
記住我
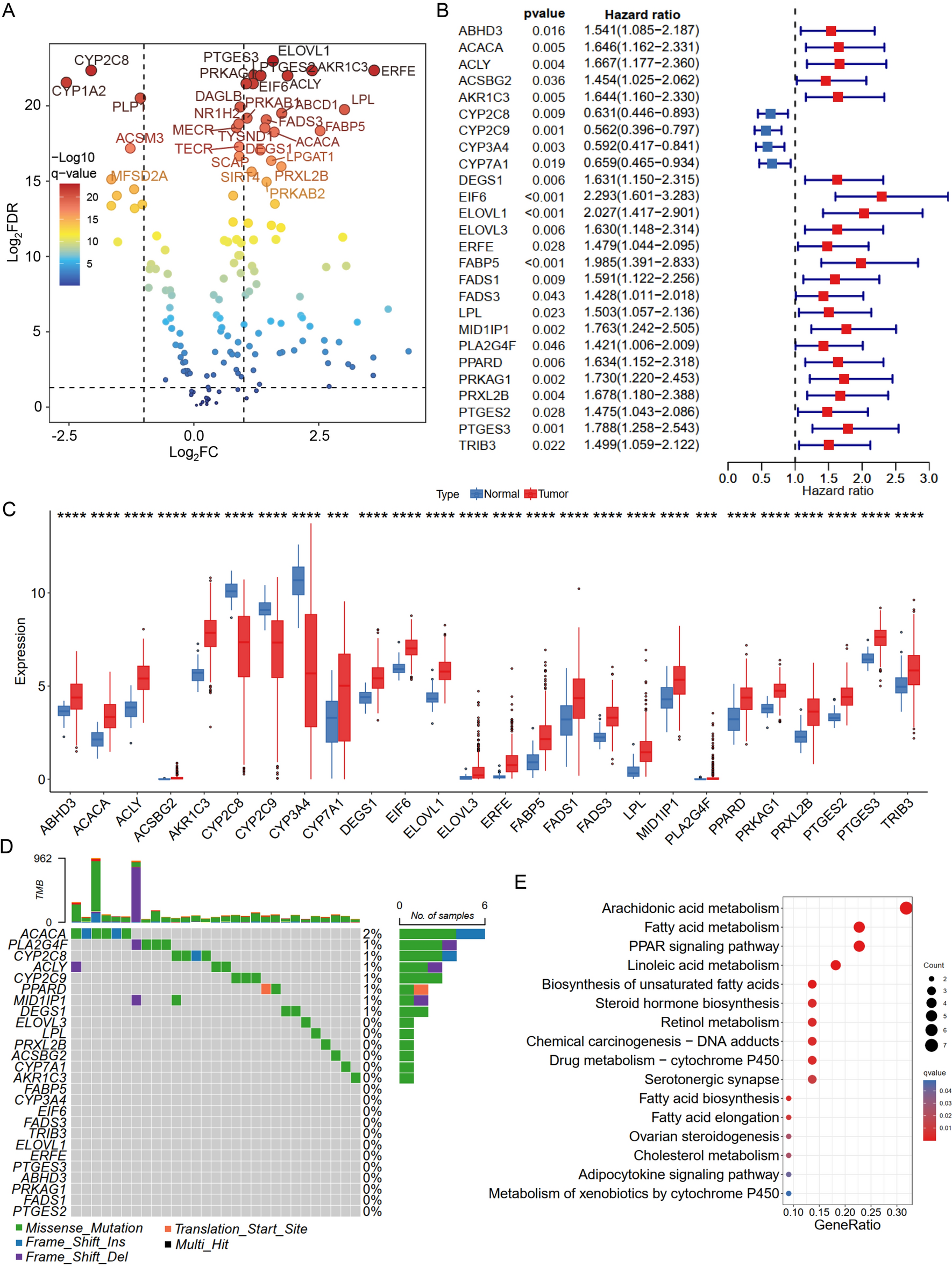
We retrieved two datasets, the sorafenib-resistant dataset for Huh7 cells (GSE94550, [9]) and the Roessler Liver microarray dataset (GSE14520, [10]), and utilized > 2-fold change as the criterion for selection of sorafenib-modulated molecules in sorafenib-resistant Huh7 cells compared to parental cells (GSE94550) and > 1.2-fold as the criterion for choosing oncogenes related to survival in the bottom 25% vs. the top 25% of HCC patients from the Roessler Liver microarray dataset (GSE14520) to intersect the potential candidates for further study. According to the above analysis, the selected genes were validated and filtered more stringently and applied to evaluate highly significant molecules. After retrieving these datasets, 545 upregulated genes (sorafenib resistance vs. control >2-fold) and 609 downregulated genes (sorafenib resistance vs. control < 2-fold) were identified in the sorafenib-resistant hepatoma Huh7 cell dataset (GSE94550). In the Roessler Liver microarray dataset (GSE14520), we used > 1.2-fold as the criterion for choosing oncogenes related to survival in the bottom 25% vs. the top 25% of HCC patients, and only 12 dysregulated genes were identified (Fig. 1A). Finally, we identified 4 potential candidates in the two intersecting microarray datasets, namely, neurotensin (NTS), AGR2, alpha-fetoprotein (AFP) and meprin A, alpha (MEP1A) (Fig. 1A, B). Previously, the AGR2 protein was demonstrated to be a part of the PDI family of ER proteins that mediate the formation of disulfide bonds and catalyze protein folding [32]. Moreover, AGR2 is highly expressed in numerous cancer types, including liver cancer [16]. Through a clinical study, Hrstka et al. showed that AGR2 expression can be used as a marker to predict poor prognosis in breast cancer [15]. Hence, we suggest that AGR2 may be a potential candidate target in sorafenib-treated HCC. Interestingly, AGR2 is the major gene highly correlated with the survival rate and sorafenib resistance in liver cancer; however, the relationship between AGR2 and sorafenib treatment in HCC has not been demonstrated. Therefore, AGR2 was selected for further study to investigate its molecular mechanism associated with sorafenib resistance and its physiological role in HCC.
Fig. 1
