
記住我

To examine whether the chorioallantoic membrane (CAM) culture can be used as a research model of lung cancer, we used a 3D co-culture system of cell line-derived organoids that recapitulated human lung cancer tissues (Additional file 1: Fig. S1a) [14]. The organoid consisted of A549-OKS cells, human umbilical vein endothelial cells (HUVECs) and mesenchymal stem cells (MSCs).
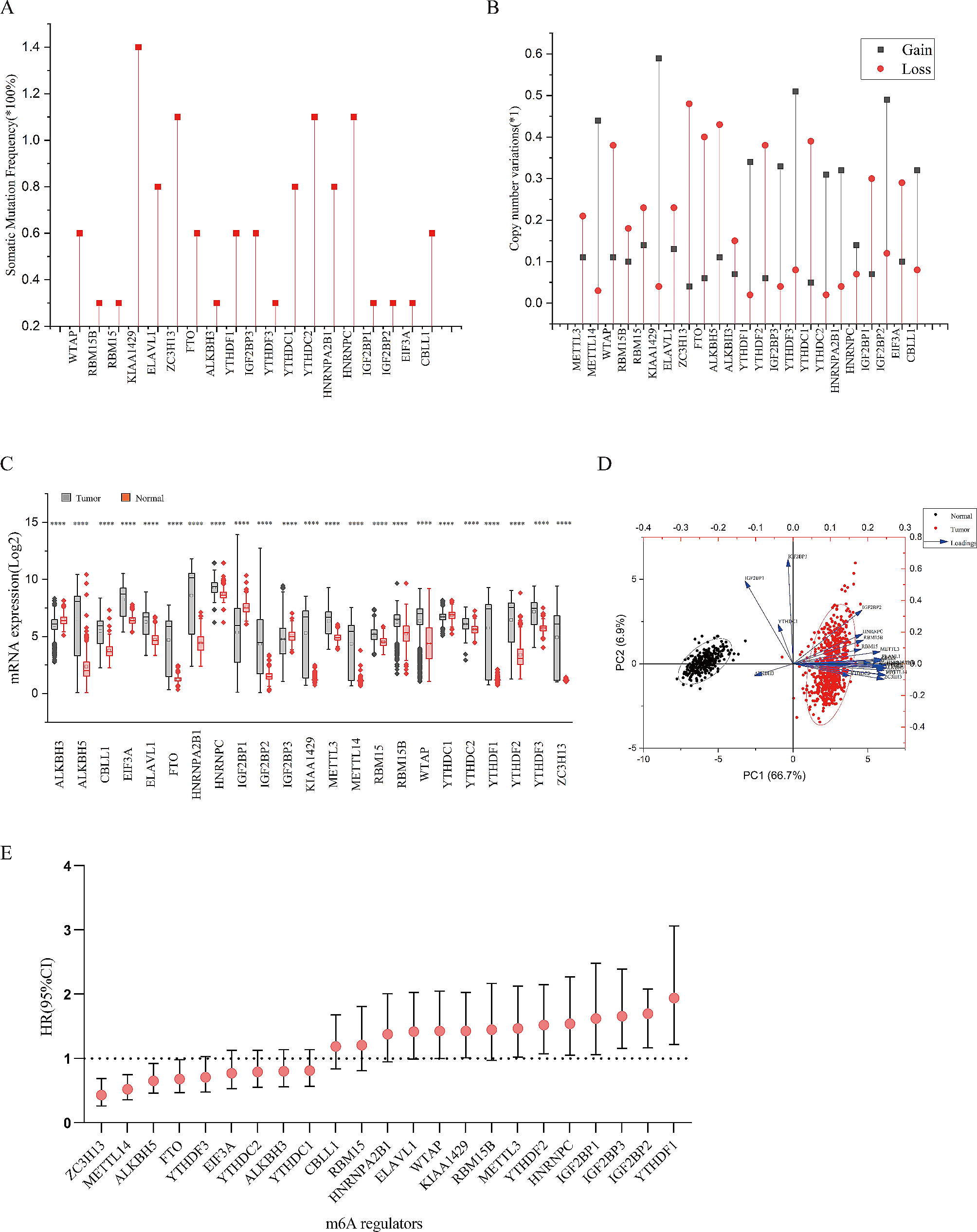
Since the CAM, which is a respiratory organ for chick embryos, rapidly developed from embryonic development day (EDD) 10 to 15 (Additional file 1: Fig. S1b), we used a strategy based on the previously reported CAM assay, as illustrated in Fig. 1a [15, 16, 19]. We transplanted cell line-derived organoids onto the CAM on EDD 10 (Additional file 1: Fig. S1c) and examined whether or not they were grafted five days after implantation. Even though we had no prior experience in performing the CAM assay, we were able to successfully reproduce the previously reported CAM preparation and transplantation methods. The results showed that engraftment on the CAM was observed in 10 of 12 trials (83%) (Fig. 1b and data not shown).
