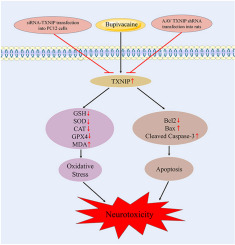
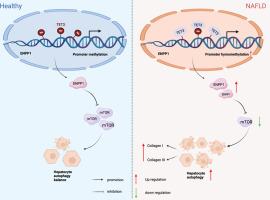
Over the past decades, various forms of regulated cell death (RCD) have been explored to develop anticancer drugs [1,2]. Apoptosis is a well-established RCD, and target apoptotic pathways have already been a practical strategy for anticancer drug research and development [3]. Recently identified non-apoptotic RCDs, including necroptosis, ferroptosis, and pyroptosis, also demonstrate great potential in cancer therapy [1,2]. Mitochondrial permeability transition (MPT)-driven necrosis is a form of non-apoptotic RCD triggered by specific intracellular perturbations, especially oxidative stress and cytosolic Ca2+ overload, and highly relies on cyclophilin D (CypD) [4]. MPT was first proposed in the 1970s, when Ca2+-mediated membrane transition in the mitochondria was discovered [[5], [6], [7], [8]]. MPT refers to increased permeabilization of the inner mitochondrial membrane (IMM), leading to an abrupt loss of mitochondrial membrane potential (MMP), ATP depletion, and osmotic breakdown of mitochondrial membranes [4,9]. MPT is mediated by a supramolecular entity with pore properties called mitochondrial permeability transition pore (MPTP), which is assembled at the junctions between the IMM and outer mitochondrial membrane (OMM) [4]. A series of proteins, such as CypD, voltage-dependent anion channel, adenine nucleotide translocator, translocator protein, hexokinase II, mitochondrial creatine kinase, the Bcl-2 family proteins, and F1/Fo ATP synthase, were proposed to be the key components of MPTP. However, none of them were confirmed in genetic deletion models [[9], [10], [11]]. CypD is the only protein whose essential role in MPT has been formally validated [4,12,13]. CypD inhibitors, including cyclosporin A (CsA), sanglifehrin A, and JW47, effectively prevent MPT-driven necrosis [12,14,15]. MPT-driven necrosis is involved in a variety of pathological processes, including cardiac diseases, degenerative diseases, and cancer [9]. CypD-dependent necrosis contributes to the cytotoxicity of cisplatin and taxol in cancer cells [16,17]. Natural compounds, such as curcumin, honokiol, and icaritin, were reported to trigger MPT-driven necrosis in cancer cell lines [[18], [19], [20]].
Isobavachalcone (IBC), a natural chalcone, has anticancer, antimicrobial, anti-inflammatory, antioxidative, and neuroprotective activities [21]. It was identified as a natural SARS-CoV-2 entry inhibitor and might have therapeutic potential for SARS-CoV-2 virus infections [22,23]. The anticancer effect of IBC was reported in leukemia [24], lung and breast [25], liver [26], prostate [27], colorectal [28], and gastric [29] cancers in vitro. IBC potentially inhibits Akt by reducing Akt phosphorylation in lung cancer cells [25]. In acute myeloid leukemia cells, IBC directly targets dihydroorotate dehydrogenase (DHODH) to induce apoptosis and differentiation [24]. Other studies showed that thioredoxin reductase 1 [27] and ERKs/RSK2 [26] are involved in the anticancer effect of IBC.
Our previous study showed that IBC can remarkably improve experimental colitis in mice by targeting NF-κB p65 [30]. In the present study, we demonstrated that IBC has anticancer potentials by triggering reactive oxygen species (ROS)-mediated non-canonical MPT-driven necrosis, which is distinct from the previously identified apoptotic characteristics induced by IBC. This type of necrosis might have the potential to fight cancers.

留言 (0)