Selection of potential candidate genes and analysis of clinical parameter correlations and survival. A Schematic diagram of the analytic protocol for the selection of sorafenib-mediated candidate genes. The 4 selected candidate genes were consistently observed in these two datasets. B The gene descriptions and fold changes in the GSE14520 and GSE94550 datasets are shown. C–D, J–K KM survival curves (OS and RFS) of 2 groups of patients with HCC grouped by AGR2 expression level [cutoff established on the basis of Roessler liver microarray scores] (C-D, GSE14520) and the cycle threshold value (Ct) obtained by quantitative PCR in our collected HCC specimens (J–K, median, 39-dCt). Patients with high AGR2 levels have worse overall survival and recurrence-free survival. E–I The AGR2 levels with several parameter correlations are shown. L, M Immunohistochemistry staining showing AGR2 expression in HCC specimens (200X). AGR2 is highly expressed in tumor tissues compared with normal liver tissue. d: delta
The clinical significance of AGR2 expression was analyzed in the Roessler Liver database (GSE14520) and in our collected cohort (Fig. 1C–K). Patients with lower AGR2 expression (last 40%) had a better OS rate (log-rank P < 0.05; AGR2 high (top 40%): standard error, 2.738; 95% CI 36.196–46.927; AGR2 low (last 40%): standard error, 2.432; 95% CI 45.322–54.857) and RFS rate (log-rank P < 0.05; AGR2 high (top 40%): standard error, 2.766; 95% CI 27.398–38.243; AGR2 low (last 40%): standard error, 2.645; 95% CI 36.365–46.734) (Fig. 1C, D). The Roessler Liver microarray dataset (GSE14520) provides detailed clinical information; therefore, the correlation between AGR2 and various clinical parameters was statistically analyzed to define the role of sorafenib-regulated AGR2 in HCC progression (Tables 1–3). High AGR2 expression was significantly correlated with high AFP and ALT levels, a high predicted risk metastasis signature score (Fig. 1E, F, G), a large primary tumor size and more advanced pathological stages of HCC (Fig. 1H, I). A high AGR2 level in HCC was correlated with a high serum AFP level (P = 0.002) (Table 1). Univariate analysis showed that male sex (P = 0.009), tumor size (P = 0.045), CLIP score (P = 0.002), BCLC stage (P < 0.001), AJCC stage (P < 0.001), and high AGR2 level (P = 0.003) were significant predictors of worse RFS (Table 2). Multivariate analysis showed that male sex (P = 0.023, HR = 2.124, CI = 1.112–4.058), BCLC stage (P = 0.021, HR = 1.653, CI = 1.077–2.535), AJCC stage (P = 0.019, HR = 1.607, CI = 1.083–2.386) and a high AGR2 level (P = 0.010, HR = 1.572, CI = 1.112–2.224) were independently associated with RFS (Table 2). For OS, univariate analysis showed that cirrhosis (P = 0.023), AFP level (P = 0.011), tumor size (P = 0.001), CLIP score (P < 0.001), BCLC stage (P < 0.001), AJCC stage (P < 0.001), and a high AGR2 level (P = 0.002) were significant predictors of worse OS (Table 3). Multivariate analysis showed that cirrhosis (P = 0.034, HR = 4.563, CI = 1.122–18.555), BCLC stage (P < 0.001, HR = 2.928, CI = 1.908–4.494), and a high AGR2 level (P = 0.008, HR = 1.735, CI = 1.154–2.609) were independently associated with OS (Table 3).
Table 1 Association of AGR2 level (Roessler liver array) with clinicopathologic indicators of hepatocellular carcinomaTable 2 Prognostic significance of clinicopathologic indicators and AGR2 for recurrence-free survival in the Roessler liver arrayTable 3 Prognostic significance of clinicopathologic indicators and AGR2 for overall survival in the Roessler liver arrayMoreover, our HCC specimen cohort was also analyzed. We utilized qRT–PCR to examine the levels of AGR2, and a median level of 15.3 (39-△Ct) was defined as the cutoff to divide the HCC specimens into high and low AGR2 expression groups. Similar results were observed; high AGR2 levels were related to significantly worse OS and RFS rates (Fig. 1J, K). Furthermore, the correlation between AGR2 expression and various clinical parameters in our collected cohort was analyzed (Tables 4–6). A higher AGR2 level in HCC was observed in female patients (P = 0.007) (Table 4). A high AGR2 level (39-△Ct ≥ 10.8) in HCC was significantly associated with worse OS (P = 0.016) (Fig. 1J) and RFS (P = 0.045) (Fig. 1K). Univariate analysis showed that cirrhosis (P = 0.022), AFP level (P = 0.033), vascular invasion (P = 0.003), AJCC stage (P = 0.003), and a high AGR2 level (P = 0.047) were significant predictors of worse RFS (Table 5). Multivariate analysis showed that cirrhosis (P = 0.023, HR = 1.600, CI = 1.067–2.401), vascular invasion (P = 0.005, HR = 1.825, CI = 1.198–2.780), and a high AGR2 level (P = 0.043, HR = 1.662, CI = 1.017–2.716) were independently associated with RFS (Table 5). For OS, univariate analysis showed that cirrhosis (P = 0.020), AFP level (P = 0.012), vascular invasion (P < 0.001), AJCC stage (P = 0.007), and a high AGR2 level (P = 0.018) were significant predictors of worse OS (Table 6). Multivariate analysis showed that cirrhosis (P = 0.017, HR = 1.655, CI = 1.093–2.505), vascular invasion (P = 0.001, HR = 2.200, CI = 1.392–3.476), and a high AGR2 level (P = 0.015, HR = 1.945, CI = 1.138–3.324) were independently associated with OS (Table 6).
