
記住我
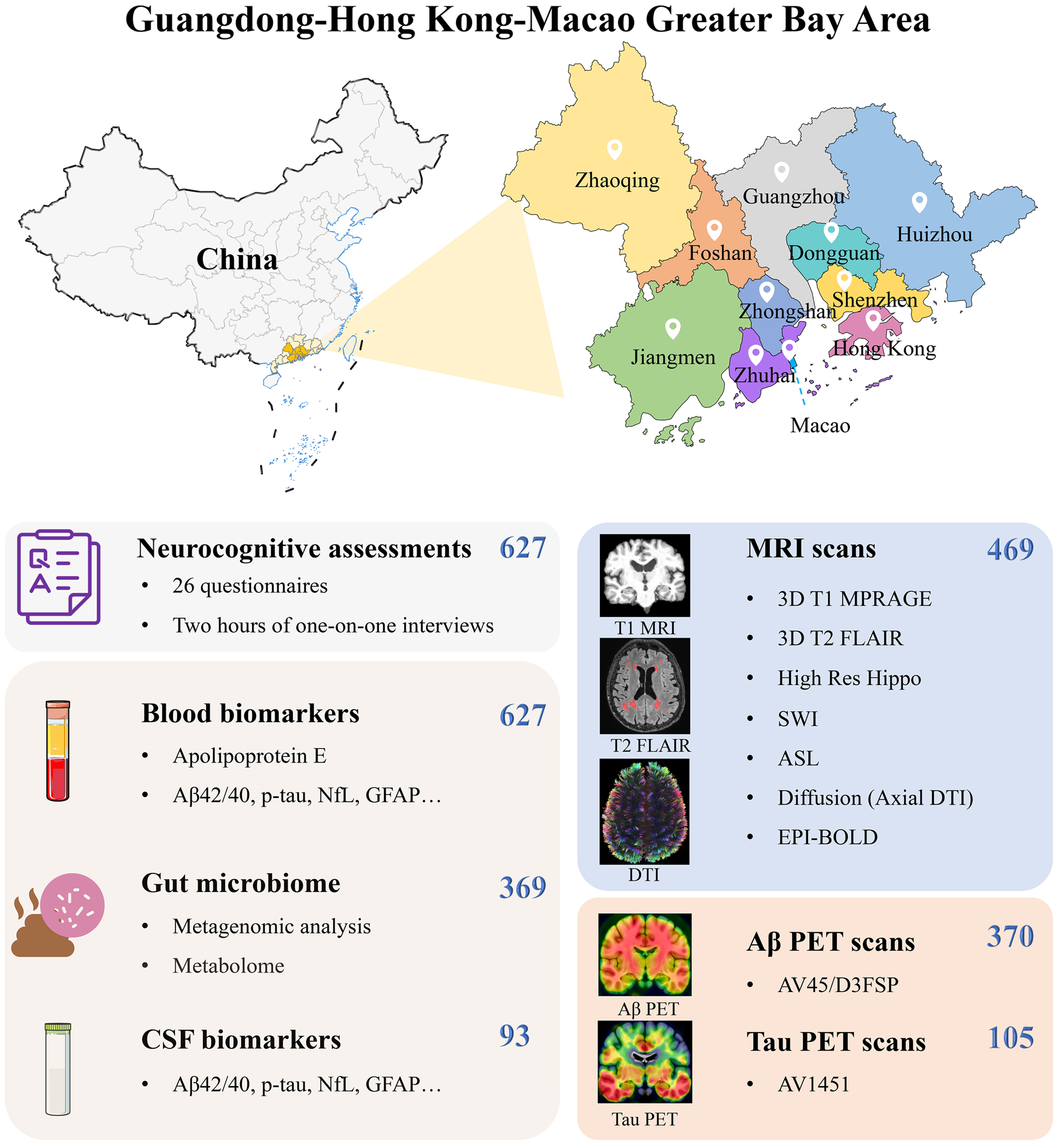
To further address whether AD-relevant mouse models show any skeletal bone deficit and to investigate whether PTH1-34 treatments affect AD-associated bone deficits, we used 5XFAD mouse, a well-characterized AD animal model that expresses mutant human APP and presenilin genes under the control of Thy1-promoter, and exhibit early onset Aβ based brain pathologies (~ 2 MO) and cognition deficits (~ 4 MO) [1, 29, 39]. C57BL/6J mice were used as WT controls. We first examined bone phenotypes by microCT (μCT) analyses in femurs of control/WT and 5XFAD female mice, in light of reports of earlier onset and severer phenotypes in female than male 5XFAD mice [29, 40]. The control/WT and 5XFAD mice were treated with Vehicle (0.9% NaCl) or PTH1-34 intermittently (once per day and five days per week), starting at age of 2-MO and sacrificed at ~ 6-MO, as illustrated in Fig. 1A. Notice that in the absence of PTH1-34, μCT examinations showed reductions in trabecular bone volumes (Tb. BV/TV) and trabecular bone thickness (Tb. Th), without changes in their trabecular bone numbers (Tb. N) and cortical bone volumes (Cb. BV/TV), in 5XFAD mice (at age of 6-MO), as compared with those of litter mate control/WT mice (Fig. 1B–F), suggesting a bone deficit in 5XFAD mice. In line with this view were observations that serum levels of osteocalcin (a marker for bone formation) were decreased, but PYD (pyridinoline) (a maker for bone resorption) levels were increased, in 5XFAD mice (Fig. 1G–H). These results thus implicate both reduced bone formation and elevated bone resorption to underlie the trabecular bone-loss in 5XFAD mice.
Fig. 1
Trabecular bone loss in 5XFAD mice was diminished by PTH1-34 treatments. A Schematic of hPTH1-34 or vehicle (0.9% sodium chloride) intermittent treatment and tissue samples collection in WT/5XFAD female mice. Female WT/5XFAD mice were administered a once-daily injection of hPTH1–34, or veh (0.9% NaCl) via subcutaneous injection, starting at 2 ~ MO old, 5 days per week. Mice were sacrificed at 6 ~ MO old to detect bone phenotypes. B–F μCT analysis of femurs from 6 ~ MO WT and 5XFAD female mice with PTH1-34 or Veh treatment. Representative images are shown in B and quantification analyses of trabecular bone volume over total volume (Tb. BV/TV), trabecular bone number (Tb. N), trabecular