Neurodegeneration
Given the critical roles astrocytes play in ensuring proper CNS function, including maintaining BBB integrity, phenotypic alterations can significantly contribute to neurological impairments. A1-like astrocyte populations have been shown to arise with normal aging and are further increased in neuroinflammatory contexts; accordingly, reactive astrocyte abundance is associated with several infectious and neurodegenerative disorders [21, 75]. The first research into the newly-characterized A1-like astrocytes explored their role in the more common neurodegenerative disorders, including Alzheimer’s disease (AD), Parkinson’s disease (PD), Huntington’s disease (HD), amyotrophic lateral sclerosis (ALS), and multiple sclerosis (MS) [21, 75, 76]. Indeed, the initial work by Shane Liddelow and Ben Barres that described A1/A2 polarization in 2017 confirmed the translational relevance of their findings using human post-mortem brain tissue samples from relevant regions of neurodegeneration in these disorders [76]. In situ hybridization and co-immunofluorescent staining methods to analyze AD, PD, HD, ALS, and MS samples revealed a high degree of co-localization of the astrocyte markers S100β and GFAP and the A1 marker C3 in disease-relevant brain regions, indicating that the astrocytes present had adopted a pro-inflammatory phenotype [76]. In healthy tissue, the proportion of astrocytes that could be classified as A1-like astrocytes is low, typically ranging from 10 to 20% of the local astrocyte population, depending on the brain region [76]. A1-like astrocytes are not so rare in neurodegenerative disorders; indeed, up to 60% of astrocytes in regions of the brain affected by neurodegeneration in these chronic disorders are of an A1-like phenotype, effectively taking over the astrocyte population [76]. When they examined the presence of reactive astrocytes within demyelinating lesions across different MS stages, they found that A1-like polarization correlated with disease progression [76]. Neurotoxic A1-like astrocytes have altered, cytotoxic functions, secrete pro-inflammatory signals, and promote neuroinflammation, contributing to neurodegeneration [21, 75, 76].
The primary goal of neuroinflammation is to defend against pathogenic infiltration into the CNS; however, neuroinflammation is also a hallmark of neurodegenerative diseases. Typically, there is a low-level, chronic expression of pro-inflammatory signals throughout the course of neurodegeneration, and this long-term inflammatory state may contribute to disease progression [66]. Neurodegenerative diseases like AD and PD are characterized by toxic aggregates of pathological proteins, such as tau and amyloid-β in AD and α-synuclein in PD. Astrocytes and microglia can function cooperatively to process toxic protein deposits [105]. Specifically, co-cultured human iPSC-derived astrocytes and microglia exposed to amyloid-β and α-synuclein engulf the protein aggregates, then transfer the internalized protein between each other through tunneling nanotubes [105]. The use of human iPSC-derived astrocytes is significant, as rodent-derived iPSCs do not necessarily share the same gene expression profiles [9]. This cell-to-cell transfer of pathological protein is consistent with the organized manner these cells clear debris [105]. Larger aggregations tend to be degraded by microglia, while astrocytes, which can act as antigen-presenting cells, are more likely to process the smaller or monomeric proteins [105]. However, this mechanism is insufficient to prevent the progression of diseases like AD and PD, and the clearing capacity of these cells becomes overwhelmed. Aggregations of pathological proteins further damage nearby cells, prompting the release of pro-inflammatory cytokines, which serve as signals that induce microglia to adopt an inflammatory phenotype [66]. These activated microglia go on to mount an inflammatory response, which includes astrocyte polarization via secretion of TNF-α, IL-1α, and C1q [76]. Increased levels of inflammatory markers, like TNF-α or IL-1β, have been detected in the CSF of individuals with neurodegenerative disorders, and these levels correlate to degree of cognitive impairment, emphasizing the importance of neuroinflammation in neurodegenerative disease progression [16, 66].
Alzheimer’s disease (AD)
AD is a progressive neurodegenerative disorder that currently effects just under 50 million people. With advances in medicine prolonging the average human lifespan, the incidence of AD is expected to rise to 115 million in the next 30 years [36, 58]. As the leading cause of dementia, AD is one of the most commonly-studied neurodegenerative disorders. The primary pathological features found in AD include extracellular deposits of abnormal proteins termed amyloid-β (Aβ) that form destructive amyloid plaques in the brain tissue, and intraneuronal aggregates of misfolded and hyperphosphorylated Tau protein that form toxic neurofibrillary tangles. The accumulation of these pathogenic proteins over time is associated with synapse loss, neuronal death, and chronic neuroinflammation [8, 36, 58]. The initial pathology appears to be localized to the entorhinal cortex and hippocampus, structures associated with memory formation [8, 13]. Difficulties with memory and executive functioning expand to include other cognitive dysfunctions, including aphasia. People living with moderate AD often lose the ability to complete tasks with multiple steps, learn new information, recognize faces, and consolidate long-term memories [7]. Neuropsychological symptoms also arise, frequently resulting in disorientation, hallucinations, and paranoia [7]. Increasing damage and substantial cortical atrophy results in the eventual loss of communication abilities, as well as the loss of physical functions like walking and even swallowing. As a progressive neurogenerative disorder, the complications of AD ultimately prove fatal.
