
記住我










Pulmonary arterial hypertension (PAH) is a severe life-threatening disease that features pulmonary vascular remodelling and perivascular inflammatory cell infiltration [1]. The enhanced T-helper 2 (Th2) cell immune response in PAH is well documented [2, 3]. However, the role of eosinophils (EOS), one of the Th2 effector cells [4], in PAH is seldom reported. A previous study showed that PAH patients with worse syndromes had lower circulating EOS levels [5]; however, the functions of EOS in regulating PAH development and the detailed mechanisms remain unclear.
EOS are granulocytes with segmented nuclei and eosinophilic granules [6] that mature in bone marrow under a tightly regulated process directed by various transcription factors such as GATA1, GATA2 and FOG1 and cytokines such as interleukin 5 (IL5), and are then released into peripheral blood [7, 8]. Chemokines such as C-C motif chemokine ligand 11 (CCL11) (eotaxin-1), CCL24 (eotaxin-2) and CCL26 (eotaxin-3) and cytokines such as IL5 promote EOS transmigration into tissues [8, 9]. Once they have trafficked into tissues, EOS have a function in degranulation and cytokine release [10, 11], DNA release and eosinophil extracellular trap (EET) formation [12–14] and lipid metabolite production [6, 15, 16]. These processes facilitate the interaction of EOS with immune cells [15, 16] and tissue-resident cells [10, 11]. The detrimental roles of EOS have been widely recognised for decades, especially in asthma [4, 12]: EOS interact with mast cells [17], aid B-cells to produce antibodies [18, 19], promote Th2 polarisation [20] and mediate epithelial damage [4, 21]. However, EOS have regained attention in recent years because they show protective effects in maintaining tissue homeostasis in heart [11], lung [15], liver [22], gut [16] and adipose tissue [23, 24], which suggests that EOS have distinct roles in physiology and pathophysiology processes.
To determine the effect, if any, of EOS on PAH development, we explored the function and underlying mechanisms of action of lung EOS in PAH patients and in the sugen/hypoxia (SuHx)-induced PH rat or mouse model. We found that EOS migrated from peripheral blood to lung tissues in PAH patients and PH mice. By using GATA1-deletion (ΔdblGATA) mice and anti-IL5 antibody (TRFK5)-treated mice and rats, we found EOS deficiency worsened PH development by aggravating perivascular inflammation and pulmonary artery smooth muscle cell (PASMC) proliferation in animal models. Finally, we show that EOS function by releasing lipid mediators, among which 14-hydroxy docosahexaenoic acid (14-HDHA) and 17-hydroxy docosahexaenoic acid (17-HDHA) exert a protective effect in PH.
ResultsEOS populations decrease in blood while lung infiltration increases in PAH patientsTo examine changes in EOS abundance during PAH progression, we enrolled 123 IPAH/HPAH patients and 119 age-/sex-matched healthy control subjects (supplementary table S1). Quantification of EOS by routine tests in peripheral blood samples indicated that the EOS percentage in PAH patients was significantly lower than that in sex-matched healthy control subjects (figure 1a). Further classification of PAH patients according to the New York Heart Association revealed that type I/II patients had a higher proportion of EOS in peripheral blood compared with type III/IV patients (figure 1b). These observations were both consistent with previous reports [5]. Notably, loss-of-function mutations of bone morphogenetic protein receptor 2 (BMPR2) are found in IPAH/HPAH, and patients who carry a causative mutation in BMPR2 show more severe vascular dysfunction [29]. Here, we found that carriers of the BMPR2 mutation also had a lower percentage of EOS in peripheral blood than non-carriers (figure 1c). To further examine changes in the abundance of EOS in lung tissues associated with PAH, we conducted EOS staining assays of lung tissue samples from PAH and non-PAH subjects. Hematoxylin and eosin (H&E) staining indicated enhanced EOS infiltration into PAH lung tissues, characterised by the accumulation of red-staining granulocytes with segmented nuclei around remodelled pulmonary vessels (figure 1d). Immunohistochemistry staining showed that eosinophil cationic protein (ECP)+ EOS were more abundant in PAH lungs than in non-PAH lungs (figure 1d, e). Immunofluorescence staining also showed that most ECP+ EOS were located closer to the vimentin+ adventitial layer (figure 1f, arrow a). ECP+ EOS were also found adhering to CD31+ endothelial cells in the vascular lumen (figure 1f, arrow b). These observations suggest a dynamic process of EOS migration, with EOS recruited from the peripheral blood and residing in the parenchyma. In further support of this observed increase in EOS pulmonary migration, the expression of IL5 and eotaxin genes (CCL11, CCL24 and CCL26), which are necessary for EOS trafficking, was significantly elevated in PAH lung tissues (figure 1g), suggesting enhanced EOS recruitment in lungs during PAH development.