bone thickness (Tb. Th), and cortical bone volume over total volume (Cb. BV/TV) by the direct model of μCT analysis are presented in B–F. G Serum osteocalcin levels analyses, measured by ELISA assays, in 6 ~ MO female mice. H Serum PYD levels analyses, measured by ELISA assays, in 6 ~ MO female mice. Three different female mice from each group were examined. The data were presented as mean ± SD, *P < 0.05, **P < 0.01, ***P < 0.001, two-way ANOVA with two-stage step-up method of Benjamini, Krieger, and Yekutieli multiple comparisons test was used
Interestingly, upon PTH1-34 treatments, more trabecular bone mass was induced in 5XFAD mice than that in WT mice (Fig. 1B–C), suggesting an enhanced PTH1-34 response in 5XFAD mice. In line with this view were the observations that PTH1-34 increased serum osteocalcin levels more in 5XFAD mice than that in control mice (Fig. 1G), but had little to no effect on serum PYD levels in 5XFAD mice, as compared to that of WT controls (Fig. 1H). These results thus suggest that the PTH1-34-induced trabecular bone mass in 5XFAD mice is likely due in large part to the elevated bone formation. The two-way ANOVA analysis showed that PTH1-34 treatment has a significant effect on various measures of bone phenotypes (P < 0.05) and that genotype has a significant effect on Tb. BV/TV, serum osteocalcin levels, and serum PYD levels (P < 0.05) (Supplemental file 1 Table 1). Significant interactions between PTH1-34 treatment x genotype have been observed in Tb. BV/TV (F = 27.03, p = 0.0008), serum osteocalcin levels (F = 26.59, p = 0.0009), and serum PYD levels (F = 17.01, p = 0.0033) (Supplemental file 1 Table 1). These results suggest an enhanced bone metabolism in response to PTH1-34 in 5XFAD mice.
To understand how PTH1-34 induced anabolic response is enhanced in 5XFAD mice, we measured mouse PTH levels in control and 5XFAD female mouse serum samples and brain (cortex and hippocampus) homogenates (at age of 6 ~ MO). Interestingly, ELISA showed lower levels of PTH in 5XFAD serum and hippocampal samples than those of controls but comparable levels of PTH in 5XFAD cortex to that of controls (Fig. S1A-B). These results suggest that PTH is detectable in the brain, but it is reduced in 5XFAD serum and hippocampus. Such a PTH-deficiency may underlie the enhanced response to the injected hPTH1-34.