Table 4 Association of AGR2 expression with clinicopathologic indicators of hepatocellular carcinomaTable 5 Prognostic significance of clinicopathologic indicators and AGR2 for recurrence-free survival in the clinical cohortTable 6 Prognostic significance of clinicopathologic indicators and AGR2 for overall survival in the clinical cohortMoreover, immunohistochemical staining was utilized to examine the expression of AGR2 in 24 clinical HCC tissues, and the results indicated that AGR2 was highly expressed in tumor tissues compared to normal tissues (Fig. 1L, M). Overall, based on the evidence, we found that patients with lower AGR2 expression have a better OS rate; hence, we suggest that AGR2 might play an oncogenic role in HCC progression and thus might be a useful prognostic marker of HCC progression.
Sorafenib decreases cell viability and increases cell apoptosisFirst, to determine the effect of sorafenib on cell viability, HCC cell lines were treated with various doses of sorafenib (5–10 μM, 24–48 h). Cell viability was significantly decreased with sorafenib treatment in a dose-dependent manner in J7, Hep3B, HepG2 and Huh7 cells according to MTT assay results (Fig. 2A–D). Moreover, flow cytometry was utilized to determine whether sorafenib influences HCC cell apoptosis. HepG2 and Huh7 cells were stimulated with 5 and 10 μM sorafenib for 24 h. Cell apoptosis was slightly induced with 5 μM sorafenib; however, the increase in the apoptosis rate compared with that in the control reached approximately 25% after 10 μM sorafenib stimulation in both HepG2 and Huh7 cells (Fig. 2E–H). Based on these results, we found that sorafenib can modulate HCC cell viability and apoptosis ability; subsequently, we evaluated whether AGR2 is involved in sorafenib-regulated phenotypes.
Fig. 2
Sorafenib decreases cell viability, increases cell apoptosis and induces AGR2 secretion in HCC. A–D The viability of J7, Hep3B, HepG2, and Huh7 HCC cells treated with 5 and 10 μM sorafenib for 24–48 h was examined using MTT assay. E–H Cell apoptosis was determined in HepG2 and Huh7 cells after stimulation with 5 and 10 μM sorafenib. The quantification of apoptotic cells is shown in F, H. Sorafenib decreases cell viability and increases cell apoptosis in HCC cells. I–R AGR2 RNA I–L and protein M–R levels, both intracellular M–P and extracellular Q, R, were examined via RT–PCR and Western blotting after 5 and 10 μM sorafenib treatment for 24–48 h. Sorafenib induces AGR2 secretion from the cytosol into conditioned medium rather than exerting transcriptional or translational regulation (lane 1, 4:untreatment; lane 2, 3, 5, 6:sorafenib treatment). Ponceau S was used as an internal control
Sorafenib induces AGR2 secretion instead of transcriptional regulationAfter we retrieved the GSE94550 dataset, AGR2 was found to be induced in sorafenib-resistant Huh7 cells compared to parental cells (Fig. 1A). To further examine whether sorafenib can regulate AGR2 expression in HCC, RT–PCR and Western blotting were applied in parental HCC cell lines treated with sorafenib. First, we found that the RNA level of AGR2 was not altered by sorafenib in J7, HepG2, Huh7 and Hep3B cells (Fig. 2I-L; Additional file 1: Figure S1A–D). However, the protein level of AGR2 in the lysates of these cells was unexpectedly decreased in a dose-dependent manner after sorafenib stimulation, as shown by Western blotting (lane 3 vs 1, lane 6 vs 4; Fig. 2M–P; Additional file 1: Figure S1E–H). Based on these contradictory results, sorafenib regulated the RNA and protein levels of AGR2 in parental HCC cells. Several reports have shown that AGR2 is an ER-resident protein that is also localized in the extracellular matrix, blood and urine [21, 22]. Therefore, we collected CM after 5 and 10 μM sorafenib treatments for 24–48 h. As expected, AGR2 was detected in CM from HepG2 and Hep3B cells treated with sorafenib (lane 2, 3 vs 1, lane 5, 6 vs 4; Fig. 2Q, R; Additional file 1: Figure S1I-J). Based on this evidence, we suggest that sorafenib regulates AGR2 through posttranslational modification, not transcriptional regulation, in parental HCC cells.