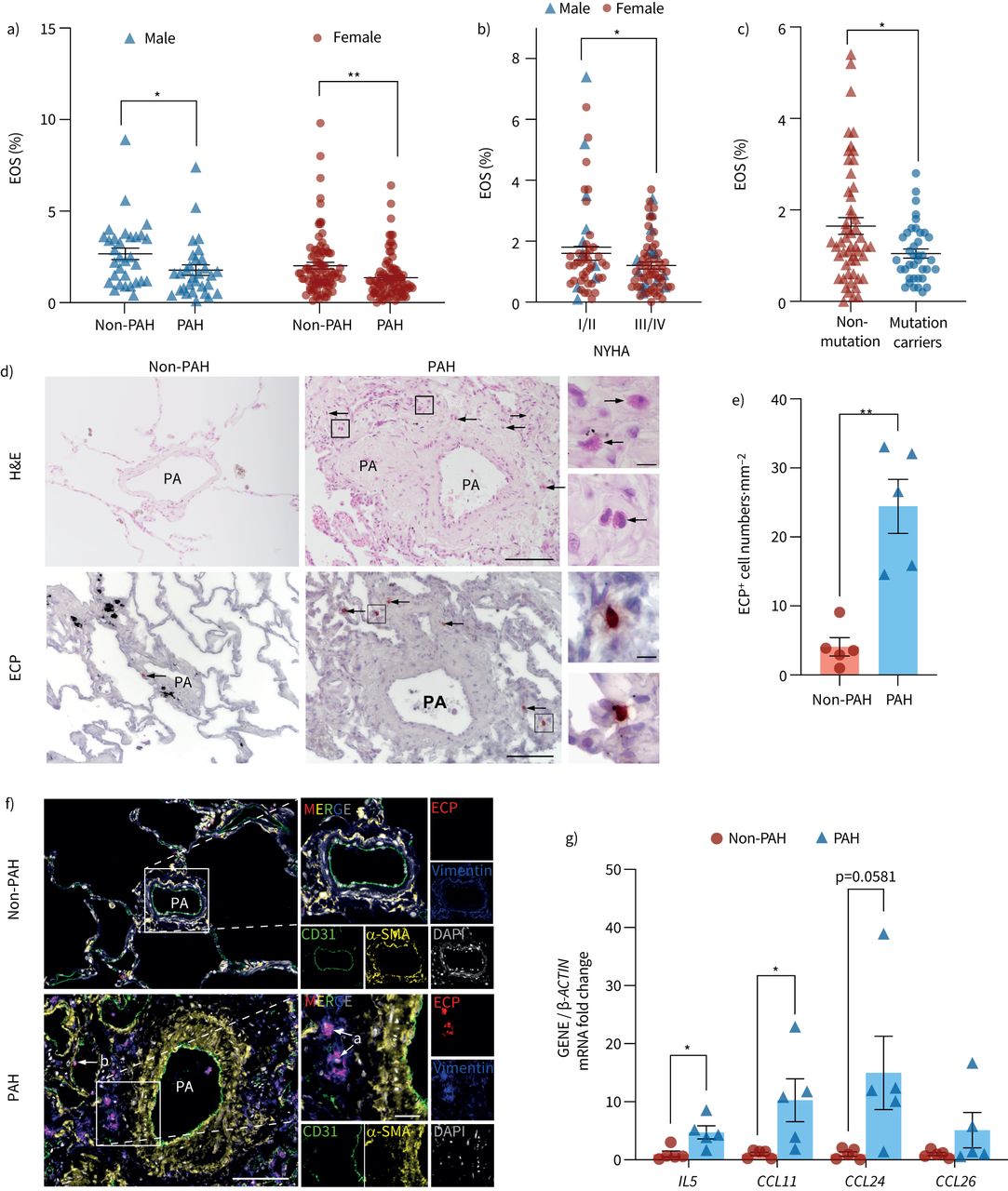 FIGURE 1
FIGURE 1 Increased lung eosinophil (EOS) accumulation in pulmonary arterial hypertension (PAH) patients. a) EOS percentage in blood of male/female PAH patients and sex-/age-matched healthy controls (male: non-PAH n=31, PAH n=30; female: non-PAH n=88, PAH n=93). b) EOS percentage in blood of PAH patients in New York Heart Association (NYHA) functional classes I/II and III/IV (I/II male:female ratio 16:39; III/IV male:female ratio 14:54). c) EOS percentage in blood of non-mutant and BMPR2 mutation-carrying PAH patients (non-mutant n=52; mutation carrier n=38). d) Representative images of haematoxylin and eosin (H&E) and eosinophil cationic protein (ECP) immunohistochemical staining of EOS (arrows) in lung sections from control subjects (non-PAH) or PAH patients. Scale bars: 100 μm (main) and 10 μm (inset). e) Quantification of ECP+ cell numbers in lung sections from human samples (n=5 for each group). f) Representative images of CD31 (green), α-smooth muscle actin (α-SMA) (yellow), vimentin (blue) and ECP (red) immunofluorescent stainings of lung tissues from control subjects and PAH patients. Nuclei were counterstained with DAPI (white). EOS located close to vimentin+ and CD31+ areas are indicated by arrows a and b, respectively. Scale bars: 100 μm (main) and 25 μm (inset). g) Lung mRNA expression of IL5, CCL11, CCL24 and CCL26 (n=5 per group). All data are shown as mean±sem. For a–c, e and g, differences were evaluated by unpaired two-tailed t-test. PA: pulmonary artery. *: p<0.05; **: p<0.01.
Taken together, these results indicate that EOS migrate to lung tissue during the progression of PAH in humans.
Decreased percentage of EOS in blood and increased proportions of EOS in lungs of PH miceTo determine whether these changes in EOS during PAH could also be observed in mice, we next established a SuHx-induced PH mouse model. We conducted flow cytometry analysis by staining for CD45+CD11b+SiglecF+ EOS and found that the percentage of EOS in blood was significantly lower in PH mice than in control mice (figure 2a), while the percentage of EOS in PH lung tissue was significantly higher (figure 2b). EOS infiltration into the lungs was then confirmed by H&E and SiglecF staining (figure 2c). In addition, analyses of cytokine and chemokine expression indicated that both mRNA and protein expression of these markers was elevated in lung tissues of PH mice compared with controls (figure 2d, e). These findings were in agreement with our above results from PAH patients.
 FIGURE 2
FIGURE 2 Increased lung eosinophil (EOS) accumulation in pulmonary hypertension (PH) mice. a) Flow cytometric analyses of CD11b+SiglecF+ EOS among CD45+ cells in peripheral blood of control and sugen/hypoxia (SuHx)-induced PH mice. b) Flow cytometric analyses of CD11b+SiglecF+ EOS among CD45+ cells in lung tissues of control and SuHx-induced PH mice (n=8 per group). c) Representative images of haematoxylin and eosin (H&E) and SiglecF immunofluorescent staining of EOS (arrows) in lung sections from control and SuHx-induced PH mice (left panel, scale bar: 25 μm) and quantification of EOS number in lung section (right panel, n=4 per group). d) Lung mRNA expression of Il5, Ccl11, Ccl24 and Ccl26 (n=5 per group). e) Lung IL5 and CCL11 protein levels in control or SuHx-induced PH mice (n=5 per group). f) The percentage of EOS among CD45+ cells in bone marrow of control or SuHx-induced PH mice (n=8 per group). g) The percentage of EOS among CD45+ cells in spleens of control or SuHx-induced PH mice (n=8 per group). h) The percentage of EOS among CD45+ cells in right ventricles of control or SuHx-induced PH mice (n=6 per group). i) The percentage of EOS among CD45+ cells in left ventricles of control or SuHx-induced PH mice (n=6 per group). All data are shown as mean±sem. Differences were analysed by unpaired two-tailed t-test. ns: nonsignificant. *: p<0.05; **: p<0.01; ***: p<0.001.
