
記住我
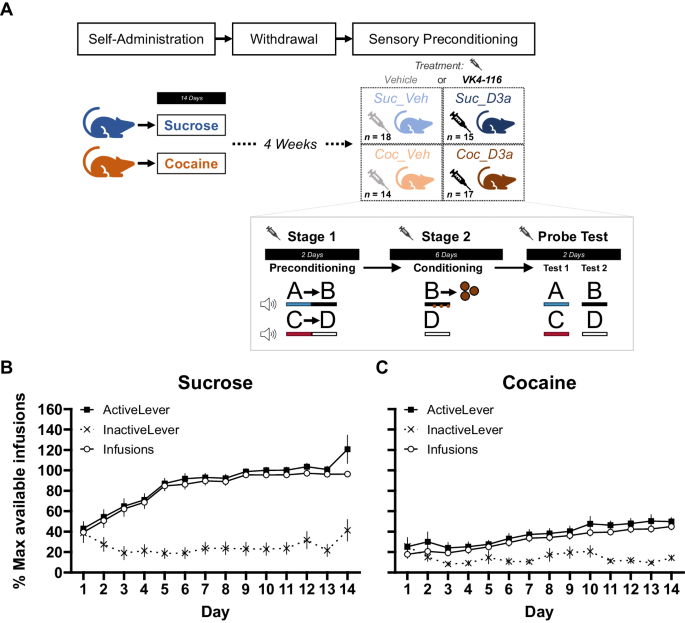
In our previous study, we discovered changes in the expression of myelin-related genes in response to CSDS [9]. These changes were accompanied by differences in myelin thickness, which occurred in a brain region- and strain-dependent manner, altogether demonstrating myelin plasticity as an integral part of the stress response [9]. These findings prompted us to further investigate whether chronic stress impacts other myelin features, such as nodes of Ranvier. To investigate differential expression of genes involved in the structure and function of nodes of Ranvier (Fig. 1a and Table S1), we re-analyzed our previously published RNA-sequencing dataset from the mPFC (including the prelimbic, infralimbic, and anterior cingulate cortex regions) of B6 and D2 mice, performed 1 week after cessation of a 10-day CSDS [9]. We examined 40 genes associated with the Gene Ontology term node of Ranvier. Twelve genes (30%) were differentially expressed in B6 susceptible mice compared to controls, with 11 of them (28%) being expressed at a lower level in the susceptible compared to control mice (Fig. 1b). These genes encoded for voltage-dependent ion channels (e.g., Kcnq2; Kv7.2 and Scn1a; Nav1.1) and structural components of the nodes (e.g., Gjc; Connexin-47; Mag; Myelin associated glycoprotein). In addition, to examine whether genes encoding for structural subcomponents of the nodes of Ranvier (Fig. 1a) were over-represented among the up- or downregulated genes, we conducted Gene Set Enrichment Analysis (GSEA) [31] on differential gene expression lists of B6 resilient, B6 susceptible, and D2 susceptible mice compared to controls (Fig. 1c, d). As D2 resilient mice are found very infrequently, they were not available in the cohort used for this experiment. We found that paranodal, juxtaparanodal, and nodal genes were significantly overrepresented among the downregulated genes in the D2 susceptible mice compared to same-strain controls. In B6 susceptible mice, we observed a statistical trend for overrepresentation of node of Ranvier genes among the downregulated genes. GSEA performed with all 40 genes associated with nodes of Ranvier produced similar results (Fig. 1c, d). Taken together, these results suggest that chronic psychosocial stress alters the expression levels of genes involved in node of Ranvier structure and function in the mPFC of B6 and D2 susceptible mice.
Fig. 1: Differential expression of nodes of Ranvier genes after chronic social defeat stress.
a Schematic representation of the node of Ranvier subregions. b Heatmap showing the expression fold change (logFC) and significance of differential expression for genes associated with nodes of Ranvier subcomponents (node, light blue; paranode, pink; juxtaparanode, orange; internode, green) in B6 and D2 defeated mice in the mPFC. c Gene Set Enrichment Analysis (GSEA) of genes associated with the individual structural subcomponents of the nodes of Ranvier (node, paranode, juxtaparanode, internode) and combined analysis of all nodal genes (N = 40 genes, see Supplementary Table 1). d–f GSEA enrichment score figures of all nodal genes. The top portion of the plot shows the enrichment score, which reflects the degree of overrepresentation of the gene set at the top or bottom of a ranked gene list. The middle portion of the plot shows the position of the genes in the ranked list. The bottom portion of the plot shows the value of the ranking metric as the analysis walks down the list of ranked genes. B6: C57BL/6NCrl; D2: DBA/2NCrl; Con control, Res resilient, Sus susceptible. *p < 0.05, **p < 0.01, ***p < 0.001.