Fig. 1
The chorioallantoic membrane (CAM) assay as a model for researching lung cancer engraftment and angiogenesis. a. A schematic diagram of the CAM assay. b. Macroscopic images are captured on embryonic development day (EDD) 10, 11, 13 and 15. A cell line-derived organoid engrafts the CAM and induces angiogenesis (upper panels). The red arrowheads indicate transplanted organoids. The right image shows an enlarged bottom view of the area enveloped by the yellow dotted lines. The sphere formation medium does not recruit the vessels (lower panels). c. Microscopic images show graft-induced angiogenesis in the CAM assay on EDD 15. A cell line-derived organoid recruited microvessels from thick vessels (right panels). Upper and lower panels were captured at 0.67 × and 1.5 × magnification. Yellow arrowheads indicate new vessels on the mass surface
Tumor cells stimulate the stroma and blood vessels to perform angiogenesis in order to supply nutrients and oxygen to themselves and grow and CAM culture is a well-established platform for evaluating tumor angiogenesis [15, 16, 19], which is defined as new vascular networks generated from preexisting vessels [20]. A closer look around the site where we transplanted the cell line-derived organoids revealed that new tortuous and dilated vessels were formed at the graft surface on the CAM (Fig. 1c right panels). We confirmed that the addition of sphere formation medium (Fig. 1b lower panels, 1c left panels) alone did not cause such abnormal vascularization.
Next, we investigated the interaction between chick vessels and transplanted cell line-derived organoids in our system by immunohistochemistry (IHC). We used Desmin as a smooth muscle marker to detect vessels [21] and Amino acid transporter type 2 (ASCT2) to distinguish human organoids from chicken tissues. ASCT2 is expressed only in mammals, not in birds [22, 23]. IHC to detect cell line-derived organoids graft onto the CAM revealed that the ASCT2-positive organoids invaded the CAM and were surrounded by Desmin-positive vessels (Additional file 2: Fig. S2a). Furthermore, we analyzed whether these vessels were derived from human or chicken using CD31 antibodies that recognize human vascular endothelium. Pre-transplant organoids in vitro contained CD31-positive cells (Additional file 2: Fig. S2b left panel) [21], but not Desmin-positive cells (Additional file 2: Fig. S2b right panel). However, organoids on the CAM after transplantation did not contain CD31-positive cells (Additional file 2: Fig. S2c). These results suggested that our transplanted organoids were supplied by chick vessels alone.
Invasion and angiogenic ability of A549-OKS cells in the CAM assayWe have previously shown that A549-OKS cells have the ability to promote invasion in comparison to A549-GFP cells in a double-layer invasion assay [14]. Moreover, it was also shown that only A549-OKS cells can form spheres containing HUVEC cells in mixed culture with MSC and HUVEC cells [14]. We investigated whether these in vitro properties of A549-OKS cells could be recapitulated in vivo as the ability of “invasion” and “angiogenesis” by transplantation of only A549-OKS or A549-GFP cells into the CAM and observed the direct interaction between human cancer cells and chicken stroma. The survival rate of the eggs in medium only, A549-GFP, and A549-OKS was 50% (8 of 16 trials), 70.6% (12 of 17 trials) and 57.1% (12 of 21 trials), respectively. Moreover, grafts of A549-GFP and A549-OKS cells were observed in 50% (6 of 12 trials) and 66.6% (7 of 12 trials) on EDD 15, respectively (Fig. 2a).
