Replication timing and transcriptional control: beyond cause and effect — part IV
The faithful duplication of a cell’s genome once per cell cycle is critical to preserve cell identity, genome integrity, and progression through the cell cycle. Two mutually exclusive steps of DNA replication origin licensing in which the Mcm-replicative helicase is first loaded onto DNA (G1 phase), followed by origin firing (S-phase), ensure that the entire genome is replicated once and only once [1]. During S-phase, origins fire at varying time points, giving rise to a distinct temporal order in which segments of the genome replicate, termed the replication timing (RT) program. The RT program is developmentally regulated and aberrations in RT are observed in diseased states such as cancer and progeroid disease 2, 3. The RT program is highly correlated with transcription and features of genome architecture. It is thus an ongoing challenge to understand how these highly correlated structural and functional properties of chromosomes are causally related.
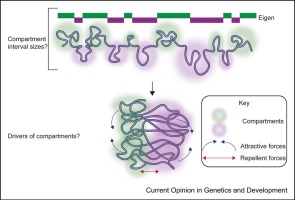
This Current Opinion article is the fourth installment in a review series we have written each seven years on the relationship between replication timing control, nuclear organization, and transcriptional regulation. In our last review, we discussed the contribution of chromatin-conformation capture methods, which confirmed cytogenetic evidence for spatial segregation of early- and late-replicating chromatin and segmentation of replication units into structural domains, allowing the alignment of microscopic features with molecular maps of chromosome folding 4, 5. Since then, new methods that measure contact frequency and distance of loci to nuclear landmarks as well as computational methods to delineate neighborhoods within the nucleus have confirmed that RT closely reflects subnuclear position [6]; late-replicating chromatin preferentially resides near the nuclear lamina, while early-replicating chromatin typically resides in the interior of the nucleus, with the earliest-replicating regions frequently near nuclear speckles 7, 8, 9, 10. Although these features are closely correlated, they have been uncoupled in many experimental systems 11, 12, 13••, so causal links between them remain a major challenge. Here, we focus on recent progress understanding major regulators of RT genome-wide (Rif1), chromosome-wide (ASARs), or domain-wide (ERCEs), as well as how RT can modulate nuclear function.










留言 (0)