AGR2 plays a role in cell viability and apoptosisTo analyze the roles of AGR2 in HCC progression, we established AGR2-silenced Hep3B, HepG2 and Huh7 cells (lane 2 vs 1; Fig. 3A(a), B(a), C(a); Additional file 2: Figure S2), and cell viability was determined using MTT assay. Cell viability was significantly decreased after AGR2 silencing, and the effect was more obvious when AGR2 silencing was combined with sorafenib treatment (Fig. 3A(b), B(b), C(b)). These findings suggest that AGR2 plays a role in promoting cancer progression. Additionally, we investigated whether AGR2 affects cell apoptosis. Flow cytometry analysis was utilized to demonstrate that sorafenib can induce significant cell apoptosis (approximately 10%) in Hep3B, HepG2 and Huh7 cells compared with control cells, which was more conspicuous after AGR2 silencing in the presence of sorafenib compared to the control (siNC) (Fig. 3D, E, F). Quantitative results are shown in panels 3D (b), E (b), and F (b). Apoptotic cells appeared among the siNC cells under untreated conditions, and we speculate that apoptosis might have been induced in these cells by the transfection process or the apoptosis assay procedures. However, the phenomenon of sorafenib-treated or AGR2-silenced cell apoptosis was not influenced by the basal apoptosis signal in untreated cells. The sorafenib-treated and AGR2-silenced cell apoptosis rates were normalized to the value in the untreated basal cells. However, we found that AGR2 can be secreted into CM and detected by Western blotting (CM, Fig. 2Q, R). Previously, Fessart et al. reported that extracellular AGR2 can be defined as an extracellular matrix pro-oncogenic regulator that makes cancer cells more aggressive [21]. Hence, we evaluated whether the addition of recombinant AGR2 protein regulates cell viability and apoptosis. Flow cytometry analysis indicated that 10 μM sorafenib can induce up to 30–40% cell apoptosis at 24 h in both HepG2 and Huh7 cells; however, the phenomenon was reversed with 60 ng/ml recombinant AGR2 (rAGR2) (Fig. 3G, H). Based on this evidence, we suggest that AGR2 secreted into CM in response to sorafenib stimulation plays an oncogenic role in HCC cancer progression.
Fig. 3
AGR2 is implicated in sorafenib-regulated cell viability and apoptosis. A–C The viability of Hep3B, HepG2, and Huh7 cells with silenced AGR2 was examined after 24 h in the presence or absence of sorafenib (5 μM) using MTT assay. The AGR2 levels (A(a), B(a), C(a)) and cell viability (A(b), B(b), C(b)) results are presented. AGR2 supports HCC cell viability. D–H HCC cell apoptosis was assessed after AGR2 silencing D–F or stimulation with 60 ng/ml recombinant AGR2 (rAGR2) G, H in the presence of 10 μM sorafenib. The quantification of apoptotic cells is shown (D(b), E(b), F(b)). AGR2 inhibition, using either siRNA transfection or recombinant protein treatment, decreases cell apoptosis. siNC: negative control siRNA, siRNA vector only; siAGR2: AGR2 siRNA