Fig. 2
Invasion and angiogenesis of A549-OKS cells on the CAM. a. Macroscopic images of CAM engrafted with A549-GFP (left panel) and A549-OKS cells (right panel) on embryonic development day (EDD) 15. The part surrounded by the yellow dotted line indicates the grafts. b. Microscopic images of CAM engrafted with A549-GFP and A549-OKS cells on EDD 15. Linear blood vessels are distributed on the surface of the A549-GFP graft. A549-OKS cells make thick vessels twist and induce microvessels. Upper and lower panels are captured in 0.67 and 1.5 × magnification. Yellow arrowheads indicate capillary vessels on the transplanted graft. c. A cross-section view of Fig. 2 b. The right panel shows an enlarged image of A549-OKS cells on the CAM. A549-OKS cells invade the CAM, and the surrounding stroma is thickened. The green, blue, and red dotted lines indicate the surface, bottom, and transplanted graft, respectively. Scale bars: 2.5 mm. d. Immunohistochemistry (IHC) to detect Desmin and ASCT2 of CAM with A549-GFP or A549-OKS cells on EDD 15. Desmin-positive cells (shown in red) and ASCT2-positive cells (shown in green) indicate the chick vessels and the transplanted cells, respectively. The grafts are on the membranes in A549-GFP (left panels) and induce a few vessels. The central panel shows that A549-OKS cells with microvessels invade the membrane. The right panel shows cells located away from the mass. Hoechst was used to stain nuclei. Scale bars: 100 μm
Since A549-OKS cells showed the increased expression of genes related to the GO terms of “blood vessel development (GO: 0001568)” and “vasculature development (GO: 0001944)”, in comparison to A549-GFP cells in our previous report [14], we investigated the A549-OKS cell-induced angiogenetic activity using the CAM assay. The results revealed that A549-OKS cells twisted pre-existing vessels and induced tortuous vessels. In contrast, A549-GFP cells recruited small linear vessels (Fig. 2b lower panels). Hence, the blood flow was magnified and observed using a previously reported technique [24]. The TOKU Capillaro allowed us to visualize the movement of red blood cells in vessels and arterial and venous blood flows from the mass, which could not be confirmed with conventional tissue specimens or under microscope. In the non-transplantation group, straight-like vessels branching from the thin vessels were observed (Additional file 8: Video S1). In A549-GFP cells, the blood vessels were tortuous and fed into the mass (Additional file 9: Video S2). A549-OKS cells formed a white mass that was covered with tortuous blood vessels. These vessels showed irregular blood flow and formed a capillary network. Furthermore, we found that the red blood cells, which drained from the graft, flowed to the graft through this capillary network again (Additional file 10: Video S3). It was noteworthy that we could confirm the graft-induced angiogenesis and complex blood flow at magnification.
The cross-sectional image of the area around the graft revealed the graft invasion and peri-tumoral edema of the chicken stroma surrounding the graft only when A549-OKS cells were transplanted (Fig. 2c). To confirm the interaction between A549-GFP or A549-OKS cells and chick vessels, we performed immunohistochemical staining of the area around the graft. When A549-GFP cells were transplanted, ASCT2-positive grafts were found above the CAM and did not invade the CAM (Fig. 2d left panels). Furthermore, Desmin-positive vessels were observed mainly near the grafts. In contrast, A549-OKS (ASCT2-positive) cells invaded the CAM and were covered by Desmin-positive blood vessels. Some cells were located away from the invasive area (Fig. 2d middle and right panels). These results may indicate that A549-OKS cells recruited vessels for growth and invasion. The enhanced features of A549-OKS cells were recapitulated in the CAM assay.
Quantification of angiogenic activities on the CAMNext, we aimed to quantify the in vivo angiogenic activity on the CAM engrafted with lung cancer cells with Bevacizumab treatment [25, 26], which is known to inhibit VEGF-induced vascularization. First, we confirmed that no antiangiogenic effect and no abnormal vessels were observed in control embryos implanted with filter paper soaked in phosphate-buffered saline (PBS) alone (Additional file 3 Fig. S3a). We placed filter paper containing Bevacizumab (12.5 μg) on the CAM at EDD 12 [27] and found the inhibition of chick microvessel formation around the paper after two days (Additional file 3: Fig. S3b).