PTH1-34 attenuation of cognition decline and memory-loss in 5XFAD miceAD is a progressive neurodegenerative disease commonly associated with memory deficits and cognitive decline [41]. We thus asked whether PTH1-34 treatments could improve cognitive function in 5XFAD mice. WT or 5XFAD female mice were administered with hPTH1–34 or Veh intermittently as illustrated in Fig. 2A, and behavioral experiments began when the mice were ~ 5-MO. Novel object recognition (NOR) and Morris water maze (MWM) tests were used to access mouse cognitive function or learning and memory [42], and Y-maze was used to test mouse working memory [43]. While the cognitive function was substantially reduced in 5XFAD female mice compared to WT mice of the same age, these declines were largely brought down by PTH1-34 (Fig. 2B–H). Upon treatments with PTH1-34, 5XFAD female mice performed significantly better during these tests, as compared with those of 5XFAD control mice, exhibiting improved recognition of the novel object (Fig. 2B–C), faster learning of the hidden platform during MWM tests (Fig. 2E), and better spatial memory of the platform location (Fig. 2F–H). Additionally, these PTH1-34 treated 5XFAD female mice showed an increase in spontaneous alternation, but comparable arm entries to those of Veh-treated 5XFAD female mice, during Y-maze tests (Fig. 2D), suggesting an improvement in spatial working memory. The two-way ANOVA analysis showed the effect of genotype (5XFAD) in the Y maze (spontaneous alternation: F = 4.283, P = 0.0499) and MWM test (target zone crossovers: F = 7.363, P = 0.0124; time in target quadrant: F = 3.095, P = 0.0918), while the significant effect of PTH1-34 treatment only detected in the Y maze (spontaneous alternation: F = 6.832, P = 0.0155). There were interactive effects between PTH1-34 treatment x genotype on NOR (discrimination index: F = 6.379, P = 0.0189) and MWM test (target zone crossovers: F = 5.417, P = 0.0291; time in target quadrant: F = 11.15, P = 0.0028) (Supplemental file 1 Table 2). The PTH1-34 improvement of the cognitive function in 5XFAD mice appeared to be more pronounced in females than males (Fig. 2 and Fig. S2). Little to no improvement in cognitive function in novel object recognition (Fig. S2A-B) nor spatial learning and memory in MWM (Fig. S2D-G) was detected by PTH1-34 treatments in 5XFAD male mice, although PTH1-34 had a positive effect on the spontaneous alternation during Y maze tests (Fig. S2C). In aggregates, these intriguing results suggest that PTH1-34 improves cognitive function in 5XFAD mice, which is currently manifested primarily in female mice.
Fig. 2
PTH1-34 attenuation of cognition decline and memory loss in 5XFAD female mice. A Illustration of PTH1-34 or vehicle intermittent treatment in WT/5XFAD female mice and behavioral testing schedule. All mice were tested for behavior from the age of 5 ~ MO old and treated with PTH1-34/Veh continuously during the testing procedure. The results shown in this figure were for female mice. B, C Novel object recognition (NOR): Representative tracing images (B), and the quantification of discrimination index ([novel object explore time − old object explore time]/[total explore time]) of NOR (C) were shown. D Y maze: Quantifications of the total arm entries and spontaneous alternation. E–H Morris water maze (MWM): The latencies to reach the hidden platform during the training period (E), the representative tracing images (F), quantification of time spent in the target quadrant (G), and target zone crossovers (H) on the testing day were showed. All quantification data were presented as mean ± SD (n = 6–8 female mice per group). *P < 0.05, **P < 0.01, two-way ANOVA with two-stage step-up method of Benjamini, Krieger, and Yekutieli multiple comparisons test was used