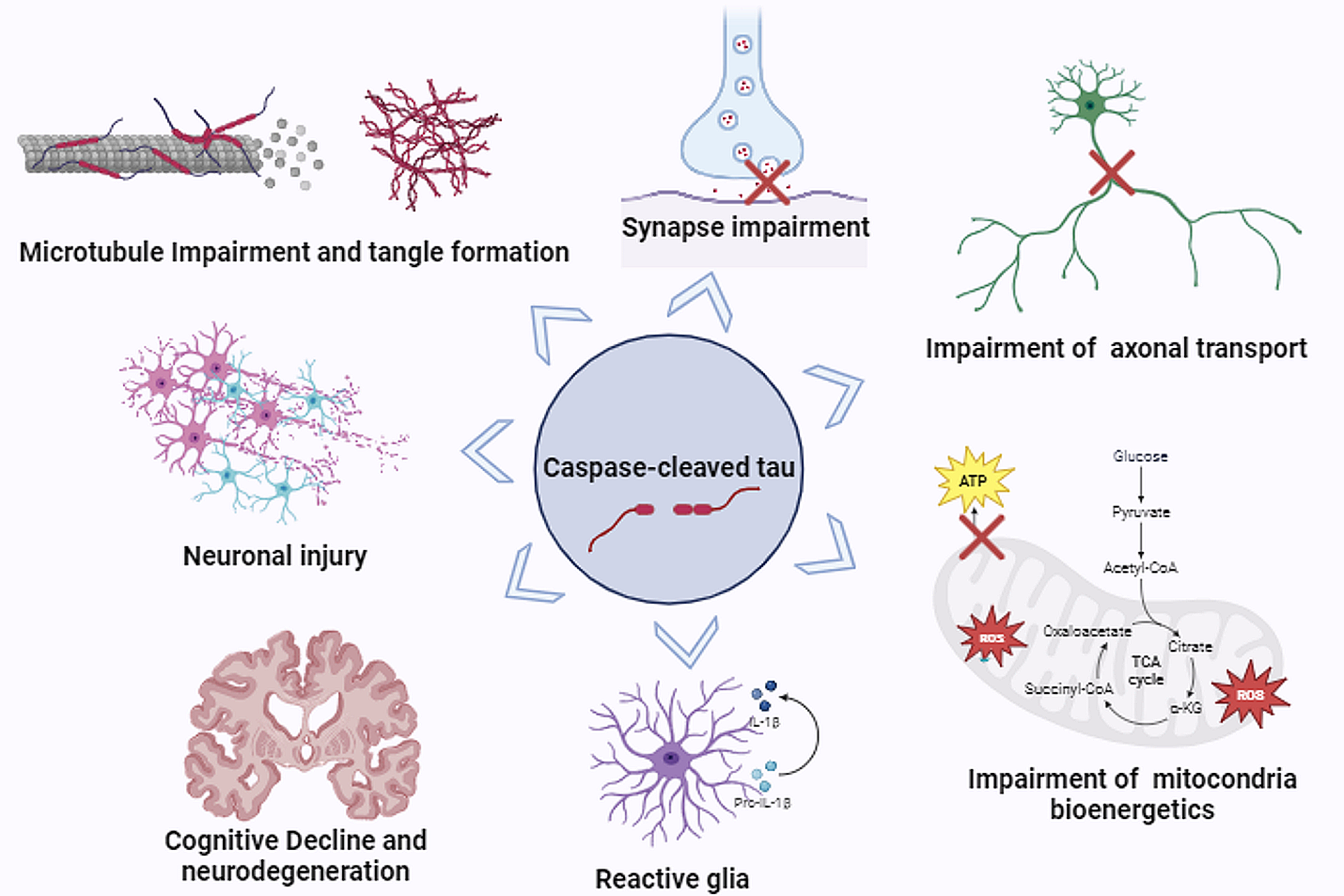
Prior to the introduction of the A1/A2 classification paradigm, non-proliferative reactivation had been observed in astrocytes in AD. Analysis of temporal cortex samples from symptomatic AD patients revealed increased levels of reactivity markers, specifically major histocompatibility complex II (MHCII) for microglia and GFAP for astrocytes [108]. However, the levels of constitutively-expressed markers, ionized calcium-binding adaptor molecule 1 (IBA1) in microglia and aldehyde dehydrogenase 1 L1 (ALDH1L1) in astrocytes, did not increase indicating the reactivation response was phenotypic in nature rather than solely proliferative [108]. Furthermore, cortical NF-κB and C3 expression levels are significantly higher in the context of AD, both in human patients and transgenic mouse models [74]. Both in vitro and in vivo, C3 upregulation is correlated with Aβ accumulation, a hallmark of AD. Further investigation indicates that Aβ exposure activates the NF-κB pathway in astrocytes, with one of the transcriptional targets being C3 [74] (Fig. 1e). Astrocytes release the upregulated C3, which binds to the C3a receptor (C3aR) on neurons. Increased C3-C3aR signaling dysregulated intraneuronal calcium homeostasis, ultimately disrupting dendrite morphology, reducing synaptic density, and impairing excitatory synaptic transmission in the context of AD [74] (Fig. 2). These findings are consistent with a pathological role for some reactive astrocytes, which display NF-κB pathway activation and increased C3 expression [76, 135].