Sorafenib induces ER stress in HCCWe determined the functions and regulatory mechanisms through which sorafenib influences AGR2 activity. Based on a literature search, AGR2 has been demonstrated to be upregulated upon ER stress, and ER stress-related molecules, such as protein kinase R (PKR)-like endoplasmic reticulum kinase (PERK), inositol-requiring enzyme 1 (IRE1) and activating transcription factor 6 (ATF6), are dysregulated in many cancer types [16]. Therefore, the relationship between sorafenib and ER-related factors was determined. Among these molecules, using Western blotting, we found that phospho-IRE1α (p-IRE1α) was upregulated by 10 μM sorafenib treatment in HepG2 and Huh7 cells (lane 3 vs 1; Fig. 4A; Additional file 3: Figure S3). The Bip protein level was determined using Western blotting after sorafenib treatments at numerous concentrations in J7 and Huh7 cells. However, Bip expression was not altered by sorafenib (lane 2, 3 vs 1; Additional file 4: Figure S4). Moreover, X-box binding protein 1 (XBP1) has been reported as a unique transcription factor that modulates ERAD gene expression and promotes protein folding [33]. In the UPR, IRE1α is activated via oligomerization and autophosphorylation, followed by the activation of its endoribonuclease to cleave and splice XBP1. The activated IRE1α endoribonuclease can remove 26 nucleotides from the intron of XBP1, converting XBP1 from preform XBP1 (XBP1 u: unspliced) to activated XBP1 (XBP1 s: spliced) [34]. Therefore, RT–PCR was used to detect the status of the IRE-1 downstream factor XBP1. Through RT–PCR analysis, we found that XBP1 was spliced from inactive XBP1 u to active XBP1 s after stimulation with sorafenib for 24 and 48 h in HepG2 and Huh7 cells (lane 3 vs 1, lane 6 vs 4; Fig. 4B, C; Additional file 5: Figure S5A, B). The data showed two bands: the upper band is full-length (unspliced, u) XBP1 (XBP1 u), and the lower band is spliced (s) XBP1 (XBP1 s). Therefore, we found that spliced XBP1 was increased and unspliced XBP1 was decreased after sorafenib treatment for 24 and 48 h. Subsequently, we sought to verify whether AGR2 plays a role in the conversion from inactive XBP1 u to active XBP1 s. As expected, the sorafenib-induced XBP1 s levels were more robust in HepG2 and Huh7 cells with silenced AGR2 (siAGR2) compared with vector control (siNC) under sorafenib treatment (lane 4 vs 2; Fig. 4D; Additional file 5: figure S5C, D). In contrast, we further verified whether added recombinant AGR2 (rAGR2) could modulate the splicing of inactive XBP1 u to activate XBP1 s. Similarly, the levels of XBP1 s induced by 10 μM sorafenib were reduced after stimulation with 60 ng/ml rAGR2 in HepG2 and Huh7 cells (lane 4 vs 3; Fig. 4E; Additional file 5: figure S5E, F). Moreover, we analyzed AGR2 and XBP1 expression in our HCC clinical specimen cohort using RT‒PCR. We analyzed 9 normal liver tissues and 5 HCC tumor tissues. AGR2 was slightly more highly expressed in HCC tumor tissues than in normal tissues, and the spliced XBP1 (XBP1 s) level was slightly more highly expressed in HCC tissues. We suggest that the correlation between AGR2 and XBP1 needs to be further demonstrated in more clinical specimens in the future (Normal: lanes 1–9, Tumor: lanes 1–5; Additional file 6: figure S6). Collectively, these findings indicate that sorafenib induces HCC ER stress via the IRE1α-XBP1 cascade through AGR2 regulation.