PTH1-34 reduction of soluble Aβ level and in-soluble Aβ deposition in 5XFAD brainIt is known that the β-amyloid accumulation and deposition in 5XFAD mice is a crucial pathology for AD development [29]. To investigate the effect of PTH1-34 treatments on AD-relevant brain pathology, 5XFAD mice (both male and female) were administered with hPTH1–34 or vehicle (Veh) as illustrated in Fig. 3A. We first measured both human Aβ40 and Aβ42 levels in soluble fraction from brain homogenates (cortex and hippocampus) of 5XFAD female mice in response to PTH1-34 treatments. ELISA analyses showed significant reductions in human Aβ40 and Aβ42 levels in homogenates of both cortex and hippocampus from 5XFAD mice treated with PTH1-34, as compared with those of Veh treatments (Fig. 3B–C), suggesting PTH1-34’s inhibitory effect on Aβ accumulation. To determine whether the insoluble human Aβ and mouse Aβ levels are altered in 5XFAD mice by PTH1-34 treatments, we then evaluated Aβ plaques or deposition in both groups of brain sections by Thio-S staining. In line with the view of PTH1-34 inhibition of Aβ accumulation, PTH1-34 treatments decreased Thio-S positive ( +) Aβ plaques in cerebral cortex and hippocampus of 5XFAD female mice; both plaque density in each layer of cortex or sub-regions of hippocampus and plaque size showed significant reductions, as compared with those of Veh-treated 5XFAD female mice (Fig. 3D, F, I, L). Notice that this PTH1-34 effect was detected in not only female but also in male 5XFAD mice (Fig. 3D–E). Plaque density (mainly in the fifth and sixth layers of the cortex and in the DG region of the hippocampus) (Fig. 3G, J) and plaque size (Fig. 3L) were all decreased in PTH1-34-treated 5XFAD male mice, as compared with Veh-treated male mice, indicating PTH1-34’s gender independence in this pathology. Remarkably, in the absence of the PTH1-34, the levels of amyloid deposition (both plaque density and plaque size) in the cortex and hippocampus of female 5XFAD mice were significantly higher than those in male 5XFAD mice (Fig. 3H, K, L). Interestingly, PTH1-34 reduction of the levels of amyloid deposition appeared to be more dramatically in female than those of male 5XFAD mice, with ~ 46% and ~ 59% reductions in plaque density in female 5XFAD cortex and hippocampus, respectively, while ~ 32% and ~ 50% reductions in male 5XFAD cortex and hippocampus, respectively, and ~ 34% and ~ 56% reductions in plaque size in female 5XFAD cortex and hippocampus, respectively, while ~ 28% and ~ 23% reductions in male 5XFAD cortex and hippocampus respectively (Fig. 3H, K, L). Significant effects of gender and PTH1-34 treatment were showed in both plaque density and plaque size (P < 0.05), no matter in the cortex or hippocampus (Supplemental file 1 Table 3). Significant interactions between PTH1-34 treatment x gender were observed in both plaque density (cortex: F = 11.17, P = 0.0027; hippocampus: F = 5.021, P = 0.0346) and plaque size (only hippocampus: F = 16.2, P = 0.0005) (Supplemental file 1 Table 3), indicating a sex difference in response to PTH1-34 treatment. Together, these results suggest that intermittently PTH1-34 treatments attenuate Aβ pathology in 5XFAD brain, and this effect is more obvious in female mice.