No significant changes were observed in EOS levels in either the bone marrow or spleen (figure 2f, g), suggesting that the production of EOS was unaffected during PH progression. However, EOS quantification showed a slight but significant increase in the percentage of EOS in remodelled right ventricle tissues (figure 2h), but no difference from controls in the left ventricle (figure 2i), suggesting that EOS may be related to impaired right ventricular function in PH. In lung tissues, EOS are classified as either resident eosinophils or inflammatory eosinophils based on CD101 expression [20]. Here we found that although there was a slight decrease in the percentage of CD101− resident EOS (from 91.2% to 85.4%) in PH lungs compared with controls (supplementary figure S1a), they were still the predominant subtype in lung and peripheral blood under PH (supplementary figure S1a, b) and the only detectable subtype in bone marrow of mice (supplementary figure S1c).
Collectively, these results show that the proportion of EOS decreases in blood but increases in lungs of PH mice, similar to what was observed in PAH patients.
CCL11 upregulation in adventitial fibroblasts of PH lungs is associated with EOS recruitmentIn light of our observations of pulmonary EOS migration during PH, we next examined EOS trafficking-related cytokine and chemokine expression. Among all genes tested, CCL11 (eotaxin-1) was the most highly expressed in mouse lungs (supplementary figure S2). Subsequent single-cell RNA sequencing (scRNA-seq) analysis of lung tissues from healthy humans (figure 3a) and control mice (figure 3b) revealed that a considerable proportion of fibroblasts expressed high levels of CCL11 in both species. Further comparison of fibroblasts from PH mice showed that Ccl11 was primarily expressed in adventitial rather than alveolar fibroblasts, and that both the percentage of cells and the average expression levels were greater in PH lung tissues than in control (figure 3c). Immunostaining of lung tissue from PAH patients and SuHx-induced PH mice showed that the area of tissue expressing CCL11 overlapped with that expressing platelet-derived growth factor receptor α (PDGFRα), which confirmed that CCL11 was derived from fibroblasts (figure 3d, e). However, CCL11 did not co-localise with CD31, α-smooth muscle actin (α-SMA), CD45 or club cell 10-kDa protein (CC10), which further suggests that endothelial cells, smooth muscle cells, immune cells and epithelial cells were not the main source of the CCL11 in lungs of PH mice (supplementary figure S3a–d). In human and mouse pulmonary adventitial fibroblasts activated by treatment with transforming growth factor β (TGFβ) or platelet-derived growth factor bb (PDGFbb), we found that CCL11 expression was significantly higher than in untreated cells (figure 3f, g). Moreover, CCL11 was not obviously upregulated in similarly treated endothelial cells or smooth muscle cells (supplementary figure S3e, f), allowing us to exclude these cell types as the main source of CCL11 during PH development. Using an EOS adhesion assay, we found that CCL11 stimulation increased EOS adhesion to endothelial cells (figure 3h), supporting the chemotaxis role of CCL11 in EOS recruitment. Collectively, these results suggest that the impaired function of pulmonary adventitial fibroblasts during PAH development results in enhanced CCL11 expression, which in turn promotes EOS migration.
 FIGURE 3
FIGURE 3 C-C motif chemokine ligand 11 (CCL11) expression in pulmonary adventitial fibroblasts. a) Dot plots of gene expression (IL5, CCL11, CCL24 and CCL26) among cell clusters in human. b) Dot plots of gene expression (Il5, Ccl11, Ccl24 and Ccl26) among cell clusters in mouse. c) Violin plots of the expression of Ccl11 among two subgroups of fibroblasts in control and sugen/hypoxia (SuHx)-induced pulmonary hypertension (PH) mice lung tissue. d) Representative images of CCL11 and platelet-derived growth factor receptor α (PDGFRα) immunofluorescent staining of the lung sections from control subjects without pulmonary arterial hypertension (non-PAH) or PAH patients. Scale bar: 25 μm. e) Representative images of CCL11 and PDGFRα immunofluorescent staining of the lung sections from control or SuHx-induced PH mice. Scale bar: 25 μm. f) Relative mRNA expression of CCL11 in human pulmonary adventitial fibroblasts (PAFs) under different treatments (n=3 for three independent repeats). g) Relative mRNA expression of Ccl11 in mouse PAFs under different conditions (n=3 for three independent repeats). h) Representative images and quantification of eosinophil (EOS) adhesive assay. Green arrow points at the 5-(and 6)-carboxyfluorescein diacetate succinimidyl ester (CFSE)-labelled EOS (green). Scale bar: 100 µm. All data are shown as mean±sem. Differences were evaluated by unpaired two-tailed t-test. NK: natural killer; TGFβ: transforming growth factor β; PDGFbb: platelet-derived growth factor bb; ns: nonsignificant. *: p<0.05; ***: p<0.001; ****: p<0.0001.
