
記住我

Cholera still continues as a major public health menace for centuries in developing and underdeveloped countries of Asia and Africa. The abrupt and deadly onset of cholera epidemics and outbreaks have been a persistent health problem in coastal and tribal regions of Odisha, India [9]. This has been exacerbated by unhygienic living practices, inadequate health infrastructures and limited access to potable drinking water. The consumption and utilization of contaminated water from environmental sources like streams, nalas and rivers made the population prone to cholera outbreaks and epidemics in rainy seasons, such as the major outbreaks of 2007 and 2010 in Rayagada district, of 2009 in Kendrapara district and 2014 in Kalahandi district of Odisha [10, 23]. In the mid of 2007, a severe cholera epidemic was reported in the kashipur block of Rayagada district and continued transmitting to the neighboring districts such as Gajapati, Koraput, and Kalahandi leading to be one of the most extreme cholera outbreaks in Odisha [14].
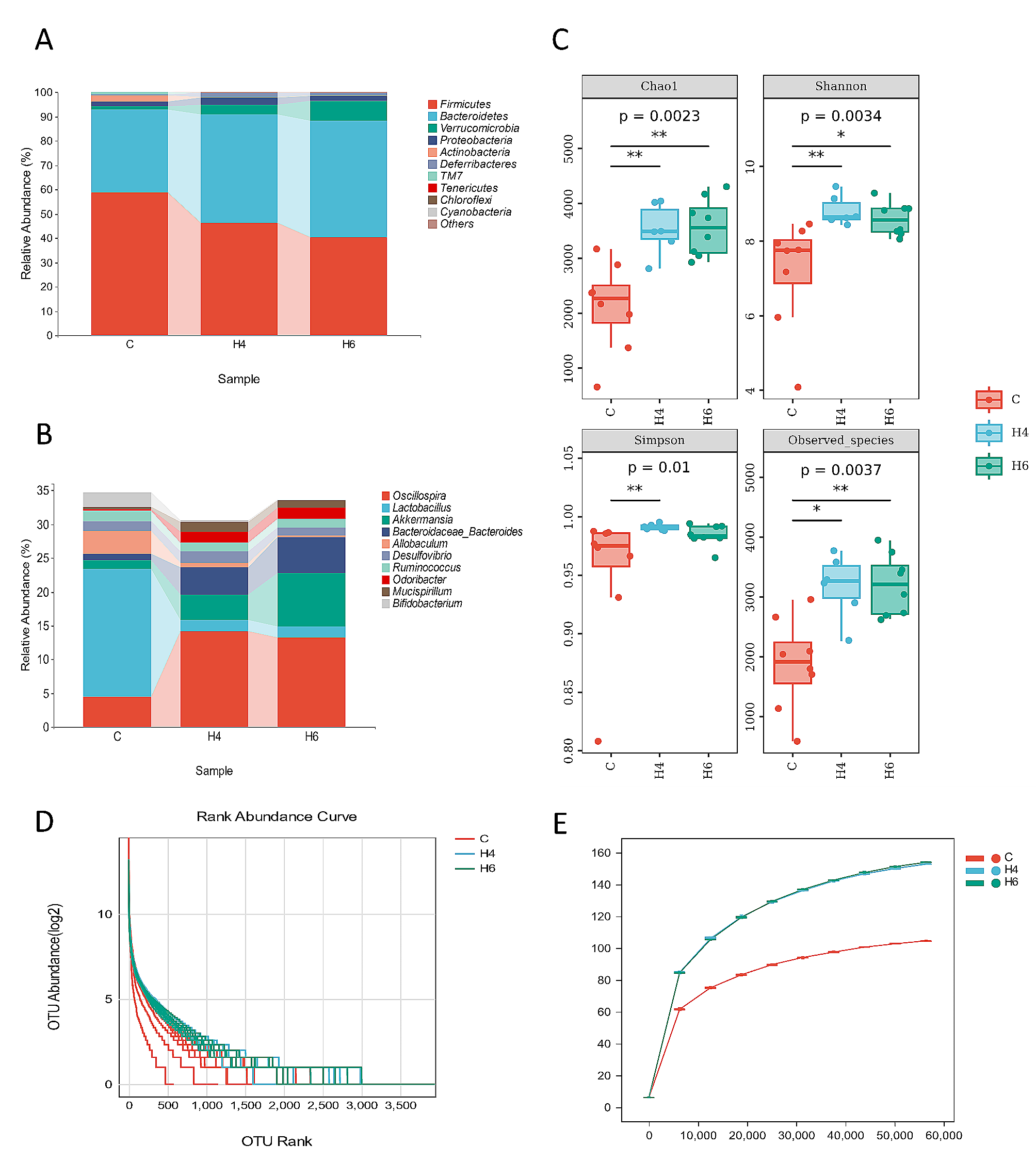
This study provides a comprehensive analysis of the diversities of virulence genes, antibiotic resistance genes and antibiogram trend over more than a decade from 2007 to 2019 along with the co-existence of V. cholerae O1 and non-O1/non-O139 strains from environmental sources like nala, chua, streams, ponds, rivers and road side reservoirs. The results from this study confirmed higher prevalence of V. cholerae O1 than non-O1/non-O139 in environmental water samples which contradicts to the result reported in Bepanda, Douala [24]. At present, the altered El Tor strains of V. cholerae O1 carrying ctxB7 genotypes were identified in all the environmental isolates from 2007 onwards and it predominated over the normal