AD has quickly become a commonly used model to study A1-like astrocytes. Since their characterization, a variety of mechanisms by which A1-like astrocytes contribute to AD pathology have been described. The link between A1-like astrocytes and AD appears to begin at the genetic level. Among the 40 or so loci that may possess alleles associated with an increased risk of developing AD is the SPI1 gene which encodes PU.1, a transcription factor that regulates myeloid cell development and differentiation [101]. For example, in response to injury or insult in the brain, PU.1 expression may be upregulated, allowing it to facilitate transcriptional changes in microglia and resident macrophages [101]. In AD there are abnormally high levels of PU.1 expression, and several of the genes it controls are also associated with AD development [101]. One consequence of this altered microglia response is altered astrocyte activation. When microglia with AD-like PU.1 overexpression are exposed to LPS, they are more efficient at inducing polarization in astrocytes, as evidenced by increased A1-specific transcriptional markers when exposed to LPS-stimulated MCM from PU.1 overexpressing microglia, compared to wildtype microglia [101].
Interestingly, it appears that microglia are not the only mechanism by which A1-like astrocytes are activated in the context of AD. One model of AD uses trimethyltin (TMT) intoxication, in which the resulting hippocampal degradation gives rise to molecular and behavioral signatures of AD in rodents [28]. When applied to primary cortical astrocytes in vitro, TMT disrupts intracellular Ca2+ homeostasis by causing an influx of Ca2+ through L-type voltage-gated ion channels [28]. Subsequently, the mitochondrial membrane becomes depolarized, levels of ROS and NOS increase, and A1-associated pro-inflammatory pathways are activated (Fig. 1f). The C3 + A1 phenotypic switch that occurred in this model of AD was also accompanied by morphological changes including retraction of cellular processes [28]. Other in vitro and in vivo models have also been useful in cataloging the roles of A1-like astrocytes in AD. Aβ has been shown to act in vitro similarly to a pro-inflammatory cytokine and even at low concentrations can be substituted for C1q in the TNF-α and IL-1α cocktail that induces polarization to a pro-inflammatory phenotype [70] (Fig. 1e). Further research has demonstrated that Aβ-treated microglia-conditioned media not only promoted A1-like polarization, but also upregulate astrocytic glycolysis [145] (Fig. 2). Interestingly, changes in glucose metabolism are characteristic of early AD, and the role of glycolysis in astrocyte activation in both AD and other pathologies requires continued investigation [145]. Overall, these experiments suggest that at least one of the pathological features underlying AD, Aβ aggregation, can itself activate astrocytes and promote a phenotypic polarization, potentially explaining the high proportion of A1-like astrocytes in post-mortem human brain tissue of individuals with AD [76] (Fig. 1e). As AD most often appears in elderly populations, this issue is likely compounded by the natural accumulation of A1-like astrocytes as the brain ages [21].
