Tuberculosis (TB), a chronic respiratory infection caused by Mycobacterium tuberculosis, is the leading cause of death from a single source of infection and is becoming an important public health concern [1]. In 2020, approximately 10 million new active TB cases were diagnosed, and more concerningly, 1.28 million people died from the disease [2]. The standard treatment regimen recommended by the World Health Organization (WHO), which consists of four different drugs taken orally over a six-month period, including isoniazid (INH), rifampicin (RIF), pyrazinamide (PZA), and ethambutol (EMB), has been shown to curb the progression of TB; however the hepatotoxicity caused by these drugs poses a massive challenge to clinical treatment [3]. Anti-tuberculosis drug-induced liver injury (ATB-DILI) is a common serious adverse event observed during the clinical treatment of TB [4] and is associated with increased mortality [5]. Compared with developed countries, the incidence of ATB-DILI is much higher in developing countries, especially Iran (27.7%) and Pakistan (19.7%) [6]. A previous study involving 4577 patients on standard anti-tuberculosis (anti-TB) drug regimens in four cities in China showed that 10.47% of patients stopped TB treatment because of drug-induced liver injury [7]. Therefore, ATB-DILI is one of the most important cause of treatment interruption in patients with TB, leading to a range of adverse effects and subsequent drug resistance in TB. However, the mechanisms underlying ATB-DILI remain largely unknown.
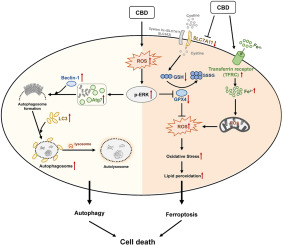
Experimental and clinical studies have shown that anti-TB drug metabolites can lead to excessively increased levels of reactive oxygen species (ROS) in the liver, which is deeply involved in ATB-DILI [8,9]. During the metabolism of these drugs in the liver, large amounts of intracellular glutathione are depleted, leading to increased lipid peroxidation and iron ion concentrations [10]. Lipid accumulation is exacerbated by lipid metabolism disorders, which contribute to the pathogenesis of anti-TB drug-induced hepatotoxicity [11]. Ferroptosis is a lipid-ROS- and iron-dependent form of regulated cell death (RCD) that plays a crucial role in organ injury [12]. It is driven by the loss of activity of the lipid repair enzymes glutathione peroxidases (GPxs), and the subsequent redox imbalances and increased levels of lipid-based ROS, particularly membrane lipid hydroperoxides [13]. Although a recent study indicated that ferroptosis is involved in INH- and RFP-induced lipid peroxidation and hepatocyte death [14], little is known about the role of ferroptosis in ATB-DILI caused by the standard treatment regimen recommended by the WHO.
The cystine/glutamate antiporter solute carrier family 7 member 11 (SLC7A11) is the catalytic subunit of the amino acid transporter system xc− that imports extracellular cystine into the cell, which is subsequently converted to glutathione (GSH) [15]. Glutathione peroxidase 4 (GPx4) is a phospholipid peroxidase that uses reduced GSH as a co-factor to detoxify lipid peroxides to lipid alcohols, thereby suppressing ferroptosis [16]. Correspondingly, GPx4 is widely regarded as a protective factor against ferroptosis in cells [17,18]. Recent studies have demonstrated that the downregulation of SLC7A11 leads to a decrease in intracellular cystine levels and subsequent depletion of glutathione biosynthesis, indirectly suppressing GPx4 activity and subsequent ferroptosis activation [19,20]. Pharmacological inhibition of SLC7A11-mediated cystine uptake, such as erastin-induced ferroptosis [21,22]. Currently, the mechanisms by which ATB-DILI regulates SLC7A11 activity remain largely unclear.
Hypoxia-inducible factor-1α (HIF-1α) is a critical transcription factor that regulates the expression of a large battery of genes involved in oxidative stress, inflammation, and cell death [23]. Studies have suggested that HIF-1α is implicated in the toxicity found in many models of liver injury, including drug-induced liver injury [24], fibrosis [25], and alcoholic liver disease [26]. A case-control study has shown that the HIF-1α gene is associated with a higher risk of ATB-DILI [27]. Nonetheless, the molecular mechanism by which HIF-1α regulates ATB-DILI progression is still unclear. Notably, HIF-1α regulates iron metabolism-related gene expression and promotes tumorigenesis by inhibiting ferroptosis [28,29]. Studies have also suggested that HIF-1α regulates SLC7A11 levels, as it has been shown that the knockout of HIF-1α sharply decreased SLC7A11 protein levels in rat brains [30]. Moreover, overexpression of HIF-1α using a plasmid increased the SLC7A11 protein level in hepatocytes [25]. Therefore, we hypothesized that HIF-1α/SLC7A11 plays a role in anti-TB drug-induced hepatocyte ferroptosis.
Although hepatotoxicity due to clinical anti-TB chemotherapy is well established, the molecular mechanisms by which ferroptosis affects ATB-DILI remain unclear. Here, we assessed the role of ferroptosis in hepatotoxicity induced by first-line anti-TB drugs. Additionally, our study elucidated that anti-TB drug-induced hepatocyte ferroptosis was initiated by the inhibition of the HIF-1α/SLC7A11 signaling pathway. Our findings may provide a potential effective target for future clinical agents for hepatoprotection during anti-TB treatment.








留言 (0)