prototype El Tor strains causing all outbreaks and epidemics in Odisha. Our earlier studies proved that these strains carrying ctxB7 genotype was first reported from the coastal district of Odisha during super cyclone of 1999 [10]. Our data also supported the fact that environmental and clinical El Tor strains having ctxB7 gene were simultaneously circulating in the tribal districts of Odisha subsequent to the cholera epidemic in 2007 [12]. However, by correlating with the results from our earlier studies on clinical strains it was evident that both clinical and environmental strains showed 100% prevalence of tcpA Haitian genes from 2007 to 2019 except in years 2011 to 2013 and 2016 where clinical strains fall behind [21].
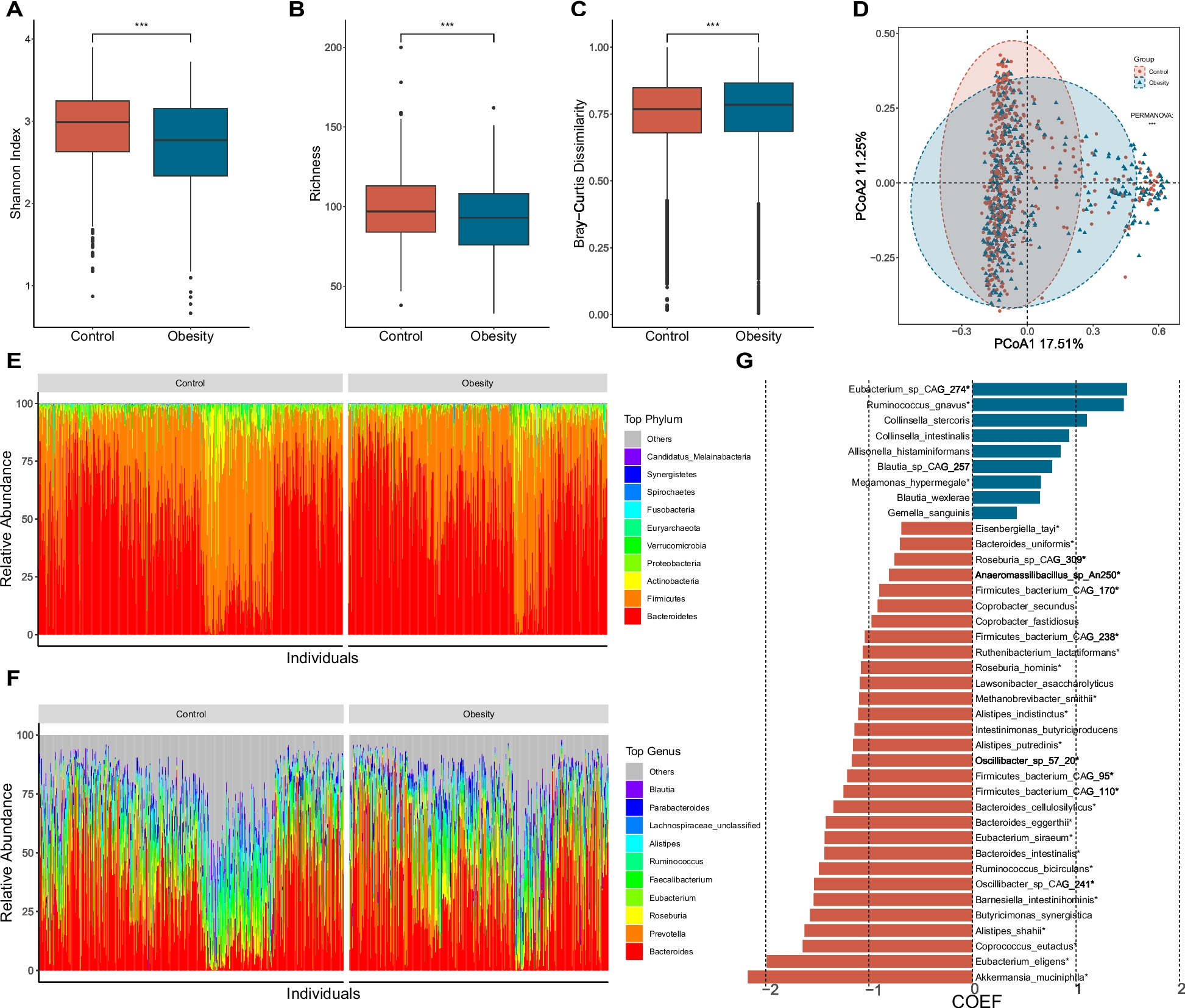
The multiplex PCR-I established the presence of different genes like zot, tcpA, rfbO1, ctxAB, and ompW genes; whereas multiplex PCR-II analysis confirmed the presence of toxR, ompU, hlyA, ace, and rtxC genes. The serological result complemented with the results of multiplex PCR I assay. This assay confirmed that all the 51 V. cholerae strains were toxigenic as they were positive for ompW, rfbO1 and ctxAB genes [16]. However, 20% of the 34 V. cholerae non-O1/non-O139 strains that were serologically negative possessed sero-specific rfbO1 gene confirmed through multiple PCR-I assay, which was an exception. The multiplex PCR-II assays possessing the biotype-specific marker genes like rtxC, tcpA and ctxB revealed that the confirmed V. cholerae O1 were of altered El Tor prototype but mostly possessed Haitian tcpA and ctxB Haitian genotypes. However, the results produced were contradictory to the findings reported in Bepanda in Doula, Accra in Ghana, and Dhaka in Bangladesh, as the prevalence of tcpA El Tor and ctxB1 genotypes