Based on these results, we set the timing of transplantation and Bevacizumab treatment as shown in Fig. 3a. We implanted the organoids onto the CAM on EDD 10, put the filter paper soaked with or without Bevacizumab near the graft on EDD 12 and observed the vessel formation on EDD 14. In the non-transplantation group, the effect of Bevacizumab on microvessels was unclear (Fig. 3b upper panels). In the non-treatment group, chick vasculature penetrated the tumor graft and new vessels (indicated with blue arrowheads) were visible at the surface of the tumor grafts (Fig. 3b left lower panel). In contrast, the number of newly-formed macroscopic blood vessels was reduced in Bevacizumab-treated group (Fig. 3b right lower panel).
Fig. 3
Quantification of angiogenic activities in the CAM assay. a. A schematic diagram of treatment with Bevacizumab in the CAM assay. The filter paper soaked with phosphate-buffered saline (PBS) containing Bevacizumab was placed near the graft on embryonic development day (EDD) 12 and the transplant site was observed two days later. b. Macroscopic images from the bottom side. Bevacizumab inhibited tumor angiogenesis. Lower and upper panels show CAMs with and without transplantation. Pictures were captured at 1.5 × magnification. Blue arrowheads indicate the blood vessels that we measured. Blood vessels marked with “※” were preexisting vessels and were excluded from the measurements. c. Dot plots of the vessel area (mm2) and mass area (mm2) measured by Image J. Bevacizumab reduced the vessel area. The left graph shows the vessel area. The right graph shows the mass area. Non-treatment; n = 7, Bevacizumab; n = 9. Mean values ± SE, unpaired t test, **P < 0.01. d. Hematoxylin and eosin (HE) staining is shown on EDD 14. The part surrounded by the red dotted line is the graft. Scale bars: 100 μm
Next, we quantitated the in vivo angiogenic activity by measuring the area of neo-vessels and mass area using Image J (Additional file 3: Fig. S3c) [28,29,30]. All images for measurements are shown Additional file 3: Fig. S3d. In the Bevacizumab-treated group, there was a significant decrease in vessel area (p = 0.0087) and mass area (p = 0.0085) in comparison to the non-treatment group (Fig. 3c).
In the blood flow analysis, the TOKU Capillaro exhibited arterial and venous blood vessels that flowed around the graft and formed capillary webs in the non-treatment groups (Additional file 11: Video S4). In contrast, Bevacizumab treatment inhibited the formation of microvessels, as shown in the macroscopic image (Additional file 12: Video S5). As we expected, Bevacizumab inhibited tumor-induced microvessels on the CAM.
Histologically, hematoxylin and eosin (HE) staining of CAM sections showed necrotic grafts in the Bevacizumab-treated group (Fig. 3d). This indicated that the grafts on the CAM were supported by the chick vessels and could not survive when their formation was inhibited. In conclusion, we established a simple method to quantify the changes in vascular distribution on the CAM by microscopic imaging.
A549-TIPARP cells induced angiogenesis in the CAMIn a previous study, we found the expression of genes involved in “blood vessel development” and “vasculature development” was increased by the introduction of OCT3/4, KLF4 and SOX2 into A549 cells by a GO analysis [14]. Among these genes, we focused on 2,3,7,8-tetrachlorodibenzo-p-dioxin-inducible poly (ADP-ribose) polymerase (TIPARP) [31], which mediates mono-ADP ribosylation and was reportedly upregulated in A549 under P.M 2.5 exposure [32]. Since there are no reports of its association with lung cancer-angiogenesis, we investigated whether TIPARP is involved in the enhancement of angiogenesis by the introduction of three lung cancer-related factors using CAM assay.
First, we generated a stable line by overexpressing TIPARP in A549 cells using a retroviral vector and confirmed that the mRNA level of TIPARP was elevated in TIPARP-transduced A549 cells (A549-TIPARP) in comparison to A549-GFP cells, based on semi-quantitative RT-PCR (Fig. 4a). Cell morphologies were not changed by the transduction of TIPARP (Additional file 4: Fig. S4a).