Fig. 4
Sorafenib regulates ER stress-related molecules. A–E The protein A and RNA B, C, D, E levels of ER stress-related molecules were examined by Western blotting A and RT–PCR B, C, D, E after treatment of HepG2 A, D, E (left), B and Huh7 A, D, E (right), C cells with 5 μM and 10 μM sorafenib with or without AGR2 silencing D and 60 ng/ml recombinant AGR2 (rAGR2) stimulation E. Sorafenib induces the dysregulation of several ER stress-related molecules. AGR2 is involved in sorafenib-induced XBP-1 splicing. C: cleavage; u: unspliced; s: spliced; siNC: vector control siRNA; siAGR2: AGR2 siRNA. 0: untreatment; 5, 10: sorafenib treatment
AGR2 plays diverse rolesTo determine whether AGR2 plays diverse roles in resistant sublines compared to sorafenib-sensitive HCC cells, HepG2 sorafenib-resistant (HepG2-SR) and Huh7 sorafenib-resistant (Huh7-SR) cells were established (Fig. 5A, B). We applied 7 µM sorafenib to the culture medium to generate sorafenib-resistant cell lines for the following experiments. Using MTT assay, we found that cell viability was decreased by approximately 50% after sorafenib treatment (7 μM) in parental HepG2 and Huh7 cells (indicated as PCs); however, sorafenib only reduced cell viability by 10–20% in resistant cells (indicated as SR cells) (Fig. 5A, B), indicating that these SR cell lines are protected against sorafenib challenge. We found that AGR2 was related to resistance and upregulated by sorafenib in the GSE94550 dataset (Fig. 1A, B). Therefore, we verified whether this regulation was observed in our sorafenib-resistant cells. As expected, AGR2 was highly expressed intracellularly in sorafenib-resistant HepG2 and Huh7 cells compared with parental cells (lane 2 vs 1; Fig. 5C; Additional file 7: figure S7A). We also found that sorafenib reduced intracellular AGR2 levels in HepG2-SR and Huh7-SR cells (lane 3 vs 1; Fig. 5D; Additional file 7: figure S7B), and the tendency was similar to that of sorafenib-regulated AGR2 in HepG2-PCs and Huh7-PCs (Fig. 2M–P). Moreover, we found that sorafenib can induce AGR2 secretion in CM in HepG2-SR and Huh7-SR cells compared with controls. The effect was stronger in resistant cells than in parental cells (lane 4 vs 3; Fig. 5E; Additional file 7: figure S7C). Collectively, these results indicate that AGR2 induction was more robust in both the cell lysate and CM of sorafenib-resistant cells than in parental cells in the presence and absence of sorafenib. This finding might explain why SR cells are protected against sorafenib.
Fig. 5
AGR2 regulation in sorafenib-sensitive and sorafenib-resistant cells. A–E Cell viability A, B and AGR2 regulation C–E in both cell lysate C, D and CM (CM, E) from parental cells (PC) and sorafenib-resistant (SR) cells treated with sorafenib. Sorafenib-resistant cells show higher cell viability, and AGR2 is highly expressed in sorafenib-resistant cells compared to sorafenib-sensitive cells. 0: untreatment; 5, 10: sorafenib treatment
Sorafenib-resistant cells modulate ER stress and reduce cell apoptosisTo determine whether sorafenib-resistant cells can resist the effect of sorafenib-induced cell apoptosis, flow cytometry analysis was utilized to demonstrate that cell apoptosis was induced after stimulation with various doses of sorafenib for 24 h in HepG2 (Fig. 6A, B) and Huh7 (Fig. 6C, D) parental cells. The ratio of cells undergoing sorafenib-induced apoptosis reached approximately 30% among parental cells; however, the effect was not observed in sorafenib-resistant cells (Fig. 6A–D). Based on this evidence, we suggest that these resistant cells are protected against sorafenib toxicity, which prolongs cancer cell viability. Furthermore, we analyzed whether AGR2 plays a vital role in inducing HCC resistance to sorafenib. We silenced AGR2 in HepG2 SR and Huh7 SR cells and performed an apoptosis assay. The results indicated that sorafenib can induce cell apoptosis, but the effect was more robust after AGR2 knockdown (Fig. 6E–H). According to these findings, we speculate that AGR2 plays a critical role in inducing sorafenib resistance in HCC.
