributing gene. Eur J Hum Genet 2019; 27:1033–1043. 28. Lin AE, Lippe BM, Geffner ME, et al. Aortic dilation, dissection, and rupture in patients with Turner syndrome. J Pediatr 1986; 109:820–826. 29. Bondy CA. Aortic dissection in Turner syndrome. Curr Opin Cardiol 2008; 23:519–526. 30. Galian-Gay L, Rodriguez-Palomares JF. Turner syndrome and aortic complications: more benign than previously thought. Heart 2022; 109:82–83. 31. Meester JAN, De Kinderen P, Verstraeten A, Loeys BL. The role of biglycan in the healthy and thoracic aneurysmal aorta. Am J Physiol Cell Physiol 2022; 322:C1214–C1222. 32. Stromme P, Bjornstad PG, Ramstad K. Prevalence estimation of Williams syndrome. J Child Neurol 2002; 17:269–271. 33. Kruszka P, Porras AR, de Souza DH, et al. Williams-Beuren syndrome in diverse populations. Am J Med Genet A 2018; 176:1128–1136. 34▪▪. Kozel BA, Barak B, Kim CA, et al. Williams syndrome. Nat Rev Dis Primers 2021; 7:42. 35. Williams JC, Barratt-Boyes BG, Lowe JB. Supravalvular aortic stenosis. Circulation 1961; 24:1311–1318. 36. Beuren AJ, Apitz J, Harmjanz D. Supravalvular aortic stenosis in association with mental retardation and a certain facial appearance. Circulation 1962; 26:1235–1240. 37. Collins RT 2nd. Cardiovascular disease in Williams syndrome. Curr Opin Pediatr 2018; 30:609–615. 38. Curran ME, Atkinson DL, Ewart AK, et al. The elastin gene is disrupted by a translocation associated with supravalvular aortic stenosis. Cell 1993; 73:159–168. 39. Ewart AK, Jin W, Atkinson D, et al. Supravalvular aortic stenosis associated with a deletion disrupting the elastin gene. J Clin Invest 1994; 93:1071–1077. 40. Li DY, Toland AE, Boak BB, et al. Elastin point mutations cause an obstructive vascular disease, supravalvular aortic stenosis. Hum Mol Genet 1997; 6:1021–1028. 41. Tassabehji M, Metcalfe K, Donnai D, et al. Elastin: genomic structure and point mutations in patients with supravalvular aortic stenosis. Hum Mol Genet 1997; 6:1029–1036. 42. Metcalfe K, Rucka AK, Smoot L, et al. Elastin: mutational spectrum in supravalvular aortic stenosis. Eur J Hum Genet 2000; 8:955–963. 43. Hobart HH, Morris CA, Mervis CB, et al. Inversion of the Williams syndrome region is a common polymorphism found more frequently in parents of children with Williams syndrome. Am J Med Genet C Semin Med Genet 2010; 154C:220–228.
留言 (0)