

記住我






Circadian rhythms influence almost all aspects of biology, from complex organismal physiologies and behaviors to cellular processes and synchronized patterns of molecular gene expression in health and disease (Chaix et al., 2016; Lee et al., 2021). The elucidation of the genes, logic, and feedback loops that constitute most eukaryotic molecular clocks has not only extended our knowledge to the mechanistic, molecular level, but reemphasized how pervasive and conserved the functions of the clock are in biology. One outstanding problem in the field is understanding the relationship between the central clock and the different “peripheral clocks” that regulate specific physiological processes (Hardin et al., 2003; Ito and Tomioka, 2016; Sehgal, 2016; Di Cara and King-Jones, 2016; Selcho et al., 2017; Yildirim et al., 2022).
Until recently, data directly linking circadian rhythms with memory formation has been somewhat limited (Eckel-Mahan et al., 2008; Gerstner et al., 2009; Gerstner and Yin, 2010; Phan et al., 2011; Xia and Storm, 2017; Price and Obrietan, 2018; Rawashdeh et al., 2018; Snider et al., 2018), or seemingly contradictory (Price et al., 2016; Wang et al., 2021). Like other circadian-regulated processes, initial experiments showed that the “time-of-day” (TOD) of behavioral training had effects on behavioral performance, with a certain time across the 24-h being optimal (Ralph et al., 2013). Animals perform best when tested at 24-h intervals from the training time, a property termed “timestamping” (Ralph et al., 2002; Cain et al., 2004a,b, Cain et al., 2017). Long-term potentiation, a popular cellular model for memory formation, exhibits TOD properties, potentially linking timing preference to a mechanism involved in memory formation (Barnes et al., 1977; Chaudhury et al., 2005; Nakatsuka and Natsume, 2014). Importantly, core molecules involved in memory formation (in different subcellular locations including at synapses) oscillate in their amounts or activities, and these are indirectly under circadian control (Eckel-Mahan et al., 2008; Phan et al., 2011; Bruning et al., 2019; Noya et al., 2019; Smies et al., 2022). Similar behavioral results in simpler organisms, including Drosophila, further reinforced the likely universality of circadian involvement in memory processes (Fernandez et al., 2003; Lyons et al., 2005; Lyons and Roman, 2008; Fropf et al., 2014; Lubinski and Page, 2016; Fropf et al., 2018). However, the actual role(s) of circadian biology in learning and memory formation have not been mechanistically defined in detail.
Drosophila has been used to investigate many problems in neurobiology, including the ground-breaking work that led to the elucidation of the molecular clock (Hall, 2017; Young, 2018; Rosbash, 2021). Two different behavioral assays have been utilized to study learning and memory formation. The olfactory avoidance behavior uses classical conditioning to test the ability of flies to associate and remember one of two odors that is presented with an electric foot shock (Tully and Quinn, 1985; Tully et al., 1994). The courtship suppression assay tests the ability of male flies to learn and remember the non-responsive mating behavior of previously mated females (Siegel and Hall, 1979). Subsequent suppression of the normally aggressive male mating behavior is used to assay learning and memory formation. The two assays differ in their ease to setup, the ability to control the salient behavioral cues, and the detailed anatomical, kinetic, and molecular information that is required during memory formation.
Sakai et al. (2004) initiated experiments showing circadian influences on courtship suppression behavior. All the circadian mutants that were tested, with the exception of period, did not affect the memory of courtship suppression. However, in their ground-breaking work, Inami et al. discovered that post-training incubation of flies in constant darkness (DD) but not constant light (LL), disrupted memory formation (Inami et al., 2020). They went on to define a window of time when DD incubation inhibited memory, and showed that activity of the central clock neurons, in particular the ones that secrete the peptide PDF, are required. Secretion of PDF peptides from the l-LNvs is sufficient to rescue the requirement for light, suggesting that light reception “upstream” or in these neurons results in PDF secretion. Finally, they showed that the PDFR is required and that dCREB2 activity downstream of PDF/PDFR is involved in memory formation. In a follow-up article, Inami et al. inactivated the central clock neurons prior to training and showed that this inhibited this process (Inami et al., 2021).
Lyons and Roman (2008) pioneered the demonstration that circadian biology affects fly learning in the olfactory avoidance assay. Learning is traditionally defined as immediate performance after flies are exposed to a single training trial. They showed that the TOD of training affects performance, with a peak in performance during the early nighttime period. Fropf et al. (2014) showed that the TOD of training also affects long-term memory, memory that results from 10 cycles of spaced training which is typically measured a day or more after the end of training. 1-day (1d) memory showed a TOD effect, and the core circadian gene, period, is somehow involved in this process (Fropf et al., 2018). A TOD effect that requires an intact central clock has also been demonstrated for the memory of a fly appetitive behavioral paradigm (Chouhan et al., 2015, 2017).
Several outstanding issues remain after the publication of these important articles. There is confusion about whether the circadian genes themselves are involved, and if so, why mutations in multiple core clock genes (tim01, clkjrk, cyco) do not affect memory formation in courtship suppression (Sakai et al., 2004). These mutations result in “loss-of-function” phenotypes in locomotor activity, and the simple expectation is that they would disrupt memory formation if circadian genes are needed. In contrast, the per mutants have strong effects on performance in courtship suppression and overexpression can even enhance the process (Sakai et al., 2004). Using the olfactory avoidance behavior, Chen et al. (2012) previously showed that the dorsal anterior lateral neurons (DAL) are important for memory formation and that some of the circadian genes are expressed in those cells in response to training (Lin et al., 2021). However, just like with courtship suppression, 1d memory was unaffected in timo, clkjrk, or cyco mutants, while per mutants have a strong effect on 1d memory (Chen et al., 2012).
In this report, we use multiple approaches to ask about circadian clock involvement in memory formation. We use the term memory formation to describe the overall process of encoding, consolidation (both cellular and systems reorganization), memory maintenance, and retrieval (Squire and Alvarez, 1995). Until more behavioral, cellular, and molecular data is available, the distinctions between these processes are somewhat arbitrary. We think it is simplest to posit that maintenance begins after all anatomical reorganization (systems consolidation) is complete (Cervantes-Sandoval et al., 2013; Dubnau and Chiang, 2013). We present evidence for circadian dependency during a temporal window 1–3 days after spaced training. This window is significantly later than the 1-day timepoint which has historically been viewed as “long-term memory.” Our window is most likely only one of many transcriptional windows required during memory formation (Hirano et al., 2016; Chen et al., 2020; Mizuno et al., 2020; Inami et al., 2021). Though our results do not resolve all the questions in this field, they provide important kinetic information, genetic tools, and an experimental strategy to address these uncertainties in the future.
Materials and methods FliesThe w1118 (used here as a “wild type”) and the HS-vri stocks have been described and validated previously (Glossop et al., 2003; Gunawardhana and Hardin, 2017; Gunawardhana et al., 2020). The HS-vri transgene was backcrossed multiple times into the w1118 background so that they are isogenic. The HS-Clkjrk transgenic fly was made from the previously described CLKjrk mutant fly (Allada et al., 1998; Darlington et al., 2000). A plasmid containing the fly CLKjrk mutant open reading frame was obtained and an EcoRI fragment was subcloned into the pCaSpeR-HS plasmid, and the resulting plasmid was injected into a w1118 background stock to make transgenic flies (Thummel, 1996). That w1118 background was used as the control in Figure 2, Expt #7. The iso31 (used here as “wild type”) and Pdp13135 mutant stocks have been described and validated previously (Zheng et al., 2009). The Pdp13135 mutation resides in the iso31 background.