Fig. 3
PTH1-34 reduction of soluble Aβ level and in-soluble Aβ deposition in 5XFAD brain. A Illustration of PTH1-34 intermittent treatment in 5XFAD mice. Mice were sacrificed at 5 ~ MO old to detect Aβ level and accumulation. B, C ELISA analyses of human Aβ40(B) and Aβ42(C) levels in the soluble fraction of brain homogenates including cortex and hippocampus (200 μg total protein) from female 5XFAD mice with PTH1-34 or Veh treatment (n = 4 mice per group). D, E Representative images of ThioS staining for Aβ plaque depositions analysis in the cortex and hippocampus of 5XFAD-Veh (control) and 5XFAD-PTH1-34 mice. Representative images of female mice were shown in D and representative images of male mice were shown in E. F, G Quantification of plaque density (the amount of plaque deposition in each sub-region) in the cortex of 5XFAD female and male mice. H Quantification of the total plaque density in the cortex of female and male 5XFAD mice. I, J Quantification of plaque density in subregions of hippocampus in 5XFAD female and male mice. K Quantification of the total plaque density in the hippocampus of female and male 5XFAD mice. L Quantification of average plaque size in 5XFAD female and male mice. n = 8 per group for female mice and n = 6 per group for male mice in F–L. Scale bars as indicated in each panel. All data were presented as mean ± SD. *P < 0.05, **P < 0.01, ***P < 0.001, two-way ANOVA with Sidak’s multiple comparisons test was used
PTH1-34 reduction of plague-associated dystrophic neurites, but not plaque-associated microglial cells in 5XFAD miceTo understand how PTH1-34 suppresses Aβ accumulation and deposition in the 5XFAD brain, we further examined Aβ plaque-associated pathology in 5XFAD female mice (~ 6 MO) treated with PTH1-34 or vehicle. Aβ plaques are surrounded by dystrophic neurites, a feature of neurodegenerative pathology, and activated glial cells, including IBA1+ microglia and GFAP+ astrocytes [44]. Co-immunostaining analysis of ThioS with ATG9A, a pre-autophagosome protein that accumulates in the dystrophic neurites and is commonly used as a marker for dystrophic neurites [44], showed smaller and less dense ATG9A+ dystrophic neurites in PTH1-34 treated 5XFAD brain (both cortex and hippocampus) than those of Veh-treated 5XFAD mice (Fig. S3A-B), indicating a reduction of dystrophic neurite formation by PTH1-34, in line with the view of PTH1-34 decrease of Aβ plaques.
We then performed a co-immunostaining analysis of ThioS with IBA1, a marker for microglial cells. Although the overall IBA1 fluorescence intensity was lower in PTH1-34 treated 5XFAD cortex and hippocampus than that of Veh controls (Fig. S4A-B), the ThioS+ Aβ plaque associated IBA1 fluorescence was unchanged by PTH1-34 treatments (Fig. S4D-E). Additionally, the IBA1 + cell density and fluorescence intensity in regions without Aβ plaques were also comparable in PTH1-34 treated 5XFAD mice to those of Veh treatments (Fig. S4C). These results suggest that the overall reduction in IBA1 fluorescence intensity may be due to the reduced number of Aβ plaques.
Notice that the Aβ plaque-associated microglial cells are also called DAM (Degeneration Associated Microglia), because of their unique distribution pattern and expression of molecular features (e.g., higher expressions of genes such as LPL, but lower levels of TMEM119 and CX3CR1, than those of Aβ-un-associated microglia or resting microglia) [33, 45]. As DAMs are implicated in promoting Aβ clearance [33, 46], we further examined whether DAMs are affected by PTH1-34 treatments. Co-immunostaining analysis of IBA1 and LPL (a marker for DAM) with ThioS showed little to no difference of Aβ associated LPL+IBA1+ microglial cells in the brain between PTH1-34 and Veh-treated 5XFAD mice (Fig. S4F-G), suggesting a little effect on DAM formation by PTH1-34. These results demonstrate decreased amyloid plaques accompanied by decreased neural toxicity in PTH1-34 treated 5XFAD mice, suggesting PTH1-34’s function in preventing both Aβ accumulation and dystrophic neurites formation.
PTH1-34 diminishment of GFAP+ reactive astrocytes and brain inflammation in 5XFAD miceTo further identify the effect of PTH1-34 on Aβ plaque-associated pathology, we examined GFAP+ astrocytes in female 5XFAD brains with PTH1-34 or Veh treatments (~ 6 MO). As shown in Fig. 4A–B, co-immunostaining analysis of GFAP (a marker for reactive astrocytes in cortex) with ThioS showed a similar overall response as those of IBA1+ microglial cells to PTH1-34 treatments. The increase in GFAP + reactive astrocytes was found in areas of Aβ deposition in 5XFAD-Veh mice, including each layer of the cortex, as well as the DG region of the hippocampus. The GFAP+ fluorescence intensity in layers of cortex (except layer II/III) and DG region of hippocampus were lower in PTH1-34 treated 5XFAD mice than that of Veh controls (Fig. 4A–B), in correlation with the reduced Aβ plaques by PTH1-34 treatments. However, in contrast to unchanged Aβ-plaque associated IBA1/LPL fluorescence (Fig. S4D-G), the plaque-associated GFAP fluorescence appeared to have diminished by PTH1-34 treatments (Fig. 4C–D). There were significant numbers of ThioS + plaques with little or no GFAP+ astrocytes surrounding the brain (cortex and hippocampus) of PTH1-34 treated 5XFAD mice (Fig. 4C). As the overall fluorescence intensity of IBA1 and GFAP decreased in PTH1-34 treated 5XFAD brain, we further tested this view by Western blot analysis. Certainly, lower IBA1 and GFAP protein levels were detected in PTH1-34 treated 5XFAD brain homogenates (cortex and hippocampus) (Fig. 4E–F), supporting the view for an overall reduction in glial reactivation by PTH1-34. Notice that this PTH1-34 inhibition of glial cell activation was detectable in both female and male 5XFAD mice (Figs. 4, S5); but PTH1-34 had little effect in these glial cells in wild-type (WT) mice (Fig. S6). These results thus suggest PTH1-34’s inhibitory effect on both astrocyte and microglial activation in 5XFAD, but not WT, mice in a sex-independent manner.