Strain-dependent alterations of mPFC nodal morphology after chronic psychosocial stressTo investigate whether the observed gene expression differences are associated with changes in node of Ranvier morphology, we developed a novel 3D reconstruction analysis of nodal subdomains. Particularly in gray matter nodes of Ranvier can present highly variable orientations, and as a result, two-dimensional maximum projection analysis of morphology can induce significant distortions of measured parameters. By measuring nodes as they exist in three-dimensional space, we can obtain accurate morphological measures. We analyzed nodal subdomains after CSDS in the anterior cingulate cortex subregion of the mPFC, a critical hub for the regulation of anxiety [33]. This brain region contains several short- and long-range connections [34] as well as excitatory and inhibitory neurons, both of which are known to be myelinated [35]. To investigate whether genetic background or susceptibility or resilience to stress influence nodal modifications in response to stress, we conducted the morphological analysis both in B6 and D2 mice, classified either as stress-resilient or -susceptible based on social avoidance behavior following CSDS [9, 27] (Fig. S1).
We found that B6 resilient mice had shorter paranodes (by 10.3%), identified by contactin-associated protein (CASPR) immunoreactivity (Fig. 2a, b) compared to same strain controls (Fig. 2c). In D2 mice we found a statistical trend toward increased paranode length in susceptible mice compared both to same strain resilient (by 27.0%) and control mice (by 11.1%) (Fig. 2f). Moreover, node width, defined as the distance between two flanking paranodes (Fig. 2a, b), was shorter (by 5.2%) in D2 susceptible mice compared to same strain control mice (Fig. 2g), but not in B6 mice (Fig. 2d). In accordance with shorter paranodes, total nodal region length was shorter in resilient B6 mice (by 8.4%) compared to same strain controls (Fig. 2e, h). These changes in paranode length are similar in magnitude with previous findings in multiple sclerosis, in which paranodal changes associate with predicted changes in conduction velocity [36].
Fig. 2: Strain-dependent changes in nodal morphology after chronic social defeat stress.
a, b Examples of 3D reconstructed paranodes in the anterior cingulate cortex layer V/VI in B6 and D2 mice after chronic social defeat stress. Quantification of paranode length (c, f), node width (d, g), total nodal region length (e, h). c Paranode length B6: Con = 266 paranodes from 5 mice, Res = 311 paranodes from 6 mice, Sus = 286 paranodes from 7 mice. Statistical differences were identified by linear mixed-effect modeling with pairwise comparisons: Con vs. Res, t(13.5) = −2.356, p = 0.034; Con vs. Sus, t(13.8) = −1.488, p = 0.159; Res vs. Sus, t(9.4) = 1.148, p = 0.279. d Node length B6: Con = 132 nodes from 5 mice, Res = 152 nodes from 6 mice, Sus = 168 nodes from 7 mice. Con vs. Res, t(9.4) = −0.141, p = 0.891; Con vs. Sus, t(9.5) = −0.972, p = 0.355. e Nodal region length B6: Con = 124 nodes from 5 mice, Res = 136 nodes from 6 mice, Sus = 152 nodes from 7 mice. Con vs. Res, t(13.6) = −2.630, p = 0.020; Con vs. Sus, t(13.7) = −1.628, p = 0.126. f Paranode length D2: Con = 334 paranodes from 7 mice, Res = 274 paranodes from 5 mice, Sus = 476 paranodes from 9 mice, Con vs. Res, t(16.6) = 0.040, p = 0.968; Con vs. Sus, t(16.8) = 1.889, p = 0.076; Res vs. Sus, t(10.5) = 1.885, p = 0.087. g Node length D2: Con = 151 nodes from 7 mice, Res = 131 nodes from 5 mice, Sus = 215 nodes from 9 mice. Con vs. Res, t(17.5) = −0.487, p = 0.632; Con vs. Sus, t(18.1) = −2.402, p = 0.027. h Nodal region length D2: Con = 154 nodes from 7 mice, Res = 124 nodes from 5 mice, Sus = 219 nodes from 9 mice. Con vs. Res, t(15.7) = 0.043, p = 0.966; Con vs. Sus, t(15.6) = 1.035, p = 0.317). B6: C57BL/6NCrl; D2: DBA/2NCrl; Con control, Res resilient, Sus Susceptible.