Olfactory avoidance behaviorYoung flies (less than 1 week of age) were used for behavior. Flies were collected and kept in groups of 100–150 individuals/vial and entrained to a 12-h lights on:12-h lights off schedule at 20°C for 3d prior to training. Training started between ZT = 14 and ZT = 16, and flies were returned to their dark period and kept at 20°C until testing. Testing always occurred around ZT = 16 to ZT = 18. For experiments involving changes in post-training lighting conditions, flies were incubated after training under standard LD conditions until constant conditions started, at which time the appropriate group was shifted to darkness or light (at 20°C). For the HS-vri flies that underwent post-training heat-shock, flies were placed in food-containing vials and incubated under light:dark control at 28°C for almost two full-days. For pre-training heat-shock, HS-vri flies were induced at 28°C for 3 h of time. For the HS-Clkjrk flies that underwent post-training induction, flies were transferred to empty food vials, and incubated at 36°C in a water bath for 30’ duration at 32 h and 56 h after the end of training, then returned to regular vials and incubated at 20°C under light:dark control until testing. For HS-Clkjrk flies that underwent pre-training induction, heat-shock (a 20°C to 36°C temperature shift for 30’) was done in a water bath, and flies were allowed to recover for 90’ prior to training.
Flies were trained in the olfactory avoidance-training paradigm developed by Tully and Quinn and modified to allow for automated training sessions (Tully and Quinn, 1985; Tully et al., 1994). A single-cycle of training consists of 90 s exposure to ambient air; 60 s of electric shock [the unconditioned stimulus which consists of 70 V pulses lasting 1.5 s and administered every 5 s (12 total)] accompanied by simultaneous exposure to 1 odor (the conditioned stimulus, CS+); 45 s of ambient air exposure to clear the first odor; 60 s of exposure to the second odor with no shock (the CS- condition), 45 s of ambient air to clear the second odor. This single training trial takes about 2.6 min. Spaced training consists of 10 single cycles separated by 15-min rest intervals. This training requires about 2 h 40 min of time. Massed training consists of 10 consecutive single cycles of training, and takes 37 min. Testing was done by placing flies in a choice point and giving them 2 min to decide between the CS+ and CS- stimuli. We used 3-octanol and 4-methylcyclohexanol as the odors. The performance index = [the number of flies making the correct choice] − [the number of flies making the incorrect choice]/total number of flies, multiplied by 100. To avoid odor-avoidance biases, we calculate the performance index of every single N by taking an average performance of two groups of flies, one trained with 3-octanol as CS+, and the other with 4-methylcyclohexanol. Flies were trained in a balanced manner, such that the sequence of shock-paired odors alternates, as well as the assignment of left vs. right arm at the choice point during testing. Data is presented as the standard error of the mean, and the Student’s T-test was used to evaluate statistical significance in pairwise comparisons.
SleepMale flies within 3 days of eclosion were collected and entrained to a 12:12 light/dark cycle for 2 days before assaying for sleep. Individual flies were loaded into tubes under CO2 anesthesia and placed in the Drosophila Activity Monitor (DAM) System and put into an incubator maintained at 20°C under 12 h light:12 h dark cycles (Pfeiffenberger et al., 2010a). Raw activity counts were collected over a 24-h period. An Excel program was developed and used to calculate the various sleep measures and to plot the sleep profiles (Pfeiffenberger et al., 2010b; Cui personal communication). The sleep curves represent the average amount of sleep in each hour and the error bars represent the standard error of the mean. Dead flies (those with two or more consecutive days without locomotor activity) were manually eliminated. Sleep was measured from two groups of flies, one group was maintained in a 20°C incubator for the entire duration of the experiment. The second group was placed in a different incubator kept at 20°C until the time of heat-shock, when the temperature of the incubator was shifted to 28°C.
Circadian locomotor activityMale flies within 3 days of eclosion were collected and entrained to a 12:12 light/dark cycle at 20°C for 2 days prior to assaying for circadian activity. Individual flies were anesthetized using CO2, loaded into tubes, and placed in the DAM system to measure locomotor activity under 12:12 light/dark cycles at 20°C for 3 days (Pfeiffenberger et al., 2010a). The incubator containing the DAM monitors was shifted to constant darkness for 2 days, then shifted to 28°C for the remaining 4 days while maintained under constant darkness. Locomotor activity was monitored for the entire duration of the experiment, and data were analyzed using an Excel program we developed (Pfeiffenberger et al., 2010a; Cui personal communication). Table 2 compiles the average total number of beam crossings per minute (binned across hourly units) for the different flies and treatments during the entire duration of the recording period.
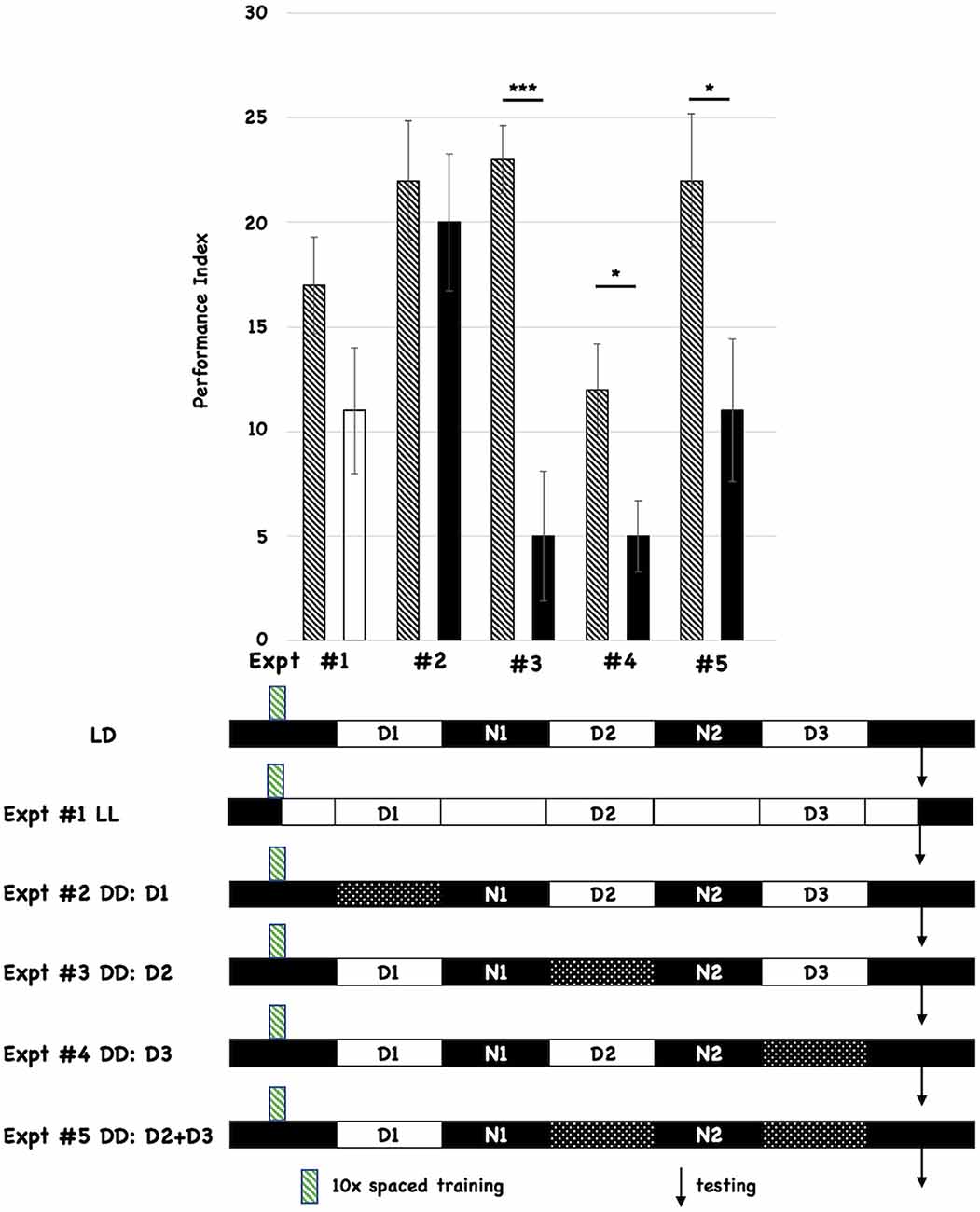
Results Post-training dark periods (D) inhibit memory formationAll flies were initially entrained to 12-h light:12-h dark cycles for 3 days prior to behavioral training (10 cycles of spaced training, indicated with the green hatched box below the histograms in Figure 1). We tested the effects of post-training changes in light:dark incubation by comparing the performance of the experimental group relative to control flies that were incubated in standard light:dark conditions after training (shown as the first cartoon at the bottom of Figure 1, and labeled LD). Each different post-training lighting condition was tested individually against the control LD group, and the behavioral data for each experiment (numbered #1-#5) is shown as a separate pair of histograms. The single hatched histogram is the control condition (post-training LD), while the white or black histograms indicate the different incubation conditions being tested. In the cartoon below the Figure, the double white bars indicate constant light, while the double-hatched bars show when D is imposed. All flies were tested for 3-day (3d) memory (indicated with black arrows on the timelines). We initially chose post-training constant light (LL; Expt #1) because it was known that this treatment disrupted the oscillations of the molecular clock (Hardin et al., 1990). However, post-training LL treatment did not significantly affect 3d memory. We then tested the effect of post-training constant darkness (DD). Imposing DD during the first light period after training (D1; Expt #2) did not affect 3d memory. However, imposing constant darkness during D2 (Expt #3), D3 (Expt #4), or both periods (D2+D3; Expt #5) disrupted 3d memory. These results almost exactly parallel those reported by Inami et al. (2020) who used the courtship suppression behavioral assay. We also tested red-eyed flies and found similar effects (see Supplementary Figure 1), suggesting that these effects are not a function of eye color or w1118-mediated retinal degeneration (Ferreiro et al., 2018).