Fig. 4
PTH1-34 diminishment of GFAP + reactive astrocytes. A Representative overall images of co-immunostaining with ThioS (green) and GFAP (red) of cortex and hippocampus from 6 ~ MO 5XFAD-Veh and 5XFAD-PTH1-34 female mice. B Quantification of relative GFAP fluorescence intensity in each layer of cortex and subregional of hippocampus. C Representative images and high-magnification images in Aβ deposition regions of co-immunostaining with ThioS (green) and GFAP (red) of cortex and hippocampus from 6 ~ MO 5XFAD-Veh and 5XFAD-PTH1-34 female mice. D Quantification of Abeta-associated GFAP fluorescence intensity. The Abeta-associated GFAP fluorescence intensity was defined by the intensity of GFAP-positive astrocytes in a plaque-centered circle within 50 μm in diameter (marked by dashed white circles). n = 8 mice per group. E Representative Western blots using antibodies against IBA1 and GFAP in homogenates of cortex and hippocampus from 6 ~ MO WT and 5XFAD female mice with PTH1-34 or Veh treatment. GAPDH was used as a loading control. F Quantification of relative protein level in E (n = 5 mice per group). All quantification data were presented as mean ± SD. Scale bars were indicated in each panel. *P < 0.05, **P < 0.01, ***P < 0.001, two-way ANOVA with Sidak’s multiple comparisons test was used
We then asked whether PTH1-34 treatments affected the expressions of proinflammatory cytokines and chemokines in 5XFAD brain. RT-qPCR analyzed ~ 10 genes’ expressions, which are either well-recognized proinflammatory cytokines (e.g., IL1β, IL6, TNFα, TGFβ) or cytokines involved in bone remodeling (e.g., RANKL, RANK, OPG, and GM-CSF), in 6 ~ MO old WT/5XFAD female brain (cortex and hippocampus) with PTH1-34 or vehicle treatments (Fig. 5A, D). While most of these factors were upregulated in 5XFAD brain (cortex and hippocampus), as compared with those of same-aged WT mice, these upregulated inflammatory cytokines or chemokines were largely brought down to nearly normal levels by PTH1-34 (Fig. 5A–B, D–E). Notice that while the majority of factors were downregulated by PTH1-34 treatments in both 5XFAD cortex and hippocampus, a few cytokines, largely factors involved in bone remodeling (e.g., GM-CSF, RANKL, and OPG in the cortex) occurred unaffected by PTH 1–34 (Fig. 5C, F). This is in contrast to the effect of PTH in osteoblastic bone cells, where PTH is known to regulate RANKL and OPG expression [47, 48]. Taken together, these results suggest that PTH1-34 treatments reduce glial cell activation and consequent brain inflammation.