Strain-dependent alterations of nodal morphology in forceps minorIn addition to cortical gray matter, we analyzed nodal morphology in response to CSDS in the forceps minor (Fig. S2a), a white matter tract adjacent to the mPFC. Paranode length was longer in B6 stress-resilient (by 7.0%) compared to same strain controls, and there was a statistical trend toward longer paranodes (by 4.4%) in stress-susceptible mice (Fig. S2b). Paranode length in D2 mice or node width in either B6 or D2 mice did not change in response to CSDS (Fig. S2c–e). Altogether, our analysis demonstrates that chronic stress alters node of Ranvier morphology both in the mPFC gray matter and in the forceps minor white matter. These effects were strongly influenced by genetic background and brain region, and they depended on the individual stress response.
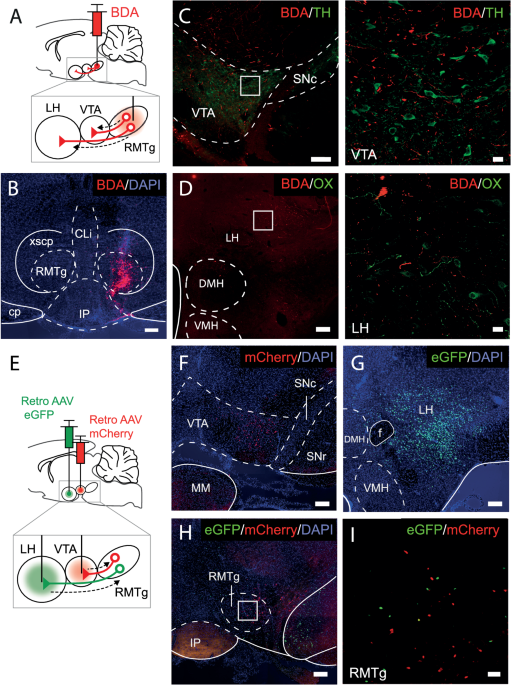
Repeated activation of the vHPC-mPFC pathway reduces anxiety-like behavior and paranode length in axon-specific mannerMechanisms underlying experience-dependent changes in node of Ranvier structure are largely unknown. We hypothesized that neuronal activity may be one such mechanism because it influences oligodendrocyte precursor cell proliferation, oligodendrogenesis, and myelin thickness [37, 38]. Furthermore, transcranial magnetic stimulation affects node of Ranvier width in the stimulated area [23]. However, whether activity-dependent nodal remodeling occurs in circuits involved in the regulation of anxiety-like behavior, and whether such effects are specific to activated axons, are not known. To study the effects of neuronal activation on nodal morphology, we chemogenetically activated neurons projecting from the vHPC to the mPFC, a pathway previously shown to increase anxiety-like behavior upon acute stimulation [39, 40]. To achieve projection-specific expression of excitatory hM3Dq-receptors, we bilaterally injected a retrograde Cre-carrying virus (AAVretro-Cre-eGFP) into the mPFC, while a Cre-dependent hM3Dq (AAV8-Dio-hM3Dq-mCherry) or an mCherry (AAV8-Dio-mCherry) control vector was injected into the vHPC (Fig. 3a) of male C57BL/6NCrl mice. Immunohistochemical analysis confirmed the accuracy of the labeling by showing robust eGFP labeling in the mPFC and mCherry expression in the vHPC CA3/1 regions, as well as eGFP/mCherry-double positive axons along the pathway (Fig. 3b).