EOS deficiency aggravates PH in animal modelsTo better understand the functional role of EOS in PH development, we induced PH in male EOS-deficient ΔdblGATA mice by exposing them to SuHx (figure 4a, supplementary figure S4). We first confirmed that the ΔdblGATA mice were deficient in EOS compared with WT mice (figure 4b, quantified in 4c). In addition, ΔdblGATA mice exhibited a significantly higher right ventricular systolic pressure and right ventricular hypertrophy index (weight of the right ventricle/left ventricle+septum (RV/LV+S)) than WT under PH (figure 4d, e), indicating that the deficiency of EOS further aggravated PH development. Immunostaining also showed that vascular thickening (figure 4f, quantified in 4g) and muscularisation (figure 4h) were more extensive in the absence of EOS in PH. Although fibroblasts were implicated in EOS recruitment (figure 3), no significant changes in pulmonary fibrosis were observed in EOS-deficient PH mouse lungs (supplementary figure S5a, b).
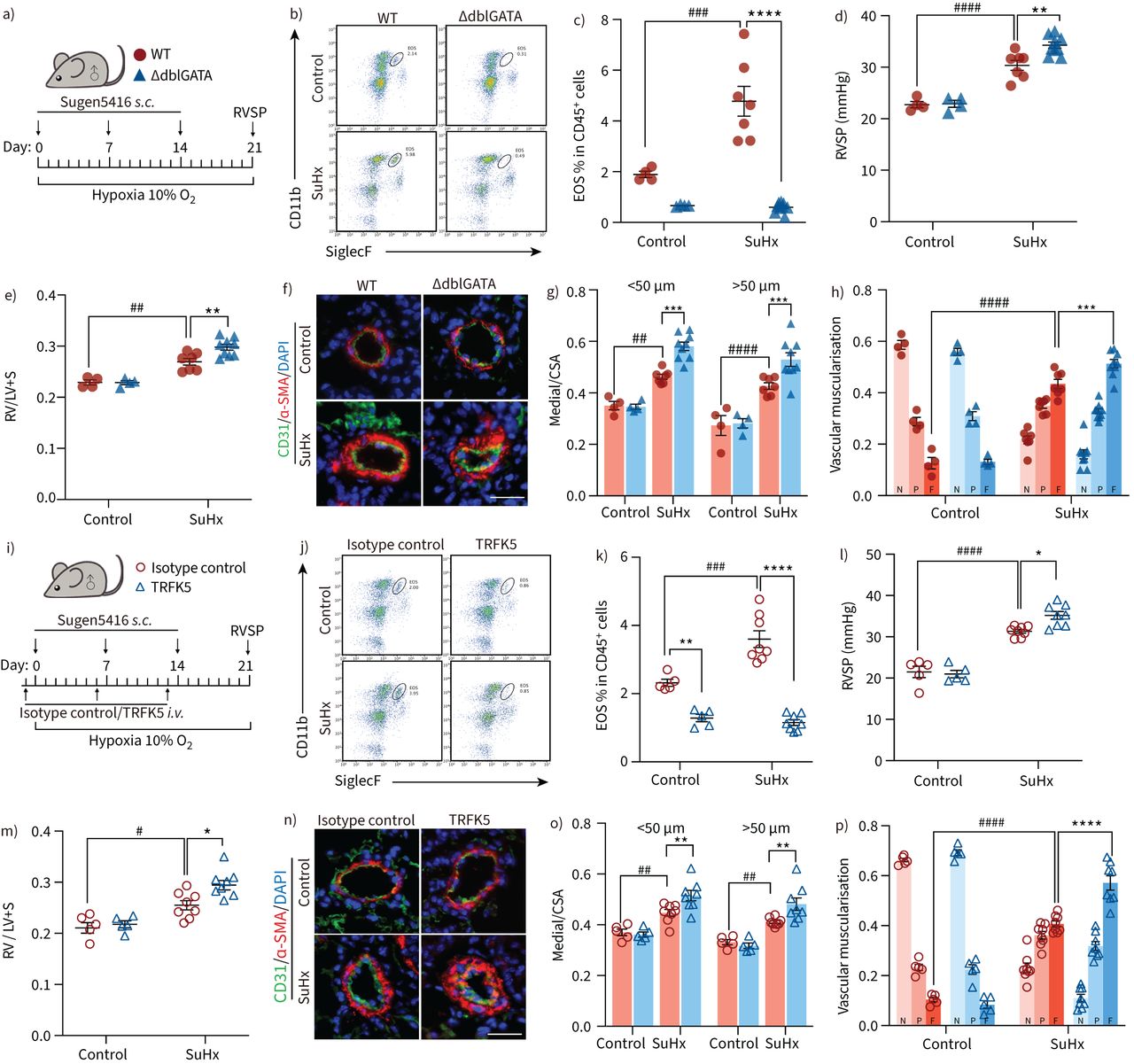 FIGURE 4
FIGURE 4 Eosinophil (EOS) deficiency and depletion exacerbate pulmonary hypertension (PH) in mice. a) Strategy for sugen/hypoxia (SuHx)-induced PH in wild-type (WT) or EOS-deficient (ΔdblGATA) male mice (control: n=4 per group; SuHx: n=7 for WT, n=9 for ΔdblGATA). b) Representative flow cytometric analyses of CD11b+SiglecF+ EOS among CD45+ cells in lung tissues of WT or ΔdblGATA mice. c) The percentage of EOS among CD45+ cells in lung tissues of WT or ΔdblGATA mice. d) Right ventricular systolic pressure (RVSP) of WT and ΔdblGATA mice. e) Right ventricular hypertrophy index (weight of the right ventricle/left ventricle+septum (RV/LV+S)) of the different experimental groups. f) Representative images of α-smooth muscle actin (α-SMA) and CD31 immunofluorescent staining of the lung sections from WT or ΔdblGATA mice. Scale bar: 25 μm. g) Quantification of pulmonary vascular medial thickness to total cross sectional area (CSA) for vessels of 20–50 μm and 50–100 μm in diameter. h) Proportion of vascular muscularisation of pulmonary vessels of 20–100 μm in diameter, classified as non-muscularised (N), partially muscularised (P) or fully muscularised (F). i) Strategy for SuHx-induced PH in male mice