Fig. 5
PTH1-34 diminishment of brain inflammation in 5XFAD mice. A Real-time PCR (RT-PCR) analysis of indicated gene expressions in the cortex of 6 ~ MO WT and 5XFAD female mice with PTH1-34 or Veh treatment. The level of GAPDH was normalized to 1, n = 3 per group. B Total quantification of relative gene expression levels in A. The level of WT group was normalized to 1, n = 10 gene per group. C The summary of altered genes in the 5XFAD-PTH1-34 treatment group compared with the 5XFAD-Veh group in cortex. D RT-PCR analysis of indicated gene expressions in the hippocampus of 6 ~ MO female mice, n = 3 per group. E Total quantification of relative gene expression levels in D. F The summary of altered genes in the 5XFAD-PTH1-34 treatment group compared with the 5XFAD-Veh group in hippocampus. All quantification data were presented as mean ± SD. *P < 0.05, **P < 0.01, ***P < 0.001, one-way ANOVA with Tukey’s multiple-comparison test was used
PTH1-34 reduction of serum inflammatory cytokinesTo understand how PTH1-34 treatments affect brain inflammation in 5XFAD mice, we investigated PTH1-34’s effect on systemic inflammation, since PTH is a hormone with systemic effects, particularly in bone cells [17, 18]. Using multiplexed antibody-based arrays to screen for altered serum/plasma proteins in 5XFAD and WT female mice (6 ~ MO) with PTH1-34 or vehicle treatments, we found that among 40 factors, few (7/40) were elevated in serum samples from 5XFAD mice, as compared with that of WT mice (Fig. 6A–C), suggesting a weak systemic inflammation in 5XFAD mice. Notice that most of these elevated serum factors (e.g., IL-1rα, IL-2, IL-3, IL-4, IL-10, CXCL9, CCL3) in 5XFAD mice were all reduced by PTH1-34 treatments (Fig. 6C–D). Additionally, PTH1-34 reduced some serum inflammation-associated factors such as IL1β, TNFα, TREM-1, CCL11, and CCL5, which were comparable between WT and 5XFAD mice, and restored serum factor CCL4, which was decreased in 5XFAD mice (Fig. 6C–D). We compared the PTH1-34 downregulated cytokines in serum of 5XFAD mice with those in brain of 5XFAD, as shown in Fig. 6E. The results showed that 4 factors, IL-10, IL-1β, TNFα, and CCL5, were reduced by PTH1-34 treatments not only in the serum but also in the brain of 5XFAD, suggesting a certain relationship between systemic inflammation and brain inflammation (Fig. 6E). We further compared these changes in serum factors between 5XFAD and Tg2576 mice, another AD animal model expressing Swedish mutant APP ubiquitously [30, 49]. In contrast to that of 5XFAD mice, Tg2576 mice exhibited elevations in many (15/40) of these serum factors (Fig. 6F). These results demonstrate the systemic effect of PTH1-34, although systemic inflammation in 5XFAD mice seems to be weaker than in Tg2576 mice.
Fig. 6
PTH1-34 reduction of serum inflammatory cytokines. A Representative images of serum L-Series label-multiplex antibody arrays of 6 ~ MO WT and 5XFAD female mice with PTH1-34 or Veh treatment. Proteins with changes are marked by dashed blue circles. B Total quantification analyses of the data in A. WT-Veh group were normalized to 1, n = 36 proteins per group. C Quantification of relative serum protein levels in A. The data showed those proteins that were significantly changed and arranged into different groups according to their characteristics. The data were presented as mean ± SD (n = 4 mice per group), *P < 0.05, **P < 0.01, ***P < 0.001, one-way ANOVA with Tukey’s multiple-comparison test. D Heat map of serum protein. n = 4, significant difference was set at P < 0.05. E Comparison of the PTH1-34 downregulated cytokines in serum of 5XFAD mice (downregulated cytokines in serum of PTH1-34 treated 5XFAD mice over 5XFAD control mice) to those detected in the cortex or hippocampus of 5XFAD-PTH1-34 mice. F Comparison between the 5XFAD with Tg2576 antibody array of secreted proteins in serum
PTH1-34 association with astrocytes in the brainGiven the significant effect of PTH1-34 in 5XFAD brain pathology, we wondered whether the injected PTH1-34 functioned directly in 5XFAD brain. To address this question, Biotin-conjugated PTH1-34 was administered into WT and 5XFAD female mice (~ 2.5 MO) by caudal vein injection, and mice were sacrificed 30 min after injection, as illustrated in Fig. 7A. Immunostaining analysis with biotin antibody showed more biotin signals in the brain of 5XFAD mice than in WT mice (Fig. 7B–C), implicating a BBB (blood–brain barrier) leakage in 5XFAD brain [3, 50] that causes more Biotin-PTH1-34 to enter the brain. Further co-immunostaining analysis showed that most Biotin-PTH1-34 signals were associated with GFAP+ astrocytes (~ 60%); ~ 10–17% of Biotin-PTH1-34 signals were in association with IBA1+microglia, SLC16A1+ blood vessels, and MAP2+ neurons, respectively (Fig. 7D). These results thus suggest that injected biotin-PTH1-34 can enter the brain through BBBs of 5XFAD mice and bind to astrocytes in large quantities.