Fig. 3: DREADD-mediated activation of vHPC-to-mPFC projection neurons reduces anxiety-like behavior.
a Projection-specific expression of hM3Dq/mCherry was achieved by injecting a retrograde Cre-carrying virus (retro-Cre-eGFP) into the mPFC and a Cre-dependent hM3Dq (Dio-hM3Dq-mCherry) or a control virus (Dio-mCherry) into the vHPC CA3 subregion. b Representative examples showing eGFP expression in the mPFC and mCherry-expressing axons in the hippocampal fimbria. c Experimental timeline. Open zone (d), closed zone (e) risk assessment (RA) zone (f) time and the number of stretch-attend postures in the RA zone (g) in the acute elevated zero maze test (EZM1) (two-sided Student’s t tests). Time spent in the center of the open field (OF) test (h) and distance traveled (i) during the OF test (two-sided Student’s t tests). j Quantification of social interaction ratio in the social approach (SA) task (two-sided Student’s t test). k Time spent in the interaction zone during empty and target sessions of the SA test (two-way repeated ANOVA: group × session F(1,22) = 0.703, p = 0.402; session F(1,22) = 6.111, p = 0.022; group F(1,22) = 0.100, p = 0.755). Time spent in the open zone (l), closed zone (m) (two-sided Student’s t tests), risk assessment zone (n) (two-sided Welch’s t-test) and the number of stretch-attend postures in the RA zone (o) in the EZM2 test after repeated activation of vHPC-to-mPFC pathway. Time spent in the closed arms (p), center zone (q), open arms (r) and the number of stretch-attend postures (s) in the elevated plus maze (EPM) test (two-sided Student’s t tests). Control n = 11; hM3Dq n = 13; Error bars represent ±SEM.
Behavioral tests confirmed that the activation of this specific pathway was effective in modulating anxiety-like behavior (Fig. 3c), although the effects differed from those previously published for acute stimulation [39, 40]. We first assessed the effects of an acute chemogenetic activation of Gq signaling on anxiety-like behavior by testing the mice in the elevated zero maze (EZM) 20–30 min after a single clozapine-N-oxide (CNO) injection, which activates the hM3Dq receptors. In the EZM, the hM3Dq and control groups spent equal amount of time in the open areas, closed areas, and in the risk assessment zone, and we did not observe differences in stretch-attend postures or general activity (Figs. 3d–g and S3a), suggesting ineffectiveness of the acute manipulation to alter anxiety-like behavior in this assay. After the acute test, we continued daily CNO injections to assess the effects of a repeated manipulation of vHPC-to-mPFC activity on anxiety-like behavior. After 12 days of CNO injections, we carried out the open field (OF) test. Anxiety-like behavior or general activity did not differ between the groups, as demonstrated by similar time spent in the center zone and the distance traveled (Fig. 3h, i). On the following day, and after another CNO injection, we tested the mice in the EZM task. In contrast to the acute CNO effects, after the repeated manipulation hM3Dq mice spent more time in the risk-assessment zone, compared to controls (Fig. 3n). This was accompanied with reduced closed area time (Fig. 3m), with no differences in the time spent in the open areas (Fig. 3l) or general activity (Fig. S3b). The behavior of the control mice did not differ between the acute and chronic EZM tests (Fig. S3c–f), supporting the feasibility of repeated testing in the EZM.
On the following day (after 14 days of CNO injections), we tested the effects of the repeated activation of the vHPC-to-mPFC projection on social behavior in the social approach (SA) task. hM3Dq and control mice spent equal amounts of time interacting with a social target (Fig. 3j, k), suggesting no effects on social avoidance behavior.
Lastly, we tested the priming effects of an acute hM3Dq activation on anxiety-like behavior. After receiving daily CNO injections for 15 days, the mice received a single CNO injection 30 min prior to testing in the elevated plus maze (EPM), to avoid a habituation effect from repeated testing in the EZM. An acute activation in primed animals resulted in reduced exploration of the closed arms in hM3Dq mice, which was accompanied with increased center zone time and increased number of stretch-attend postures compared to controls, with no change in open arm time or in locomotor activity (Figs. 3p–s and S3e). Taken together, our behavioral data demonstrate that prolonged manipulation of vHPC-to-mPFC activity decreases anxiety-like behavior, reflected particularly as increased risk assessment in the EZM test, together with reduced closed arm exploration and increased engagement in stretch-attend postures in the EPM test.