were more prominent in these regions [24,25,26]. Generally, the environmental isolates of V. cholerae non-O1/non-O139 strains were incompetent to seed any cholera outbreaks/epidemics; but the seroconversion of V. cholerae non-O1/non-O139 to O1 has heightened attention towards V. cholerae non-O1/non-O139 strains. Though non-O1/non-O139 serogroup of V. cholerae are non-lethal, evidence of genes encoding virulence factors have been identified in environmental strains, which might account for major public health risks [27].
From the present study, it was evident that the prevalence of essential toxic genes such as toxR, zot and ace of V. cholerae O1 strains were 50% and above, along with the accessory genes i.e. rtxC, ompU, ctxAB, hlyA, ompW, rfbO1 and tcpA, which varied in-between 68.6 and 100%. These findings resembled those reported by other researchers from Accra, Ghana in 2019, Bangladesh in 2004 and Lubumbashi, Congo in 2008 [25, 28, 29]. From all the virulence and accessory genes in V. cholerae non-O1/nonO139 tested, only 3 genes were prominent i.e. toxR (94.3%), rtxC (60%), and hlyA (85.7%). Although V. cholerae non-O1/non-O139 strains typically do not possess virulence associated genes, they often contain other virulence genes, which can contribute to their pathogenicity. It has also been reported that both V. cholerae O1 and non-O1/non-O139 isolated from the same aquatic habitat possessed the regulatory protein gene (toxR). Thus it can be speculated that under several biochemical and physiological conditions, horizontal gene transfer might have occurred from toxigenic V. cholerae O1 to non-O1/non-O139 when exposed to bacteriophages. This could lead to the emergence of novel pathogenic V. cholerae strains in the environment causing severe outbreaks/epidemics [30].