Fig. 4
Transplantation of A549-TIPARP cells induced angiogenesis in the CAM assay. a. Confirmation of the overexpression of TIPARP in A549 cells by semi-quantitative RT-PCR. GAPDH is used as a loading control. Water is used as a negative control. RT, reverse transcriptase. b. Macroscopic (upper panels) and microscopic (lower panels) images of CAM engrafted with A549-GFP cells and A549-TIPARP cells on EDD 15. A549-TIPARP cells induce angiogenesis on the CAM. Yellow dotted lines enveloped the mass. c. The vessel area (mm2) and mass area (mm2) were measured by Image J and are shown as dot plots in the left panel and right panel, respectively. The results were obtained from eight independent experiments. Mean values ± SE, unpaired t test **P < 0.01 and ***P < 0.001. d. Immunohistochemistry (IHC) to detect the Desmin expression of CAM engrafted with A549-GFP cells (left panels) and A549-TIPARP cells (right panels) on EDD 15 to stain chick vessels. Higher magnification images of the region outlined by yellow dotted lines in upper panels are shown in the lower panels. Red arrowheads indicate Desmin-positive chick vessels. Scale bars: 100 μm
Next, we engrafted A549-TIPARP cells and A549-GFP cells onto the CAM. A549-TIPARP cells were found on the CAM, thickened with microvessels on EDD 15 (Fig. 4b right panels). In comparison to the slight convergence of blood vessels in A549-GFP cells (Fig. 4b left panels), a robust neovascular response converging toward the graft was observed when A549-TIPARP cells were transplanted (Fig. 4b right panels). We independently repeated transplantation to CAM three times and the images of each experiment were also shown in Additional file 4: Fig. S4b. When the vessel area and the mass area were quantified, it was found that both were significantly increased when A549-TIPARP cells were transplanted (p = 0.0029 and 0.0004, respectively). (Fig. 4c). In the blood flow analysis, the graft of A549-TIPARP cells was covered with blood vessels and the arterial and venous vessels were connected (Additional file 13: Video S6), similarly to when A549-OKS cells were transplanted.
The IHC analysis of the CAM showed A549-TIPARP cells induced many Desmin-positive vessels, in comparison to A549-GFP cells (Fig. 4d). It shows that IHC could recapitulate the distribution of blood vessels in the macroscopic images. These results indicate that TIPARP was an important gene for angiogenesis in A549-OKS.
TIPARP enhances genes related to the remodeling of extracellular matrixTo reveal TIPARP-dependent molecular networks in angiogenesis, we compared the global gene expression profiles of A549-TIPARP cells and control A549-GFP cells. A scatter plot showed the overexpression of TIPARP in A549 cells with 17.2-fold increase, on average, in comparison to control (Fig. 5a). In A549-TIPARP cells, we identified 202 upregulated genes and 57 downregulated genes with a more than twofold change (Additional file 5 and 6: Table S1 and S2, respectively). Among the upregulated genes, mannose receptor C type 2(MRC2), which showed the second highest fold change (= 7.1) after TIPARP, is known to be involved in extracellular matrix (ECM) remodeling and showed an increased expression level with good reproducibility [33]. Moreover, we performed a pathway analysis for 202 upregulated genes and found twelve significant pathways (p < 0.001, Fig. 5b).
Fig. 5
TIPARP enhances genes related to the remodeling of the extracellular matrix. a. Scatter plot of the mRNA expression comparing the average values of A549-TIPARP cells (n = 3, y-axis) to the average values of A549-GFP cells (n = 3, x-axis). The RNA expression value is shown on a log 2 scale. Green lines indicate a two-fold differences in the expression level. TIPARP, MRC2 and MMP7 are shown in magenta. b. A pathway analysis for upregulated genes (fold-change > 2) in A549-TIPARP cells was performed. The top twelve pathways (P < 0.0001) are shown. Pathways related to the extracellular matrix are shown in red
Among these twelve pathways, seven (red-colored in Fig. 5b) were related to the extracellular matrix (ECM) and four were related to ECM degradation and remodeling (Fig. 5b): assembly of collagen fibrils and other multimeric structures, collagen biosynthesis and modifying enzymes, collagen degradation, and collagen chain trimerization.
Matrix metalloproteinase 7 (MMP7), which is upregulated in A549-TIPARP cells, is involved in three pathways: matrix metalloprotease, activation of matrix metalloprotease, and collagen degradation. On the other hand, the pathway analysis for 57 downregulated genes showed 18 significant pathways (p < 0.001) (Additional file 7: Table S3), including pro-inflammation cytokine associated pathways.
From these results, TIPARP might promote angiogenesis via the remodeling of the extracellular matrix and inhibition of pro-inflammation cytokine signaling on the CAM.
留言 (0)