Fig. 7
More PTH1-34 association with astrocytes in 5XFAD brain than that of WT control. A Schematic diagram of experimental design. 2.5 ~ MO WT and 5XFAD female mice were administered PTH1-34-Biotin (100 μg/100 μl) or vehicle (phosphate buffer, 100 μl) by tail-intravenous injection to detect PTH1-34 diffusion in vivo. Mice were sacrificed after 30 min for brain isolation and tissue sectioning, and immunofluorescence staining was used to analyze the distribution of PTH1-34-Biotin. B Representative images and high-magnification images of Biotin (green) co-immunostaining with GFAP, IBA1, SLC16A1, and MAP2 (red) respectively from 5XFAD + Veh, 5XFAD + PTH1-34-Biotin and WT + PTH1-34-Biotin female mice. All images were obtained from the cortex regions. Scale bars were indicated in the panel. C Quantification of relative Biotin fluorescence intensity in these three groups (mean ± SD; n = 8 mice per group). **P < 0.01, ***P < 0.001, one-way ANOVA with Kruskal–Wallis multiple-comparison test. D Quantification of PTH1-34-Biotin distribution in 5XFAD brain cells from B. The distribution of PTH1-34 in various cells was shown as a percentage, with a total proportion of 100%, derived from the mean value of data collected from all mice in the 5XFAD + PTH1-34-Biotin group
The abundant association of Biotin-PTH1-34 with astrocytes leads us to ask whether this is due to the abundant expression of PTH receptor (PTH1R) in astrocytes. Indeed, scRNA-seq database showed selective PTH1R expression in astrocytes and endothelial cells in the brain. We further verified this view by RNA scope analysis (a higher resolution of in situ analysis), which showed co-distribution of PTH1R’s mRNAs largely with Glast1-mRNA+ astrocytes (Fig. S7C-D). Moreover, western blot analysis of lysates from primary cultured astrocytes demonstrated expression of PTH1R (Fig. S7A-B). Interestingly, the PTH1R protein level appeared to be slightly higher in astrocytes from 5XFAD mice than that of WT astrocytes (Fig. S7B-D). Together, these results support the view that injected PTH1-34 can travel to the brain and bind to its receptor in astrocytes in 5XFAD mice.
PTH1-34 suppression of proinflammatory cytokines’ expression in 5XFAD astrocytesTo verify the view that PTH1-34 functions directly on astrocytes, we examined whether PTH1-34 could induce signaling and function in primary cultured astrocytes from WT and 5XFAD female pups (Fig. S8A). Interestingly, among PTH1-34 induced signaling pathways identified in osteoblasts (e.g., cAMP-driven phospho-CREB, phospho-AKT, and phospho-ERK1/2), PTH1-34 increases p-CREB, but has little effect on either p-AKT or p-ERK (Fig. S8B-E), suggesting that PTH1-34 induction of cAMP to p-CREB may be a key signaling pathway in astrocytes. Notice that a trend of increasing, but not significant, p-CREB was detected in 5XFAD astrocytes, as compared to WT astrocytes (Fig. S8C), which might be due to a slight increase in the expression of PTH1R in 5XFAD astrocytes. These results thus support the view that PTH1-34 functions directly on astrocytes.
We further tested this view by examining PTH1-34’s effect on the expression of cytokines and chemokines in astrocytes. Cultured astrocytes from WT and 5XFAD female pups were treated with vehicle or PTH1-34 for 24 h, and then subjected to RT-qPCR analysis, as illustrated in Fig. S9A. Increased levels of cytokines or chemokines (e.g., TNF-α, TGF-β1, CCL5, GM-CSF, and RANK) were detected in 5XFAD astrocytes, as compared with those in WT astrocytes (Fig. S9B-D), consistent with multiple literature reports of a higher basal inflammatory state in 5XFAD cells [51, 52]. Nearly all these factors (except GM-CSF) were reduced by PTH1-34 treatments (Fig. S9B, E). We compared these changes with PTH1-34 downregulated genes in the 5XFAD brain (cortex and hippocampus). As illustrated in Fig. S
留言 (0)