The genetic basis for the resistance to antibiotics could be due to lateral acquisition of self-transmissible conjugative SXT element in V. cholerae derived ICE element. The name SXT was given because of its contribution to help bacteria to sustain and grow in the presence of streptomycin, sulfamethoxazole and trimethoprim. SXT ICEs possibly emerged first from the Asian continents during the late 1980s and subsequently spread into several clinically important bacterial species in different countries. Currently, SXT ICEs were detected in almost every V. cholerae O1 isolates across the globe. Genomic studies indicated that V. cholerae O1 and O139 isolates independently acquired the SXT ICEs. [31, 32]. The ICE-associated genes dfrA1, sulII, SXT element and strB were reported as the genetic determinant to carry resistance against multiple drugs such as streptomycin, cotrimoxazole, nalidixic acid and neomycin [33]. In the present study, we focused on the ICE-associated genes SXT, strB, dfrA1 and sulll, which conferred resistance to different serogroups of V. cholerae against multiple drugs. The resistance genes dfrA1 and SXT were mostly prevalent in 94% and 80% of the V. cholerae O1 isolates respectively, in addition, the Sulll and strB genes were also prevalent in 50% and 57% of the strains. V. cholerae harboring these multiple antibiotic markers in the genome displayed resistance against ampicillin, streptomycin, nalidixic acid, neomycin, cotrimoxazole, and furazolidone [34]. The vast spread of dfrA1 and SXT elements in the strains tested resulted in the resistance of these antibiotics. Besides all, the mobile integron often linked with the MGEs (ICE, insertion sequence, transposons, conjugative plasmids) is also responsible for the dissemination of multiple antibiotic resistance encoding functions to the environmental V. cholerae strains. However, MGEs were the pre-existing source of antimicrobial-resistant genes which mediated the exchange of antibiotic resistance genes between pathogens or between pathogens and non-toxigenic commensals of bacterial populations which were existing in common environmental niches [32]. Thus mobile integrons, ICE, and conjugative plasmids could also attribute the genetic basis for multi-drug resistance in V. cholerae strains.
Effective treatment in cholera cases can be only possible through aggressive oral or intravenous re-hydration therapy along with the ideal course of antibiotics. This can significantly shorten the duration of diarrhea and reduce the loss of electrolytes in the patients’ body [3]. However, difficulties in antibiotic therapy have evolved in recent years due to the rapid emergence of multi-drug resistant (MDR) strains of V. cholerae in different serogroups from Asia, Africa, and America [35]. Over the past two decades, progressive increasing trend of V. cholerae O1 resistance against multiple drugs such as tetracycline, streptomycin, norfloxacin, ciprofloxacin, trimethoprim/sulfamethoxazole, ampicillin, neomycin, cotrimoxazole, nalidixic acid, and furazolidone were reported in Odisha and India [21]. The antibiotic susceptibility assay revealed loss of sensitivity to the tested antibiotics including nalidixic acid, streptomycin, co-trimoxazole, furazolidone, ampicillin and neomycin which were used in the first line of defense against cholera. Similar incidences of resistance patterns were also reported in Nepal in 2015, Ghana in 2019, and the Democratic Republic of Congo in 2015 [25, 36, 37]. In the present study, both V. cholerae O1 and non-O1/non-O139 were completely resistant to ampicillin, furazolidone, and streptomycin. A higher percentage of resistance from 60 to 80% against azithromycin, cotrimoxazole, nalidixic acid and neomycin in V. cholerae O1 strains were observed since 2007. These results coincided with the reports from Nepal and Mozambique [38, 39]. Tetracycline-resistant V. cholerae O1 in clinical strains were reported only in 2010 from tribal areas of Odisha, whereas in the present study on environmental isolates both V. cholerae O1 and non-O1/non-O139 strains were resistant to tetracycline on year 2010 and 2011. However, it was reversed in successive years [40]. The environmental isolates of V. cholerae during cholera epidemics and outbreaks were observed having higher antibiotic resistances and this might me due to the transmission of multi-drug resistant V. cholerae O1 strains from human to water bodies. The drug resistant patterns of both clinical and environmental strains varied over space and time.