Figure 1. Post-training incubation of flies in dark:dark (DD) conditions disrupts 3d memory. The behavioral data and summary cartoons of the experimental timeline are presented. At the top of the Figure, the Performance Index is plotted as a function of the post-training incubation schedules, and the different pairwise comparisons are numbered (#1–5). Each experiment compares the performance of flies trained and maintained after training under control (LD) conditions (the control group) vs. an experimental group where the post-training lighting conditions change. “Wild-type” w1118 flies are used for both control and experimental groups. The performance scores for the control groups are depicted using a histogram with black-and-white stripes. The performance scores of the different experimental groups are represented using a solid white (LL or constant light after training) or solid black histograms (for those that receive the imposition of one or more D periods). N = 8 for all experiments, T-tests were used to evaluate significance. *p < 0.05, ***p < 0.001. All unmarked pairwise comparisons are not significantly different. At the bottom of the Figure, the first horizontal cartoon (labeled LD) describes the standard, control experimental condition. Flies previously entrained to a 12h:12 h light:dark schedule are behaviorally trained with 10 cycles of spaced training during the early nighttime (green striped box). The different post-training periods are shown as white (12 h light) and black (12 h dark) boxes, and they are labeled (Day1 = D1, Night 1 = N1 etc.). Testing (3d after training) is denoted with the black arrow. Each subsequent horizontal timeline shows a different experiment where post-training lighting conditions are altered, and the Experiment number is indicated and corresponds to its behavioral data shown using histograms. Flies exposed to constant light from the end of training to the time of testing are shown with white boxes only. Flies that experience one or more dark periods (when light normally occurs) are shown using double cross-hatched boxes.
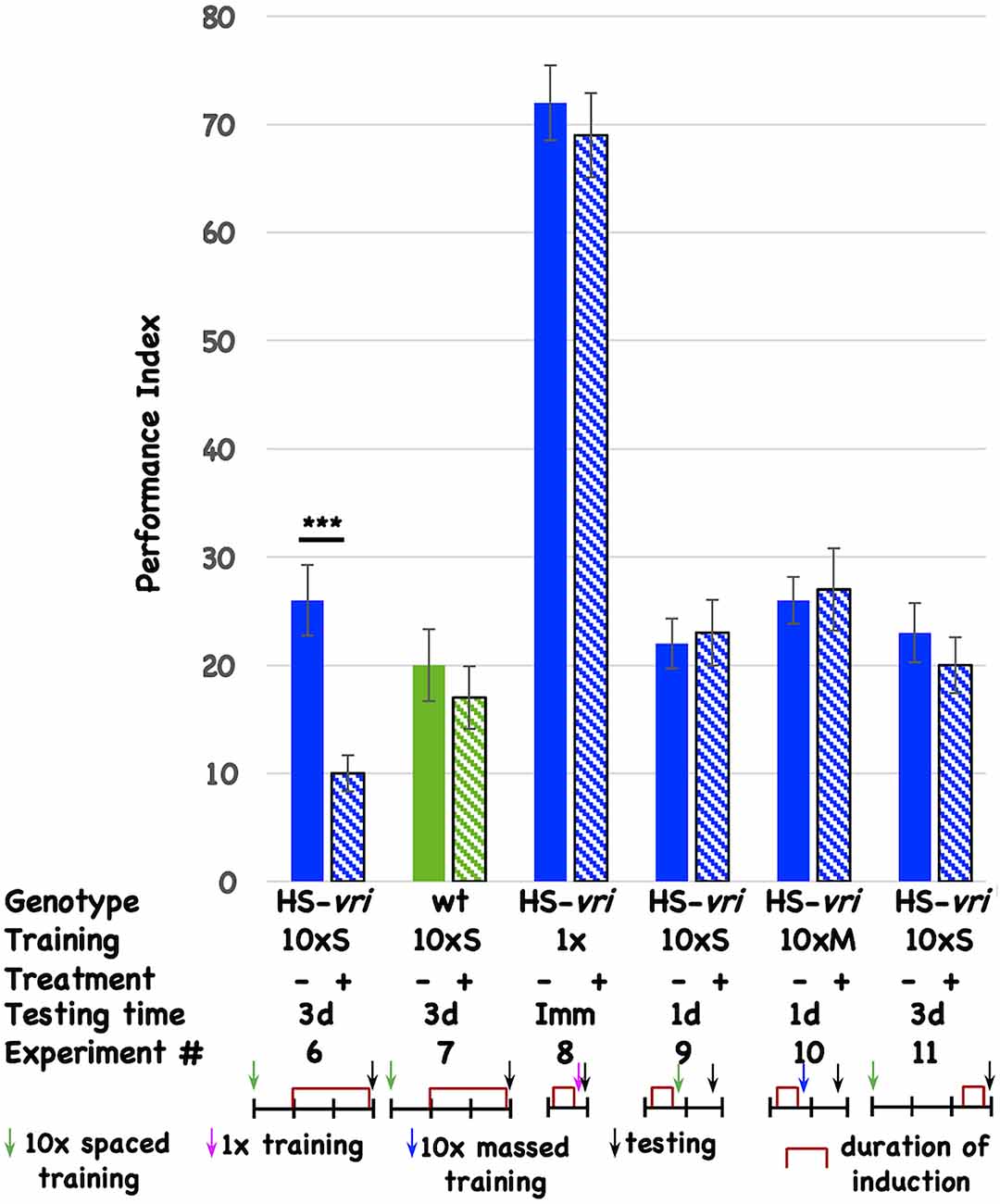
Post-training induction of vrille phenocopies D periods and specifically disrupts long-term memoryTo determine if specific circadian molecules are involved, and to minimize pleiotropic effects that circadian disruption might have on neuronal development, we use inducible transgenes to limit genetic interventions to post-development periods. The ubiquitously expressed, heat-shock-driven vrille transgene (HS-vri) has been used previously to characterize the involvement of this core circadian transcription factor in the central clock (Glossop et al., 2003). Chronic, low-level induction of the transgene beginning 1-day (1d) after the end of 10 cycles of spaced training (10 × S) affects 3d memory (Figure 2, Expt #6), while heat-shock itself does not have an effect in the isogenic wild-type stock that contains the transgene (Figure 2, Expt #7). This effect is specific for certain “phases” of memory since induction prior to 1 cycle of training does not affect immediate performance (conventionally known as learning; Figure 2, Expt #8), nor does pre-training induction affect 1d memory after 10 × S trials (Figure 2, Expt #9) or 10 cycles of massed training [Figure 2, Expt #10 conventionally known as anesthesia-resistant memory (ARM); Tully et al., 1994]. Induction of the transgene prior to behavioral testing (3d after training) likewise does not affect performance, suggesting no effects of transgene induction on retrieval (Figure 2, Expt #11).