injected with isotype control antibody or TRFK5 (control: n=5 per group; SuHx: n=8 per group). j) Representative flow cytometric analyses of CD11b+SiglecF+ EOS among CD45+ cells in lung tissues of isotype control- or TRFK5-treated mice. k) The percentage of EOS among CD45+ cells in lung tissues of isotype control- or TRFK5-treated mice. l) RVSP of isotype control- or TRFK5-treated mice. m) Right ventricular hypertrophy of the different experimental groups. n) Representative images of α-SMA and CD31 immunofluorescent staining of the lung sections from isotype control- or TRFK5-treated mice. Scale bar: 25 μm. o) Quantification of wall thickness of the pulmonary vasculature for vessels of 20–50 μm and 50–100 μm in diameter. p) Proportion of vascular muscularisation of pulmonary vessels of 20–100 μm in diameter, classified as for g. All data are shown as mean±sem. Differences between multiple groups were evaluated by two-way ANOVA with Bonferroni's post hoc test. s.c.: subcutaneous; i.v.: intravenous. *: p<0.05; **: p<0.01; ***: p<0.001; ****: p<0.0001 for ΔdblGATA versus WT mice or TRFK5- versus isotype control-treated mice. #: p<0.05; ##: p<0.01; ###: p<0.001; ####: p<0.0001 for mice in control versus SuHx groups.
 FIGURE 5
FIGURE 5 Eosinophil (EOS) deficiency or depletion promotes neutrophil and monocyte/macrophage accumulation in lung. a) Representative flow cytometric analyses of CD11b+Ly6G+ neutrophils among CD45+ cells in lung tissues of wild-type (WT) or EOS-deficient (ΔdblGATA) male mice (left panel) and the percentage of neutrophils among CD45+ cells (right panel). b) Representative flow cytometric analyses of CD11b+Ly6C+ monocytes/macrophages among CD45+ cells in lung tissues of WT or ΔdblGATA male mice (left panel) and the percentage of monocytes/macrophages among CD45+ cells (right panel). c) Representative images of Ly6G and α-smooth muscle actin (α-SMA) immunofluorescent staining of lung sections from WT or ΔdblGATA male mice (left panel) and quantification of Ly6G+ cell numbers per mm2 lung tissue (right panel). d) Representative images of CD68 and α-SMA immunofluorescent staining of lung sections from WT or ΔdblGATA male mice (left panel) and quantification of CD68+ cell numbers per mm2 lung tissue (right panel). e) