
記住我

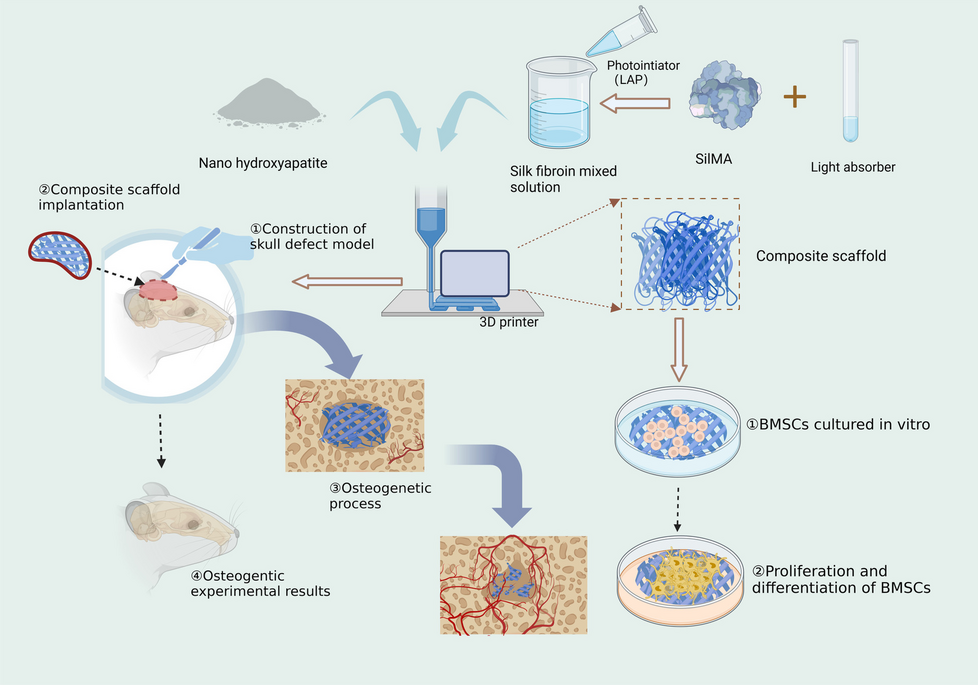
We present a newly developed and simple in vitro culture platform that can be applied to investigate the effect of the migratory behavior of cells on the spatial self-organization of hiPSCs. Figure 1 depicts the structure of the PDMS-based culture system embedded with the ring magnet, used in this study. The proposed PDMS culture system is constructed based on a magnetic ring embedded in a resin with high gas permeability. A magnetic ring embedded in PDMS was fabricated using a standard soft lithography technique. To facilitate easy removal of the ring from the culture surface, this culture system consists of two complementary parts composed of a simple removable PDMS ring with an embedded magnetic ring and a stainless-steel plate, allowing the attachment and detachment of two parts repeatedly. The ring is composed of a closed culture vessel with a height and internal diameter of 1.3 and 2.5 mm, respectively. This method allows single colony formation from multiple cells under space restriction and can induce cell migration by a simple method using rings that are placed before seeding cells and removed after cell colony formation.
Fig. 1
Establishment of ring culture system and experimental design. A Ring fabrication diagram showing the structure of PDMS-based culture system embedded with ring magnet. B Experimental design for hiPSC colony formation (Step 1) and self-organization of spatial patterns (Step 2) within the hiPSC colony with or without ring-barrier. The barrier made from detachable ring culture system is placed on culture surface and cells are seeded within this area; upon confluency, the barrier is removed, which results in a center and edge of the colony, a cell-free space that allows cell migration. In Step 2, the cell migration was monitored and the spatial self-organization within the hiPSC colonies was compared under culture conditions with and without HA exposure
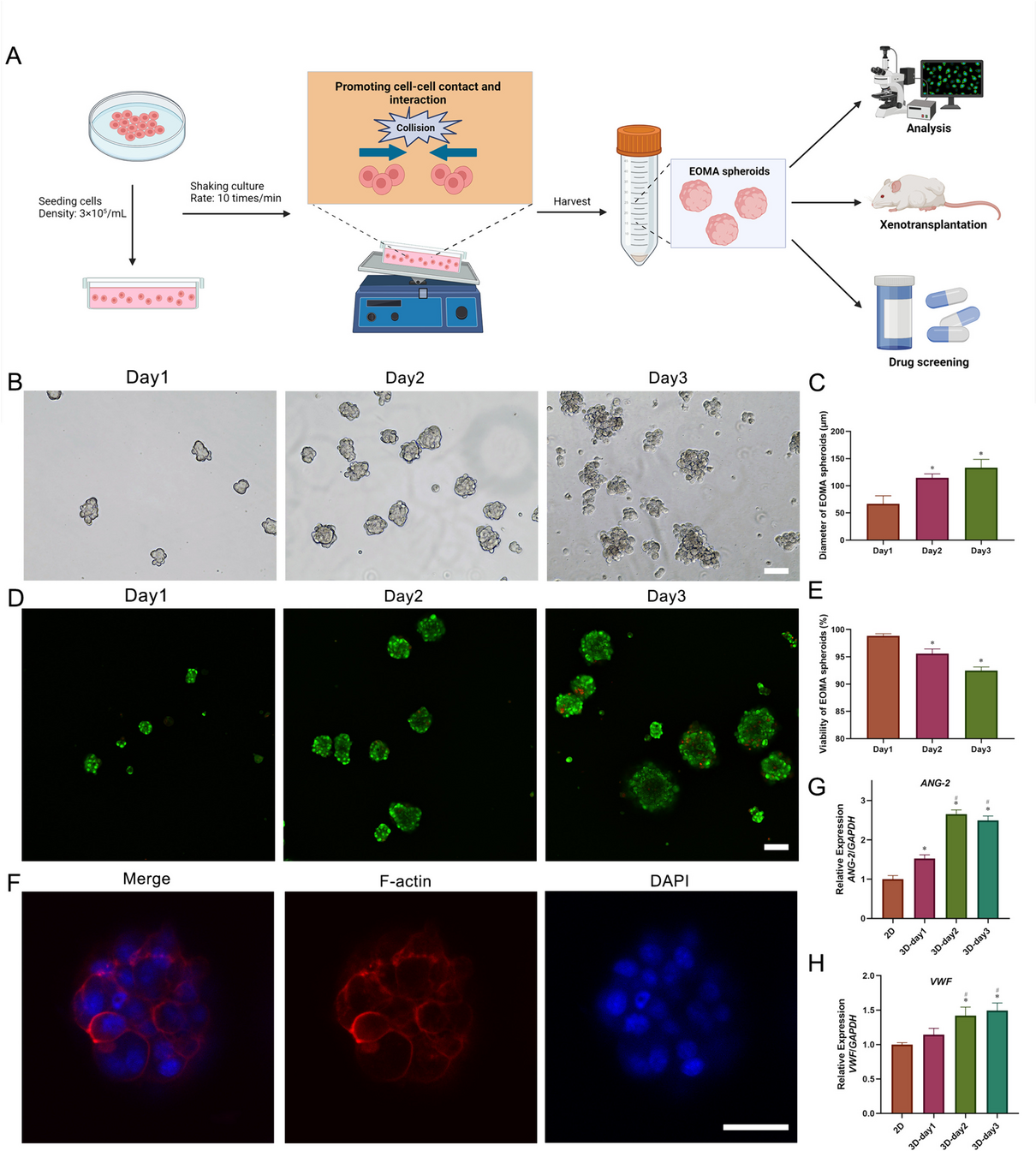
Characterization of single colony formation by hiPSC population in a ring culture systemTo study the effect of physical barriers on hiPSC colony formation by cell populations, we present a method that utilizes a PDMS-based ring culture system to spatially restrict cell adhesion to the underlying substrate (Fig. 2A). A magnetic ring was placed on the iMatrix-coated culture surface, and cells were seeded and cultured under culture conditions to maintain pluripotency. After seeding the cultures at a high density, the cells grew rapidly to generate multiple single cell-derived colonies of 2.5 mm diameter. Within a colony, cells at the edge grew faster than those at the center and had a higher local density around this barrier in keeping the barrier made from the ring culture system on day 10 (Fig. 2B, C). The cells fully grew as a tightly packed colony and well-defined edge in a confined space of the ring culture system. The edge of the colony showed a multilayered appearance along the sidewall of the ring culture system in a closed space.
Fig. 2
Spatial self-organization within the hiPSC colony with or without the ring-barrier. A Experimental design for hiPSC colony formation (Step 1) and self-organization of spatial patterns (Step 2) within the hiPSC colony with or without the ring-barrier. The barrier made from detachable ring culture system is placed on culture surface and cells are seeded within this area; upon confluency, the barrier is removed, which results in a center and edge of the colony, a cell-free space that allows cell migration. In Step 2, the cell migration was monitored and the spatial self-organization within the hiPSC colonies was compared under culture conditions with and without HA exposure. B Representative morphologies at different time points during spatial self-organization within hiPSC colony with or without ring-barrier are shown. The hiPSC colonies were compared after further exposure to HA in culture conditions without the ring-barrier. Colony diameter, 2.5 mm. Scale bar, 500 μm. C Representative images for cell morphology at the center and edge of the hiPSC colony in the image (B). All experiments were repeated independently at least three times with similar results
To examine the effect of collective cell migration at the edge of the spatially confined hiPSC colony, the ring culture system was removed on day 0 and cultured in a KSR-based medium without the addition of exogenous supplements to study the effect of collective cell migration alone. From days 1 to 5, cells at the colony edge flattened and started to migrate away from the colony (Fig. 2B, C). By day 7, the migrating cells presented flattened, elongated, and fibroblastic morphology. In contrast, cells in the central region of the original colony remained highly proliferative and retained epithelial morphology. Multilayer structures were formed at the boundary between the inside and outside of the ring-barrier.
The spatial pattern of cells within the hiPSC colony highly depends on cell–cell interactions. Since E-cadherin is known to be linked to the actin cytoskeleton, cell–cell adhesion further enhances collective cell migration [23,24,25]. To further confirm the roles of collective cell migration on the self-organization of cells within the hiPSC colony, botulinum hemagglutinin (HA), an E-cadherin adhesion-blocking agent was added to hiPSC colony [26, 27]. On day 1 after HA exposure for 24 h, the contact between the cells was lost and some cells appeared highly rounded up (Fig. 2B, C). However, it was completely restored by day 7. The cells at the colony edge showed a flat and elongated fibroblast morphology. Taken together, these results suggest that removal of the ring barrier is sufficient to induce self-organization of hiPSCs through collective cell migration at the colony edge, which suggests that stable adherens junctions during collective cell migration are necessary for self-organization of spatial patterning hiPSC colony.
Characterization of self-organization of spatial patterning hiPSCs with or without ring-barrier in the absence of exogenous differentiation factorsTo define the relative contributions of mechanical forces on the self-organization of cells, we reconstituted hiPSCs colonies in a ring culture system under culture conditions of pluripotency maintenance. We performed fluorescence staining of phosphorylated myosin light chain (pMLC), which interacts with F-actin bundles to generate contractile force in cultured cells. On day 0, a three-dimensional (3D) reconstruction of the colonies based on staining of F-actin pMLC revealed a multilayered appearance alongside the internal walls inside a confined space made from the ring culture system (Fig. 3A). pMLC colocalized with visibly pronounced F-actin development at the edges of the self-organized hiPSC colony. Quantification of the co-localization of pMLC with F-actin also revealed a peak near the colony edge, further suggesting enriched actomyosin activity at the edge near the barrier of the ring culture system (Fig. 3B). Considering the observed edge retraction upon ablation, this suggests the presence of mechanical tension at the overhanging edges of the self-organized hiPSC colony. Immunostaining of pluripotency markers OCT3/4 and SOX2 revealed loss of OCT3/4 expression in cells at the colony edge, while SOX2 expression was maintained in cells localized throughout the colony (Additional file 1: Fig. S1A). In addition, immunostaining of Ki67, a cell proliferation marker was carried out, and this indicated that cells at the colony edge still had increased proliferation as compared to cells at the colony center (Additional file 1: Fig. S1B). Taken together, these results suggest that the involvement of mechanical tension at the edge of a self-organized hiPSC colony can in turn influence the pluripotency state even under culture conditions that maintain pluripotency.
Fig. 3
Characterization of spatial self-organization in the hiPSC colony with and without the ring-barrier. A Representative images in the XY plane showing pMLC and F-actin within the hiPSC colony in cultures with (a–c) the ring-barrier at day 0, and with (d–f) or without (g–i) the ring-barrier at day 2. The hiPSC colonies were compared after further exposure to HA in culture conditions without the ring barrier (j–l) at day 2. Images show the whole colony. Scale bar, 500 μm. Panels a1–l1 are the tomograms sectioned at the XZ plane (yellow dashed lines) in panels a-l. Red dashed lines indicate location of the ring culture system. Scale bar, 50 μm. B Quantification of staining intensity of pMLC and F-actin signals across the hiPSC colony with or without ring-barrier. Data are average ± S.D. of 5 cross-sections from the colonies. Significance was determined by one-way ANOVA with Tukey's test (**P < 0.01, *P < 0.05). All experiments were repeated independently at least three times with similar results
To better understand this process, we examined the 3D spatial structure of F-actin and pMLC in the colonies cultured in a KSR-based medium without the addition of exogenous supplements. In cultures with a ring-barrier, extended actin filaments assembled into network-like structures, particularly at sites of intercellular connections within a compact cell colony on day 2 (Fig. 3A and Additional file 2: Movie S1). The cells at the colony center formed adherent monolayers of polygonal cells that possessed a defined vertical organization to the actin cytoskeleton. F-actin was highly enriched at the colony edges with clearly visible bundles compared with cells at the colony center. In addition, the cells at the colony center exhibited cortical actin network but lacked visible F-actin bundles. F-actin and pMLC were more enriched closer to the apical side of cells at the edge of the colony, while no enrichment was observed at the center, indicating that such a polarized distribution facilitates the balance of stresses arising from actomyosin contractility. The pMLC staining intensity in F-actin bundles also revealed a peak at the edge near the barrier of the ring culture system (at the location of the ring barrier) (Fig. 3B).
In cultures without a ring-barrier, actin fibers increased in cells within the hiPSC colony through the regulation of pMLC during collective cell migration on day 2 (Fig. 3A and Additional file 3: Movie S2). The multilayered cells at the edge near the barrier of the ring culture system (at the location of the ring barrier) were monolayered by cell migration at the edge of the colony. Actin stress fibers were abundant along the apical and basal sides of the cells in monolayers. They exhibited epithelial-like characteristics with specialized cell–cell junctions anchored by apical bundles of actin filaments. Moreover, actin-based plasma membrane protrusions called lamellipodia and filopodia were concentrated at the leading edge of the colony. Compared with cells at the colony center, cells at the ring barrier and colony edge exhibited pronounced expression of pMLC that interacted with F-actin bundles to generate contractile force. On the apical side, the thick circumferential belt of F-actin was interconnected between the cells and strongly colocalize with pMLC. Both F-actin and pMLC were observed at the same level of staining intensity from the center to the edge of the colony (Fig. 3B).
In cultures where the ring barrier was removed and exposed to HA, the cells within colonies had a markedly round morphology compared to that of untreated cells, confirming an increase in shrinkage of the apical surface of the cells (Fig. 3A and Additional file 4: Movie S3). pMLC strongly colocalized with F-actin at the apical surface of cells at the colony center. However, the actin fibers increased at the colony edge, and a perpendicular alignment of cells and actin fibers with respect to the direction of stretching was observed. In addition, cells displayed relatively uniform staining intensity of both F-actin and pMLC (Fig. 3B). Taken together, these results suggest that the changes of mechanical tension through collective cell migration at the colony edge may trigger the transition of pluripotency state even in the absence of exogenous supplements.
To determine the pluripotency status and lineage commitment within the hiPSC colony, we immunostained the pluripotency markers OCT3/4 and SOX2, ectodermal marker PAX6, definitive endoderm marker SOX17, and mesoderm/primitive streak marker BRACHYURY on day 7. Immunostaining of pluripotency marker OCT3/4 revealed that OCT3/4 expression was maintained localized in cells through the colony in culture with or without a ring-barrier, while loss of OCT3/4 expression was throughout the colony in culture without a ring-barrier through disruption of cadherin-based cell junctions with HA exposure (Fig. 4A). SOX2 is a core transcription factor that is maintained during ectodermal differentiation but downregulated during meso–endodermal differentiation [5]. The SOX2 and PAX6 were expressed throughout the spatially confined hiPSC colony within the ring culture system. However, in cultures without a ring-barrier, the cells at the colony center expressed SOX2 and PAX6, marking the prospective ectoderm. The rings at progressively larger radii expressed BRACHYURY, SOX17, and CDX2, marking the emergence of mesoderm, endoderm, and extra-embryonic trophoblasts, respectively. These findings suggest a two-part mechanism whereby spatially restricted high cell-adhesion tension initiates at the colony edge and then feeds forward to drive collective cell migration that induces meso–endodermal specification to reinforce the gastrulation-like phenotype even in the absence of exogenous differentiation factors (Fig. 5). Accordingly, inhibition of collective cell migration through disruption of cadherin-based cell junctions with HA exposure was restricted to ectodermal differentiation only and did not develop other lineage-differentiated cells.
Fig. 4
Characterization of spatial self-organization within the hiPSC colony with or without the ring-barrier in the absence of exogenous differentiation factors. Immunofluorescent images of a pluripotency markers (OCT3/4 and SOX2) and differentiation markers including ectoderm (PAX6), mesoderm (BRACHYURY), endoderm (SOX17), and extra-embryonic trophoblast (CDX2). The hiPSC colonies were compared after further exposure to HA in culture conditions without the ring-barrier. Red dashed lines indicate location of the ring culture system. Scale bar, 500 μm. All experiments were repeated independently at least three times with similar results
Fig. 5
Schematic of our hypothesis on the mechanism by which collective cell migration induced by the removal of ring-barrier induces the spatial self-organization into gastrulation-like phenotype. A Comparison of spatial self-organization within the hiPSC colony with or without the ring-barrier in the absence of exogenous differentiation factors. In the first step, hiPSCs self-assemble into a colony primarily through cell–substrate and cell–cell adhesion. In the second step, by removing the ring-barrier, the hiPSC colony spontaneously differentiated into SOX17+/BRACHYURY+ gastrulation-like structures through collective cell migration without exogenous supplements. The hiPSC colonies were compared after further exposure to HA in culture conditions without the ring-barrier. B A detailed description of the spatial self-organization process within the hiPSC colony through collective cell migration in the absence of exogenous differentiation factor
Characterization of self-organization of spatial patterning in hiPSCs with or without ring culture systems under the influence of endodermal differentiation conditionTo investigate the effect of collective cell migration on self-organization within the hiPSC colony, we tested the differentiation potential of the cells as they transitioned from undifferentiated to endodermal progenitors by exchanging the endodermal induction media after 3 days of hiPSC colony formation. We performed time-lapse observations of the hiPSC differentiation culture after the removal of the ring-barrier on day 0. The cells at the colony edge collectively migrated towards the newly available free space, resulting in colony patterns that arise primarily through cell migration (Additional file 5: Movie S4). The cells at the colony edge with distinct boundaries began to migrate outwards into free space and remained on the perimeter of the colony to eventually form a band of differentiation. When switching to the endodermal induction media on day 0, in cultures with the ring-barrier, OCT3/4 was still expressed throughout the colony by day 7, while SOX17 and FOXA2 were expressed in a few cells at the colony edge (Fig. 6). In cultures without a ring barrier, however, the cells at the center expressed OCT3/4, whereas the OCT3/4 was completely absent at the colony edge. The SOX17 and FOXA2 were expressed in cells at the colony edge. The above results argue that in cultures without the ring-barrier, cells integrate information from the