Figure 2. Induction of vrille specifically disrupts 3d memory. The performance of flies is plotted as a function of genotype, type of behavioral training, treatment, and testing time. There are two genotypes of flies: HS-vri transgenic (histograms in blue or blue stripes) and otherwise isogenic “wild-type” (w1118, histograms in green or green stripes) flies in which the transgene resides. The performance of flies not induced is shown with solid histograms while that of flies that undergo heat-shock induction is shown in stripes. Underneath the behavioral data, the different experiments are assigned a number, and the details of each Experiment are shown in tabular and cartoon formats. In the table, the top line indicates the genotype of the flies that are used for each experiment. 10 × S indicates flies that receive 10 cycles of spaced training. 1x denotes flies that only undergo a single training trial. 10 × M corresponds to flies that receive 10 cycles of massed training. The — sign denotes no treatment, while the + sign shows data for flies that were induced with a mild, chronic heat-shock (20°C to 28°C shift). The different testing times are denoted as 3d (3-day memory), Imm (immediate performance after a single cycle of training), or 1d (24 h memory). In the cartoon portion of the Figure, the horizontal line denotes the timeline of the experiment, and each vertical tick mark represents 1d of elapsed time. Green, magenta, and blue arrows indicate whether flies received 10 cycles of spaced training, a single cycle of training, or 10 cycles of massed training. The black arrows indicate the time of testing. The red brackets show when the heat-shock induction occurred. The color scheme used for the data histograms (top of Figure) is unrelated to the one used in the cartoons (bottom part of Figure). N = 8 for all experiments, T-tests were used to evaluate significance. ***p < 0.001. All unmarked pairwise comparisons are not significantly different.
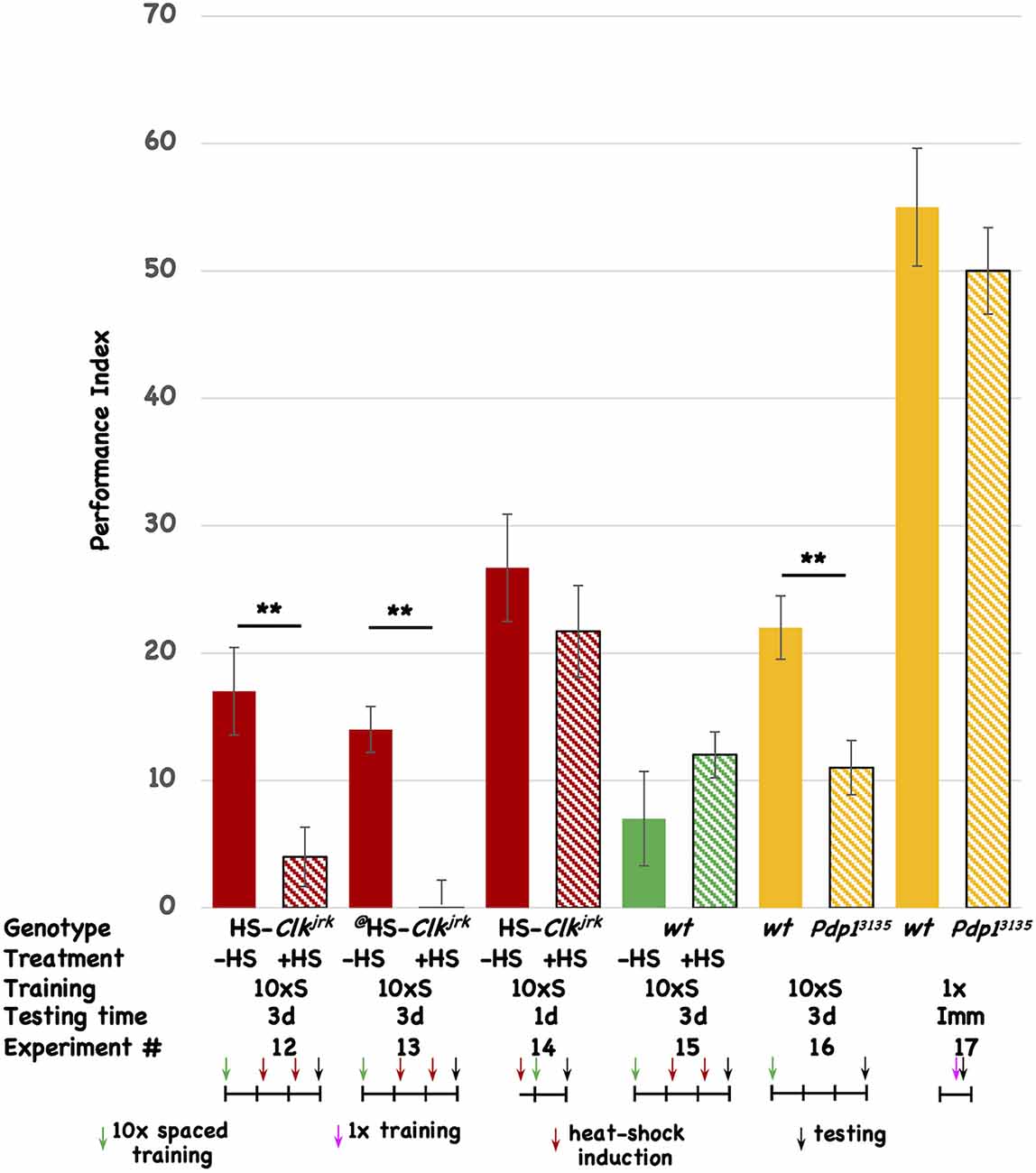
Disruption of other circadian genes affects long-term memoryTo test the involvement of other circadian molecules (and the oscillatory circadian molecular network in general) we tested two other core circadian molecules for an effect on 3d memory. The Clk gene codes for the core transcriptional activator of E-box mediated transcription, dCLK. We made transgenic flies that express the Clkjrk ORF under the control of the heat-shock promoter. The Clkjrk mutant fly contains a truncation in the Clk ORF, resulting in a dominant negative protein, and heat-shock induction of this CLKjrk-encoding transgenic fly should disrupt the clock (Allada et al., 1998; Darlington et al., 2000). Post-training induction of two independent insertions of the Clkjrk transgene both disrupt 3d memory (Figure 3, Expt #12 and #13), but induction of one of the transgenes prior to training has no effect on 1d memory (Figure 3, Expt #14). In the wild-type fly in which the transgenes reside, post-training induction does not affect 3d memory (Figure 3, Expt #15). The induction regime that is used for these experiments involves two 30’ shifts in temperature (from 20°C to 36°C) approximately 30 and 54 h after the end of the training, with the flies returning to 20°C after the end of each heat-shock. This regimen differs from the one used for the transgenic vri flies (see Figure 2) and demonstrates that brief “pulses” of Clkjrk expression are sufficient to inhibit memory consolidation. We used this acute, shorter-lived induction paradigm to insure that our behavioral effects are not due to flies being exposed to a chronic heat-shock (stress) situation. Western analysis of head extracts from uninduced and induced HS-Clkjrk flies probed with Clk-specific antibody shows induction of a band of the predicted size (see Supplementary Figure 2). Three hours after induction, the transgenic CLKjrk protein is in excess over the endogenous CLK protein (see Supplementary Figure 2), consistent with its seeming ability to inhibit endogenous CLK activity.