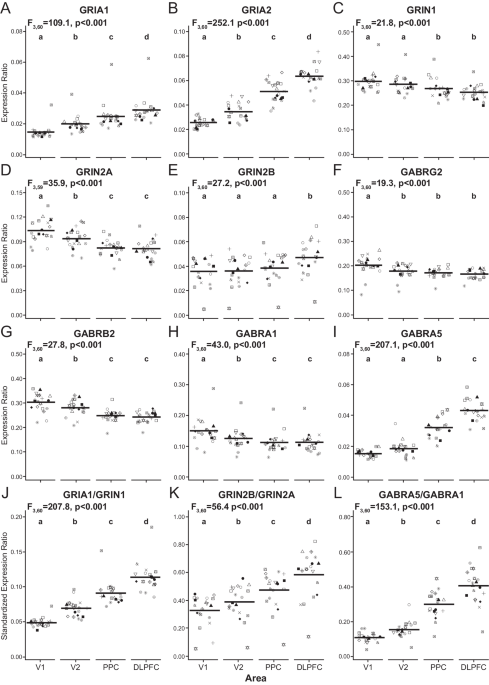
Finally, we assessed whether DREADD-mediated activation, changing anxiety-like behavior, altered nodal morphology along labeled vHPC-to-mPFC axons. For this, we imaged paranodes (identified with CASPR) on mCherry-expressing (mCherry+) axons within the hippocampal fimbria, where robust labeling of mPFC-projecting vHPC axons was observed in both hM3Dq and control mice (Fig. 4a, b). To tease out manipulation-specific effects, we also studied paranodes on non-labeled axons (mCherry−) within the same brain region. In mice with vHPC-to-mPFC projection neurons expressing hM3Dq, paranodes were significantly shorter (13.9%) than in controls (Fig. 4c). Importantly, the effect was specific for paranodes on mCherry-labeled axons, as no difference in paranode length was present between the groups in non-labeled axons. Furthermore, paranode length did not differ between labeled and non-labeled axons in control mice. In concordance with reduced paranode length, the total nodal region was shorter in hM3Dq-expressing neurons compared to control neurons (Fig. 4e). The DREADD-mediated activation did not affect node width (Fig. 4d). These results demonstrate that manipulation of neuronal activity specifically affected paranode length, and, importantly, the modification occurred in the activated, but not in the non-activated, axons, supporting our hypothesis on activity-dependent remodeling of nodes.
Fig. 4: Repeated activation of vHPC-to-mPFC projection neurons reduces paranodal length.
a A sagittal section containing the vHPC immunolabeled with CASPR antibody in mice expressing an mCherry-labeled hM3Dq or a control virus in vHPC-to-mPFC projection neurons. Outlines demonstrating paranodes along an mCherry-labeled axon. b 3D reconstruction of paranodes on labeled and non-labeled axons. c Quantification of paranode length (c Linear mixed model: Group_virus t(16.8) = −0.187, p = 0.854; Axon_type t(210) = 0.954, p = 0.341; Group_virus × Axon_type t(205) = −2.463, p = 0.015; pairwise comparisons: hM3Dq-mCherry+ vs. Control-mCherry+t(11) = −2.732, p = 0.019; hM3Dq-mCherry− vs. Control-mCherry−t(9.7) = −0.047, p = 0.963). d Quantification of node length (d Group_virus t(18.6) = −0.060, p = 0.953; Axon_type t(193) = 0.507, p = 0.613; Group_virus × Axon_type t(191) = 0.158, p = 0.874). e Quantification of total nodal region length (e Group_virus t(17.8) = 0.244, p = 0.810; Axon_type t(193) = 1.622, p = 0.106; Group_virus × Axon_type t(191) = −2.569, p = 0.011; pairwise comparisons: hM3Dq-mCherry+ vs. Control-mCherry+t(9.8) = −2.425, p = 0.036; hM3Dq-mCherry− vs. Control-mCherry−t(9.2) = 0.308, p = 0.765). c hM3Dq: 73 mCherry− and 123 mCherry+ paranodes from 6 animals; Control: 80 mCherry− and 134 mCherry+ paranodes from 6 animals. d hM3Dq: 37 mCherry− and 62 mCherry+ nodes from 6 animals; Control: 39 mCherry− and 61 mCherry+ nodes from 6 animals. e hM3Dq: 37 mCherry− and 62 mCherry+ total nodal regions from 6 animals; Control: 37 mCherry− and 64 mCherry+ total nodal regions from 6 animals. b Size bar 2 µm. Error bars represent ±SEM.
留言 (0)