Fig. 6
AGR2 is involved in HCC resistance to sorafenib. A–D Apoptosis assay performed after stimulation of both HepG2 and Huh7 parental cell s (PC) and sorafenib-resistant (SR) cells with sorafenib at various concentrations (0–10 μM). The apoptosis rate was lower in sorafenib-resistant cells (indicated as SR cells) than in parental cells (indicated as PCs). Sorafenib-resistant cells have higher protection and resistance to sorafenib-induced toxicity. E–H Apoptosis assay performed in HepG2 SR and Huh7 SR cells with AGR2 silencing in the presence of sorafenib (10 μM). More apoptotic of HepG2 and Huh7 cells were observed in the AGR2-silenced SR group than in the vector control group. Sorafenib-induced apoptosis is more obvious with AGR2 silencing. siNC: negative control siRNA, siRNA vector only; siAGR2: AGR2 siRNA
We found that the ER stress-related molecule p-IRE1α was induced by sorafenib treatment; subsequently, we determined whether p-IRE1α regulation occurred in sorafenib-resistant cells. Western blot analysis revealed that the levels of p-IRE1α were decreased after sorafenib stimulation in SR HepG2 and Huh7 cells and that the effect was inconsistent in PCs (lane 2 vs 1; Fig. 7A, B; Additional file 8: Figure S8A). Moreover, according to RT–PCR results, we found that the sorafenib-induced changes in the levels of XBP1 s, a downstream factor of IRE1α, were abolished or attenuated in HepG2-SR and Huh7-SR cells and that this was not observed in parental HepG2 and Huh7 cells (lane 6 vs 3; Fig. 7C; Additional file 8: Figure S8B). Furthermore, we evaluated whether the effect was mediated by AGR2 in sorafenib-resistant cells. The data indicated that XBP1 s expression was induced after silencing AGR2 expression and was further elevated after stimulation with sorafenib (lane 4 vs 2; Fig. 7D; Additional file 8: Figure S8C). We analyzed the cleaved (C) caspase3 and AGR2 levels in tumors from nude mice subcutaneously injected with Huh7 PCs and SR cell lines, both treated with sorafenib (defined in the figure as sora PC and sora SR, respectively). Immunohistochemistry results indicated that AGR2 was more highly expressed in sora-treated SR cells than in sora-treated PC cells; in contrast, c-casp3 was slightly more highly expressed in sora-treated PC cells than in sora-treated SR cells (Additional file 9: Figure S9). This suggests that AGR2 is more essential under cell stress conditions. Collectively, the contradictory evidence in sorafenib-sensitive and sorafenib-resistant cells may indicate that cells can resist sorafenib toxicity by modulating ER stress-related molecules to decrease cellular ER stress.
Fig. 7
AGR2 is involved in the sorafenib-regulated IRE1α-XBP1 cascade. A, B, C, D The RNA C, D and protein A, B levels of the ER stress-related molecules p-IRE1α, IRE1α A, B, and XBP1 (u and s, C, D) were examined via Western blotting A, B and RT–PCR C, D after AGR2 silencing D and treatment with sorafenib at concentrations ranging from 0 to 10 μM in both parental (PC) and sorafenib-resistant (SR) HepG2 (A, (C, D, left)) and Huh7 (B, (C, D, right)) cells. Sorafenib-induced p-IRE-1α, IRE-1α regulation and XBP-1 splicing in sorafenib-sensitive cells are attenuated in sorafenib-resistant cells. u: unspliced; s: spliced; siNC: negative control siRNA, siRNA vector only; siAGR2: AGR2 siRNA. 0: untreatment; 7, 10: sorafenib treatment
Based on the evidence, we propose that sorafenib reduces cell viability and induces cell apoptosis via downregulation of AGR2 in the cell lysate and increased secretion in CM, which induces ER stress via upregulation of p-IRE1α and spliced XBP1 in HCC. However, the phenomenon of sorafenib-induced apoptosis was abolished in sorafenib-resistant cells, and this effect may occur through increased AGR2 expression in cell lysates and downregulation of p-IRE1α and spliced XBP1. Overall, AGR2 might modulate ER stress to protect cells from sorafenib toxicity and extend cell survival (Fig. 8).
Fig. 8
Proposed model of the roles of AGR2 in sorafenib-sensitive and sorafenib-resistant HCC. AGR2 plays different roles in sorafenib-sensitive (PC) and sorafenib-resistant (SR) cells. Sorafenib decreases AGR2 expression in the cell lysate and in turn induces secretion into CM. Sorafenib reduces cell viability and induces cell apoptosis. However, AGR2 is highly expressed in sorafenib-resistant cells, and the induction of AGR2 expression in CM was more robust in sorafenib-resistant cells than in parental cells, which in turn induces cell survival and suppresses cell apoptosis
留言 (0)