The neurotoxic and pro-inflammatory phenotype of A1-like astrocytes likely contributes to the progression of AD pathology. Astrocyte-derived exosomes (ADEs) obtained from the blood plasma of individuals living with AD were found to contain high levels of a variety of complement proteins, including C3, and decreased levels of complement regulatory proteins, compared to ADEs from individuals without AD [34] (Fig. 2). This suggests the astrocytes adopted an A1-like phenotype. When analysis was applied to a longitudinal investigation, samples from patients in early pre-clinical stages had complement levels comparable to the control group, and as the disease progressed and symptoms developed, complement levels in ADEs increased [34]. A1-like astrocytes may also exert pathological effects in the context of AD by contributing to signaling dysfunction [8]. In one study that looked at hippocampal and entorhinal cortical samples from a rat model of AD and non-AD control, C3 + reactive astrocytes significantly upregulated expression of serine racemase (SR) [8]. SR catalyzes the synthesis of D-serine, a co-agonist for the neuronal NMDA receptor (NMDAR), and its overexpression by A1-like astrocytes was associated with increased NMDAR activation and associated excitotoxicity [8]. Consistent results were observed in human post-mortem brain tissue samples obtained from previously diagnosed AD patients, indicating reactive astrocyte-mediated excitotoxicity may be another mechanism by which neurotoxic astrocytes contribute to neurodegenerative disease [8].
A major challenge to treating AD is the lack of any easily-detectable biomarkers in living patients. AD is diagnosed via a battery of neuropsychological assessments and can only be definitively confirmed at autopsy where Aβ and pTau aggregates can be identified in brain tissue [36]. Neuroimaging techniques, including PET and MRI, in combination with CSF analysis can provide a more thorough examination, but there are major financial and access barriers to these invasive methods [36]. Symptoms do not appear until amyloid plaques and neurofibrillary tangles have elicited enough irreversible tissue damage, rendering early treatment impossible. The recent discovery of reactive astrocytic phenotypes may ultimately provide a method of pre-symptomatic identification of AD, potentially by analyzing an unlikely location: the retina. Aβ and pTau aggregates were detected via immunostaining of post-mortem retinal slices from patients with AD, as well as associated increased levels of IL-1β, which co-localized with microglial Iba1, and C3 colocalized with astrocytic GFAP [36]. This could pave the way for future diagnostic applications of non-invasive retinal scans.
Supporting the hypothesis that different reactive astrocyte polarization dynamics may in some way complement each other, one study identified the presence of both phenotypes in the context of AD [58]. Astrocyte populations were assessed using a cohort of post-mortem human samples from diagnosed and neuropathologically-confirmed cases of AD at similar progression stages and non-AD controls [58]. Immunostaining with C3 was used to indicate A1-like neuroinflammatory astrocytes, while S100A10 was used to indicate neuroprotective A2-like astrocytes [58]. The number of A1-like cells was higher than A2-like cells in both control and AD, suggesting that the two phenotypes are not evenly balanced [58]. In AD, the density of A1-like cells was significantly increased in certain tissues, specifically in the upper cerebral cortex. Interestingly, the density of A2-like cells was also significantly increased in AD, in the upper and lower cortex as well as the white matter [58]. The researchers suggest that a neuroinflammatory response occurs in AD, in which quiescent astrocytes become A1-polarized and pre-existing age-associated A1-like astrocytes become more active [58]. Reactivity associated with pathogenic protein accumulation may explain why reactive astrocyte growth is more localized rather than widespread. Subsequent neuroinflammation and damage could elicit a generalized A2-like polarization signal across a larger tissue area to provoke a neuroprotective response [58]. More research will be needed to investigate this potential mechanism and tease out the interplay among neuroinflammatory A1-like and neuroprotective A2-like astrocytes.
Parkinson’s disease (PD)
Parkinson’s disease (PD) is the second most common neurodegenerative disorder and affects 2% of all people over the age of 65 [136]. The risk increases if an immediate family member also has PD, suggesting a hereditary link in some cases, while several spontaneous genetic mutations are suspected of being causative in other cases [23, 125]. PD is classified as a degenerative type of parkinsonism, which describes any disorder that causes slowed movement or bradykinesia, rigidity, limb dystonia, tremors, and postural instability [23]. Non-motor neuropsychological symptoms like emotional dysregulation, sleep disturbances, dementia, and psychosis, are often present prior to diagnosis and become more severe as the degeneration progresses [23, 125]. While not directly fatal, complications that arise secondary to the pathological changes in PD can be life-threatening.
The symptoms of PD arise due to loss of dopaminergic neurons in the basal ganglia, particularly in the substantia nigra, and a corresponding dopamine signaling deficiency [125, 136]. A key pathological feature of PD is the presence of cytotoxic Lewy bodies, intracellular inclusions containing accumulated misfolded α‐synuclein [136]. Neuroinflammation is another hallmark of PD. Post-mortem samples display significant levels of inflammatory microglia activation [136]. In addition to secreting pro-inflammatory factors like NO and TNF-α, activated microglia in PD may also promote T cell-facilitated neuronal cell death [4,




留言 (0)