exogenous differentiation factors to create an endoderm pattern within the colony. We found that in a conventional culture with endodermal differentiation media, the cells were a mixture of dense and relatively dispersed parts (Fig. S2). Interestingly, the OCT3/4 expression was still observed in tightly packed regions with cells, whereas the SOX17 expression was completely absent in the densely packed cells. This result is consistent with current research showing that the cells at the center region within the spatially confined hiPSC colony within the ring culture system retain the expression of OCT3/4. We hypothesized that the cell fate decision in the second step of the differentiation induction depends on EMT due to collective cell migration at the colony edge capable of outward migration. Cell migration in differentiating hPSCs was tightly linked and had been attributed to the process of EMT, which was characterized by reduced E-cadherin and increased N-cadherin expression [4, 13, 14]. To investigate the mechanisms of the effect of cell migration-driven EMT in cultures with and without the ring-barrier under the influence of endodermal differentiation condition, we performed immunostaining for two cell–cell adhesion-associated proteins, E-cadherin and N-cadherin, and a cell-substrate adhesion-associated protein, paxillin at day 7. In cultures without the ring-barrier, E-cadherin expression appeared at the colony center, whereas N-cadherin was expressed at the colony edge (Fig. 7A). In contrast, E-cadherin was expressed at cell–cell contacts within colonies in cultures with the ring-barrier. Cell nuclei at the boundary region were partially overlapped and detached from the substrate. In this regard, the relationship between switching from E-cadherin to N-cadherin was shown for the formation of a boundary separating undifferentiated and differentiated cells by collective cell migration at the colony edge under the influence of endodermal differentiation conditions.
Fig. 6
Characterization of spatial self-organization within the hiPSC colony with or without the ring-barrier using the ring culture system under the influence of endodermal differentiation conditions. A Experimental design for endodermal differentiation based on the ring culture system. B Representative morphologies are shown at different time points during endodermal differentiation within the hiPSC colony with or without the ring-barrier. Scale bar, 500 μm. C Representative images in the XY plane showing expression of pluripotency (OCT3/4) and endodermal (FOXA2, SOX17) markers. Images are shown as one quarter of a colony. Panels a1–r1 are the tomograms sectioned at the XZ plane (yellow dashed lines) in panels a-r. White dashed lines indicate location of the ring culture system. Nuclei were stained with DAPI. Scale bar, 50 μm. All experiments were repeated independently at least three times with similar results
Fig. 7
Characterization of spatial self-organization within the hiPSC colony with or without the ring-barrier in the absence of exogenous differentiation factors. A Representative images in the XY plane showing the expression of E-cadherin and N-cadherin within the hiPSC colony. Panels a1–l1 are the tomograms sectioned at the XZ plane (yellow dashed lines) in panels a-l. Nuclei were stained with DAPI. Scale bar, 50 μm. B Representative images in the XY plane showing the expression of paxillin within the hiPSC colony. Panels m1–r1 are the tomograms sectioned at the XY plane (yellow dashed lines) in panels m-r. Nuclei were stained with DAPI. Scale bar, 50 μm. C A detailed description of the endodermal differentiation process within the hiPSC colony through collective cell migration. Differences in spatial self-organization within the hiPSC colonies in cultures with and without the ring-barrier removal are shown. All experiments were repeated independently at least three times with similar results
Subsequently, the results of fluorescent staining of paxillin and F-actin showed that actin stress fibers were abundant along basal sides of cells at the colony edge in cultures without the ring-barrier (Fig. 7B). Many of the spots of paxillin, a focal adhesion protein, were distributed at the leading edge with lamellipodia and strong paxillin spots at the basal side. A comparison of the central and peripheral regions of the colony revealed an increase in actin filaments and paxillin spots in cells located at the colony edge. In contrast, in cultures with a ring-barrier, the cells showed reduced actin filaments on the basal side and weak paxillin staining with small, thin adhesions throughout the colony. Collectively, these findings demonstrate that collective cell migration and EMT at the colony edge in cultures with the ring-barrier are required to promote the induction of hiPSC-derived endoderm differentiation (Fig. 7C).
留言 (0)