In this study, it was discovered that 66.6% of all the V. cholerae O1 isolates were resistant to at least 7 out of 14 antibiotics screened and possessed a MAR index value ranging between 0.5 and 0.79. However, 73.5% of the non-O1/non-O139 were sensitive to more than 8 antibiotics screened. The highest MAR values were reported in 2007 and 2012; as 10 out of 14 antibiotics were resistant to V. cholerae O1 environmental water isolates. This could be due to prolonged usage of antibiotics for curative and prophylactic purposes in cholera-affected patients during the outbreaks and epidemics occurred in the Rayagada, Gajapati and Kalahandi districts of Odisha. This might have provoked the emergence and spread of MDR V. cholerae strains in the environment. However, the reversal in the resistance of these antibiotics against V. cholerae was also observed in successive years. This phenomenon might be due to rapid fluctuation in nature and the inability of V. cholerae to carry plasmids that conferred resistance to it [41].
NotI digested genomic DNA of V. cholerae O1 from water isolates were analyzed by PFGE, whose patterns revealed diversity of their genomes containing 3 different pulsotypes with a similarity matrix of 93%. The PFGE banding patterns in 8 strains were highly homogenous showing 100% similarity indicating origin from a single clone. Whereas, the remaining 3 pulsotypes of 2012 and 2018 possessing ctxB1 and ctxB7 genotypes respectively were distinct from the other strains. The present study also indicates that the environmental isolates of V. cholerae were the precursors of all the outbreaks and epidemics of cholera in Odisha [26].
Thus, continuous surveillance is necessary to monitor the different water bodies to study the changing over of biotypes and serotypes of V. cholerae different serogroups which will give early warning system for the control of cholera outbreaks/epidemics in this area. Again, in-depth molecular study on the toxic genes, drug resistance patterns including the ctxB genotypes of V. cholerae strains from coastal and tribal areas of Odisha is highly essential.
Table 1 Prevalence of virulence genes in Vibrio cholerae O1 and non-O1/non-O139 strains isolated from environmental sources in different districts of Odisha: 2007–2019 Table 2 Annual antibiogram profiles of Vibrio cholerae O1 and non-O1/non-O139 strains isolated from environmental sources in different districts of Odisha: 2007–2019 Fig. 2
Heat map showing percentage variation of different virulence genes among Vibrio cholerae O1 and non-O1/non-O139 strains over the years
Fig. 3
(a): Prevalence of different virulence genes like ctxB and tcpA genes in Vibrio cholerae O1 strains from environmental water isolates. (b): Prevalence of different virulence genes like ctxB and tcpA genes in Vibrio cholerae non-O1/non-O139 strains from environmental water isolates
Fig. 4
Distribution of antibiotic resistant genes in Vibrio cholerae O1 and non-O1/non-O139 strains represented in doughnut chart
Fig. 5
Multiple antibiotic resistance (MAR) Index of Vibrio cholerae O1 isolates represented in Box whisker plot
Fig. 6
Antibiogram profiles of Vibrio cholerae O1 and non-O1/non-O139 strains represented in stacked column chart [gentamicin (G), ciprofloxacin (Cf), norfloxacin (Nx), azithromycin (At), ofloxacin (Of), doxycycline (Do), ampicillin (A), chloramphenicol (C), furazolidone (Fr), cotrimoxazole (Co), streptomycin (S), nalidixic acid (Na), neomycin (N) and tetracycline (T)]
Fig. 7
PFGE analysis of NotI digested Vibrio cholerae O1 strains isolated from environmental sources in different districts of Odisha: 2007–2019
留言 (0)