Figure 3. Disruption of other circadian genes also interferes with 3d memory. The performance of flies is plotted as a function of genotype, treatment, type of behavioral training, and testing time. Colored histograms (solid or striped) show the performance of HS-Clkjrk (red), w1118 (green), or iso31 (wt), and the Pdp1ε3135 mutation (orange) that resides in the iso31 background. The @ symbol denotes a second, independent insertion of the HS-Clkjrk transgene. Solid colors represent the performance of uninduced (red and green) or wild type (orange; iso31) stocks. Striped histograms show the performance of the induced (red and green) or mutant lines (orange stripes). In the table, the last line is the Experiment #. The genotypes are shown in the top line, followed by whether heat-shock (+) or not was given to the flies. 10 × S corresponds to 10 cycles of spaced training, and 1x indicates flies that received only a single training trial. The testing times were immediate (Imm), 1d, or 3d after the end of training. The horizontal line in the cartoon shows the experimental timeline, and each vertical tick mark represents 1d of elapsed time. Green and magenta arrows indicate whether flies received 10 cycles of spaced training or a single cycle of training. The red arrows show the timing of a heat-shock induction (20°C to 36°C shift for 30’ duration), while the black arrows indicate the time of testing. The color scheme used for the data histograms (top of Figure) is unrelated to the one used in the cartoons (bottom part of Figure). N = 8 for all experiments, T-tests were used to evaluate significance. **p < 0.01. All unmarked pairwise comparisons are not significantly different.
The vri gene encodes a D-box binding repressor, while the Pdp1 gene makes a corresponding activator of transcription (Cyran et al., 2003; Zheng et al., 2009; Gunawardhana et al., 2020). Within the molecular circadian network, VRI and PDP1 are thought to compete for binding to D-box-containing promoters although it is not clear if they necessarily regulate the same ones. In addition to having opposite molecular functions, we chose to test PDP1 because of the existence of the Pdp1ε mutation, Pdp13135. Testing a loss-of-function mutation complements our experiments using transgenic overexpression of proteins (VRI and CLKjrk). The Pdp1 gene encodes multiple protein isoforms, and Zheng et al. (2009) previously showed that Pdp1ε is the circadian isoform specifically disrupted in the Pdp13135 mutant. Since overexpression of VRI disrupts 3d memory, we expect that decreased PDP1 protein should do likewise. When compared to its isogenic wild-type strain, the Pdp13135 mutant shows aberrant 3d memory (Figure 3, Expt #16). To rule out a mutant effect on the relevant sensory systems involved in odor avoidance behavior, we compared the wild type and mutant stocks for immediate performance after a single training trial. Although their scores are low, there is no difference between the two stocks, suggesting that the mutant flies have comparable sensory capabilities when compared to their isogenic wild-type stock (Figure 3, Expt #17). Taken together with the vri data, these results suggest the involvement of vri/Pdp1ε in 3d memory. Four different disruptions (DD, induced overexpression of VRI, induced overexpression of CLKjrk, and removal of PDP1ε) all affect the same temporal window (1d-3d memory). None of the three interventions that were tested affected 1d memory. Based on this data, we believe that disruption of the circadian system very likely affects 1d-3d memory.
HS-vri induction affects sleep but does not disrupt itIt is generally accepted that sleep is essential for memory formation in both vertebrates and flies (Walker and Stickgold, 2004, 2006; Ganguly-Fitzgerald et al., 2006; Bushey et al., 2007; Seugnet et al., 2008; Donlea et al., 2011; Gerstner et al., 2011; Berry et al., 2015; Dissel et al., 2015; Dudai et al., 2015; Haynes et al., 2015; Liu et al., 2019; Chouhan et al., 2021). Decreases in the total amount of sleep, and/or deterioration in the quality of sleep are hypothesized to interfere with consolidation. Sleep is believed to be under homeostatic and circadian control (Borbely, 1982). Therefore we examined whether our behavioral deficits might result from disruptions to sleep. Since mild induction of the HS-vri transgene specifically affects 3d memory, we used the same induction protocol (20°C to 28°C chronic temperature shift) as was used for behavior to test for effects on sleep. We measured the effect of induction (Figure 4, indicated with the vertical dotted line) on sleep in the transgenic HS-vri male flies, as well as in isogenic control flies devoid of the transgene. Their hourly average sleep (+/–SEM) was plotted as a function of time, and Figure 4 shows that transgene induction (red trace) has small effects on the “architecture” of sleep. The transitions between active and inactive periods are identical, as are the kinetics (slopes) of “falling asleep or waking up.” Transgenic flies exposed to heat-shock sleep more during the daytime periods. As a comparison, the isogenic “wild type” stock (w1118) shows very similar effects, although these flies have less saturated sleep during both the pre- and post-induction nighttime.
Figure 4. Effect of HS-vri induction on sleep architecture. The effect of induction on sleep architecture was measured for four different groups (w1118 and HS-vri males, pre- and post-induction). The average minutes of sleep/hour is plotted as a function of elapsed time. For both genotypes, sleep was measured under 12-h light:12-h dark conditions, indicated with the white and black horizontal bars. Flies were entrained to a 12:12 schedule and then loaded into the DAMS monitors maintained in two different 20°C incubators with identical settings. Baseline sleep was assayed for 2 days before one of the incubators was shifted to 28°C and sleep measurements continued for flies at both temperatures. The timing of the heat-shift is shown with the dotted horizontal line. The blue and red curves represent the amounts of sleep for the non-induced and induced flies respectively.
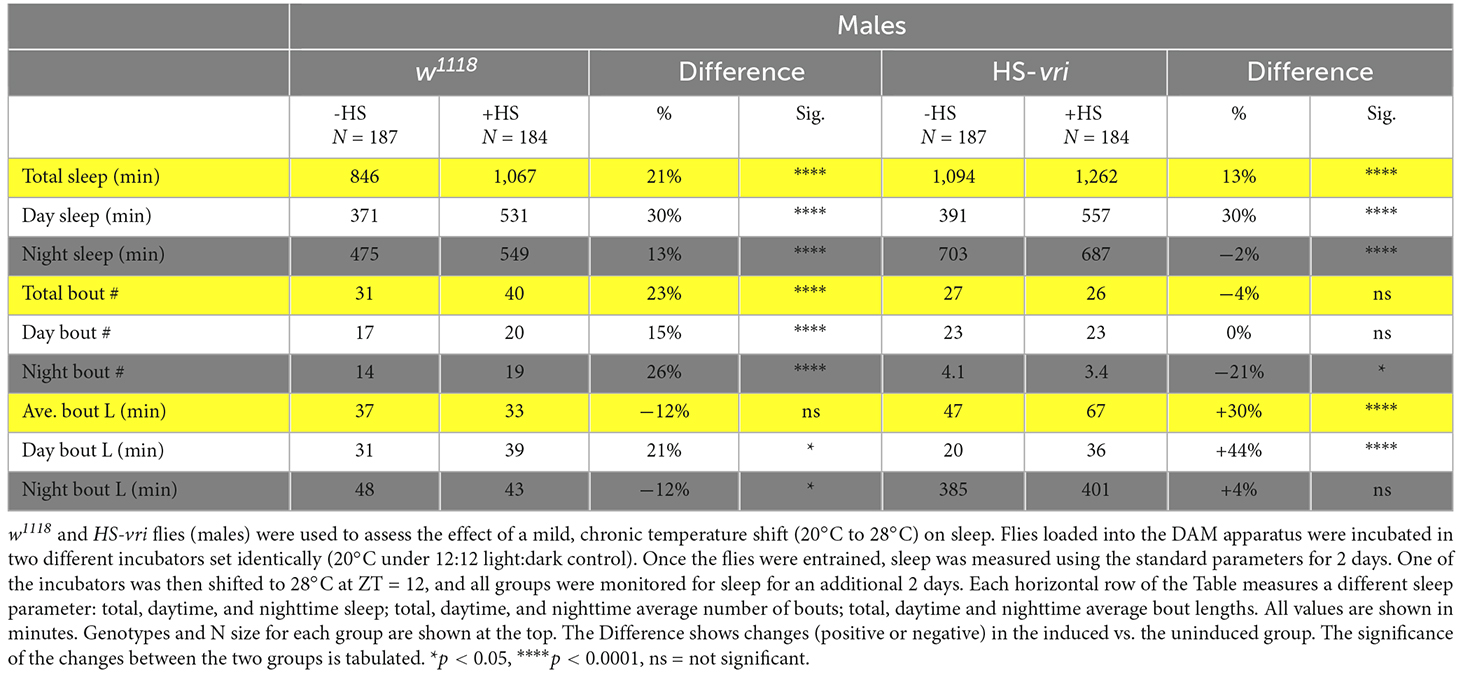
The quantitative analysis of sleep is summarized in Table 1. For the HS-vri transgenic fly, induction increased the total amount of sleep, and this was attributable to an increase in daytime sleep. There was little effect on the total number of sleep bouts, with a small decrease in the average number of nighttime bouts that was statistically significant. In contrast, the average total length of bouts, especially those during the daytime sleep period, increased. Thus the large effects of induction were to increase the amount of sleep, and most of this occurred through increasing the average daytime length of bouts. Since transgene induction disrupted memory formation, and increasing sleep is usually thought to “enhance” memory formation, it is very unlikely that our behavioral disruption results from effects on sleep.
Table 1. Effect of HS-vri induction on sleep.
For comparison, we have also measured sleep changes in the isogenic w1118 strain that is the background in which the HS-vri transgene resides. In general, the effects of heat-shock trend in the same direction, with increases in the total amount of sleep during both daytime and nighttime periods. The average number of bouts increase (total, daytime and nighttime), while the average bout lengths (total, daytime, and nighttime) show barely significant changes. Regardless of these subtle changes, there is no measurable effect of heat-shock in w1118 flies on memory formation.
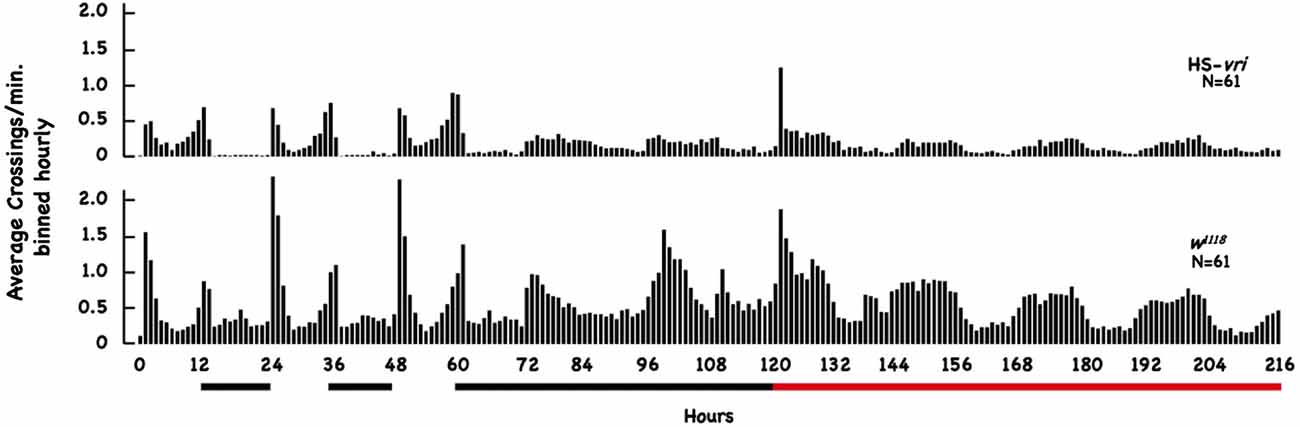
HS-vri induction mildly affects circadian locomotor activityTo evaluate the effect of our transgenic manipulations on circadian behavior, we tested the effect of HS-vri transgene induction on locomotor activity using the DAM apparatus across a 9-day period. Figure 5 plots the average number of beam crossings per minute binned hourly across the entire 216 h of elapsed time. This experiment involves two transitions, one from LD to DD (beginning at 60 h of elapsed time), and the other when heat-shock was begun (at 120 h elapsed time). The black bars below the plot show the nighttime periods and the beginning of constant D, and the red horizontal line indicates when chronic heat-shock (a 20°C to 28°C shift) was begun (in flies under constant darkness).
Figure 5. Effect of HS-vri induction on locomotor activity. Flies (w1118 and HS-vri) were placed into the DAM apparatus to measure locomotor activity. The average number of beam crossings per minute binned in hourly units was calculated and plotted as a function of elapsed time. The black horizontal lines under the Figure indicate dark periods (hours 12–24 and 36–48) and constant darkness (hours 60–120). The red bar indicates constant darkness plus heat-shock (a 20°C to 28°C chronic shift beginning at the 120th h). The N sizes for each group are shown.
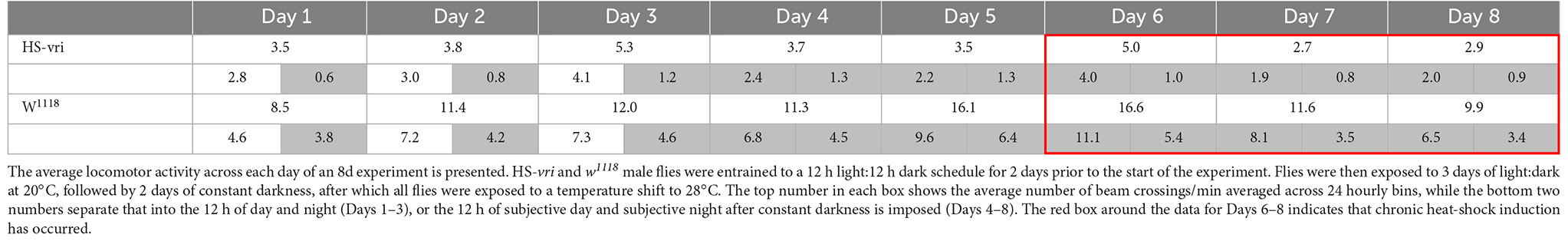
The HS-vri transgenic flies maintained under LD conditions show the classic “U-shaped” activity profile, with peaks at dawn and dusk. Upon the shift to DD, they almost immediately transition to a single peak in activity spread out evenly during the subjective “daytime.” Heat-shock results in an immediate spike in activity, decreases subsequent total activity slightly, but does not change the architecture (shape) of the activity profile. The w1118 flies have quantitative differences in their activity (when compared to HS-vri) but exhibit a similar sleep profile (shape of the curve). Under LD conditions, they show a dramatic U-shaped curve, with greater total and peak activities than the HS-vri flies. The shift to constant darkness does not change the architecture as quickly, but by the time heat-shock is delivered, they show the inverted U-shaped, single peak that is broadly distributed across the subjective daytime. The w1118 flies have a noticeably higher activity level than that of the transgenic flies. In terms of our behavioral results, the only manipulation (+HS in the HS-vri flies) that affects memory formation modestly decreases locomotor activity. The effect of heat-shock on locomotor activity in the w1118 flies is greater but does not affect memory formation. Table 2 presents a more quantitative analysis of the locomotor activity depicted in Figure 5. The white and gray shading represent light and dark periods, while the red box indicates flies that are maintained under DD conditions but at the inducing temperature of 28°C. The top number in each box indicates the total average number of beam crossings (per minute binned in hourly units) in a 24 h period, while the second line in each box breaks this into the daytime and nighttime (shown in gray) periods. Once DD is imposed, the activity is divided between the subjective day and night periods (all shown in gray). The induced HS-vri flies (Days 6–8) are less active than when they were maintained under constant dark only (Days 4–5), and these flies are less active than their w1118 counterparts under all conditions. Although affected, the decrease in the activity of the induced HS-vri flies (vs. the uninduced flies) is not a large effect, and is unlikely to contribute to their behavioral differences. We do not think the induced flies are just more “lethargic,” and less likely to move correctly at the behavioral choice point since they perform similarly to their uninduced controls after a single cycle of training (Figure 2 Expt #8), or after 10 cycles of spaced (Figure 2, Expt #9) or massed training (Figure 2, Expt #10) tested 1d after training. Their only deficit in performance is when it is measured at 3d after 10 cycles of spaced training (Figure 2, Expt #6).
Table 2. Effect of HS-vri induction on locomotor activity.
ConclusionsOur data using the imposition of post-training dark periods, inducible transgenes and a mutant provides strong evidence that circadian molecules, and probably the whole circadian system, is involved in Drosophila memory formation. Our effects seem specific to later memory, since immediate performance after a single training trial (learning), 1d memory after 10 spaced or 10 massed cycles of training, and retrieval are not affected. Since some, or all these different training regimens are hypothesized to affect different “phases” of memory, circadian disruption seems specific to late long-term memory (Tully et al., 1994; Yin et al., 1994, 1995). The same “circadian window” (1d-3d post-training) is affected with all our manipulations (DD, chronic or acute induction of vri or Clkjrk, mutant removal of Pdp1ε). This data, taken together with that from Inami et al. strongly argue for the importance of circadian regulation during memory formation in two different behavioral paradigms (Inami et al., 2020). In both behaviors, later long-term memory seems to be specifically affected. Our data add important kinetic and molecular information on circadian genes likely involved in this requirement. Some of our behavioral effects require heat-shock driven transgenes that may express VRI or CLKjrk in cells that do not normally express these proteins. This ectopic expression could indirectly affect behavior. We think that this likelihood is low since DD and the pdp13135 mutation have similar behavioral effects without any ectopic expression. Future experiments using more limited gene targeting will rigorously eliminate this possibility. It is also possible that the transcriptional targets whose gene expression is altered when HS-vri (or HS-Clkjrk) are induced and affect memory formation are outside of the circadian system. We think that this possibility is likely, but gene expression analyses in the memory cells will be needed to test this idea. The DNA sequences that CLK (E-boxes) and VRI/PDP1 (D-boxes) bind to are found broadly in promoter regions (Vinson et al., 2011; Ishibashi et al., 2019; Gunawardhana et al., 2020). Current efforts are focused on identifying when and where the circadian molecules are required.
Perhaps one of the surprising aspects of our data is that 1d memory does not seem to require the clock. Previously, both Sakai et al. and Chen et al. had reported that circadian mutants did not affect early memory formation using two different behavioral assays (Sakai et al., 2004; Chen et al., 2012). We have confirmed those findings using inducible transgenes, thus eliminating one possible source for the previously reported lack of effects. Since our transgenes affect certain memory “phases” (3d) but not others, we are confident that the reagents are effective. However, Lyons and Roman reported that learning, or immediate performance after a single training trial, is susceptible to “time-of-day” (TOD) effects (Lyons and Roman, 2008). We previously reported that the TOD also modulates 1d memory (Fropf et al., 2018). Intriguingly, the perS mutation, which exhibits a 19 h periodicity in locomotor behavior, may alter this preferred TOD of training (Fropf et al., 2018). These findings suggest that the circadian system, or at least some of the circadian molecules, are involved in influencing TOD preferences. It is unclear how the circadian system exerts TOD modulatory effects on learning and 1d memory, but “loss-of-function” mutations, or induced transgenes do not disrupt 1d memory. We believe that defining the anatomical and temporal requirements for the TOD preference, and 1d and 3d memory, will likely resolve some of the current contradictory results.
At face value, our finding that 3d memory is dependent upon an intact circadian system (while other phases are not) hints at significant differences between the mechanisms that support memory at those different timepoints. Since protein synthesis is likely needed for 1d memory, the subsequent requirement for circadian genes suggests a broader context in which memory consolidation occurs between 1–3d after training. This later requirement for circadian transcription is consistent with the emerging view that there are multiple “waves” of gene expression during consolidation in both flies and rodents (Hirano et al., 2016; Chen et al., 2020; Mizuno et al., 2020). These “waves” could contribute to features of memory formation, such as systems consolidation, that occur over a more prolonged period of time (Kim and Fanselow, 1992; Dudai, 2004, 2012). Our kinetic and molecular information on circadian genes that are likely involved in memory consolidation may provide important temporal windows and tools to investigate these events.
Another surprising result is that constant light (LL) does not disrupt memory consolidation, as first reported for courtship suppression memory (Inami et al., 2020). Our data completely supports the view that light itself is a necessary factor during memory formation. While LL treatment is known to interfere with the oscillations of the circadian molecular machinery, it does not affect memory formation. On the other hand, DD does not interfere with these molecular oscillations (Hardin et al., 1990; Qiu and Hardin, 1996) but disrupts memory formation. How these paradoxical findings can be resolved will likely require identification of the participating cells and an understanding of when the cells are needed during consolidation. One possible resolution is that memory formation recruits a process that is used in neural development, and this process requires light (Dapergola et al., 2021; Damulewicz et al., 2022).
How are our manipulations affecting memory consolidation? The two obvious neurobiological processes that might be targeted are sleep and circadian rhythms. However, all the measurable changes in response to HS-vri induction—increases in the total amounts of sleep, decreases in the average number of bouts, and increases in the average bout lengths—are usually associated with better sleep. Therefore, it seems highly unlikely that HS-vri induction is disrupting sleep and thus affecting memory consolidation. Induction of HS-vri has similarly small effects on the circadian regulation of locomotor activity. The overall “architecture” of activity (more activity during the subjective daytime and the least amount of activity near the end of the subjective night, and a single distributed peak in activity across a 24 h period) remains intact after heat-shock induction. The clear large effects of HS-vri induction on memory consolidation contrast with the minor effects on locomotor activity. We suspect that HS-Clkjrk, loss of Pdp1ε, and DD likewise have negligible effects on sleep and circadian locomotor activity, consistent with what Inami et al. reported for DD. It seems more likely that our effects are mediated by circadian involvement in “other” processes, perhaps ones that recruit “peripheral clocks” (Hardin et al., 2003; Ito and Tomioka, 2016; Sehgal, 2016; Di Cara and King-Jones, 2016; Selcho et al., 2017; Yildirim et al., 2022). Although our results do not clarify all the outstanding issues, they do provide an experimental template going forward to help resolve the current uncertainties.
Regardless of the mechanisms that circadian intervention affect during memory formation, our data and that of Inami et al. clearly show that light itself, and the circadian clock are required for memory formation (Inami et al., 2020, 2021). Inami et al. (2022) believe that circadian functions are involved in memory maintenance and we conceptually agree with this intuitively appealing idea. However, we believe that the “circadian window” described in this report is affecting an earlier part of memory formation. Future experiments will be needed to clarify the cells and the timepoints when maintenance occurs, and how the circadian system contributes to this process.
Data availability statementThe original contributions presented in the study are included in the article/Supplementary materials, further inquiries can be directed to the corresponding author.
Ethics statementThe research is conducted with Drosophila, and the University of Wisconsin-Madison Office of Vice Chancellor for Research and Graduate Education does not require ethical approval for work with invertebrates.
Author contributionsJY conceptualized, designed and helped acquire the data, analyzed the data, and wrote the manuscript. EC did the sleep and circadian experiments and wrote the Excel program to analyze the sleep and circadian data. PH helped write the manuscript, supplied key reagents and consulted on experimental design. HZ did the research experiments and helped analyze the data. All authors contributed to the article and approved the submitted version.
FundingThe experiments began in the Yin lab more than 25 years ago and were supported by NIH NS35575, HL/AR59649 (with Amita Sehgal), DA015753, MH067774, and NS063245, funds from the McKnight Foundation, FRAXA, CHDI/HQ Foundation, and AHAF/ADR. Recent research was partially supported through funds from the International Division of UW-Madison, and a UW-Madison OVCRGE Fall Competition Award. Research in the Hardin lab was supported by the John W. Lyons’59 endowed Chair.
AcknowledgmentsWe acknowledge flies and advice given over the years from Amita Sehgal, Paul Taghert, F. Rob Jackson, and Mike Young.
Conflict of interestThe authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s noteAll claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary materialThe Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnsys.2023.1129152/full#supplementary-material.
SUPPLEMENTARY FIGURE 1 | Post-training DD inhibits 3d memory in red-eyed flies. Uninduced HS-Clkjrk (red-eyed) flies were entrained to a 12 h light:12 h dark schedule at 20°C. Flies were trained with 10 cycles of spaced training beginning around ZT = 14. In Experiment S1, the flies were incubated after training in light:dark until Day 2, at which time half of the flies (whose subsequent performance is shown with a black histogram) were exposed to dark during the D2 period and then returned to light:dark until they were tested at 3 days post-training. The other half of the flies remained on light:dark throughout and were also tested for 3d memory (hatched histogram). In Experiment S2, the experimental flies (black histogram) were shifted to constant darkness at the beginning of D2 and remained in darkness until testing. Control flies (hatched histogram) were put on LD after training and remained under those conditions until testing. **p < 0.01, ***p < 0.001, N = 8, T-test.
SUPPLEMENTARY FIGURE 2 | Western analysis of HS-Clkjrk induction. HS-Clkjrk transgenic flies were entrained to a 12 h light:12 h dark schedule at 20°C. Flies in groups of approximately 100 were induced, or not, in empty food vials placed into a water bath at 36°C for 30’ at ~ZT = 7. Uninduced flies were handled, transferred to vials, but not induced. After induction (or handling only), flies were returned to food vials and incubated at 20°C for 3 h, when they were collected and flash frozen in 15 ml polypropylene tubes. The tubes were shaken and pounded, and heads were isolated using a series of sieves. 50 heads were counted out (over powdered dry ice), pulverized using a dounce-like plastic pestle, and extracts made in standard 2× Laemmli SDS loading buffer. About 15 fly head-equivalents were loaded onto a 5% polyacrylamide gel, subjected to electrophoresis, processed for western analysis using a CLK-specific antibody (gp50; Houl et al., 2006, 2008) that was used at a 1:2,000 dilution. The secondary antibody (LI-COR) was a donkey anti-guinea pig IgG conjugated with a fluor and used at a 1:20,000 dilution. The mobility of the molecular weight markers (in kD) is indicated to the left of the image. Based on mobility, the endogenous CLK and transgenic CLKjrk bands are indicated. Imaging and quantitation was done on the LI-COR Odyssey system. For quantitation, background counts (from a region of the gel that did not contain protein samples) was subtracted from both the presumed CLKjrk and endogenous CLK bands. The resulting CLKjrk intensity was greater than the adjusted intensity for the endogenous CLK band at the 3 h post-induction time point.
ReferencesAllada, R., White, N. E., So, W. V., Hall, J. C., and Rosbash, M. (1998). A mutant Drosophila homolog of mammalian Clock disrupts circadian rhythms and transcription of period and timeless. Cell 93, 791–804. doi: 10.1016/s0092-8674(00)81440-3
PubMed Abstract | CrossRef Full Text | Google Scholar
Barnes, C. A., McNaughton, B. L., Goddard, G. V., Douglas, R. M., and Adamec, R. (1977). Circadian rhythm of synaptic excitability in rat and monkey central nervous system. Science 197, 91–92. doi: 10.1126/science.194313
PubMed Abstract | CrossRef Full Text | Google Scholar
Berry, J. A., Cervantes-Sandoval, I., Chakraborty, M., and Davis, R. L. (2015). Sleep facilitates memory by blocking dopamine neuron-mediated forgetting. Cell 161, 1656–1667. doi: 10.1016/j.cell.2015.05.027
PubMed Abstract | CrossRef Full Text | Google Scholar
Bruning, F., Noya, S. B., Bange, T., Koutsouli, S., Rudolph, J. D., Tyagarajan, S. K., et al. (2019). Sleep-wake cycles drive daily dynamics of synaptic phosphorylation. Science 366:eaav3617. doi: 10.1126/science.aav3617
PubMed Abstract | CrossRef Full Text | Google Scholar
Bushey, D., Huber, R., Tononi, G., and Cirelli, C. (2007). Drosophila hyperkinetic mutants have reduced sleep and impaired memory. J. Neurosci. 27, 5384–5393. doi: 10.1523/JNEUROSCI.0108-07.2007
PubMed Abstract | CrossRef Full Text | Google Scholar
Cain, S. W., Chou, T., and Ralph, M. R. (2004a). Circadian modulation of performance on an aversion-based place learning task in hamsters. Behav. Brain Res. 150, 201–205. doi: 10.1016/j.bbr.2003.07.001
PubMed Abstract | CrossRef Full Text | Google Scholar
Cain, S. W., Ko, C. H., Chalmers, J. A., and Ralph, M. R. (2004b). Time of day modulation of conditioned place preference in rats depends on the strain of rat used. Neurobiol. Learn. Mem. 81, 217–220. doi: 10.1016/j.nlm.2004.02.003
PubMed Abstract | CrossRef Full Text | Google Scholar
Cain, S. W., Rawashdeh, O. A., Siu, M., Kim, S. C., and Ralph, M. R. (2017). Dopamine dependent setting of a circadian oscillator underlying the memory for time of day. Neurobiol. Learn. Mem. 141, 78–83. doi: 10.1016/j.nlm.2017.03.015
PubMed Abstract | CrossRef Full Text | Google Scholar
Cervantes-Sandoval, I., Martin-Pena, A., Berry, J. A., and Davis, R. L. (2013). System-like consolidation of olfactory memories in Drosophila. J. Neurosci. 33, 9846–9854. doi: 10.1523/JNEUROSCI.0451-13.2013
留言 (0)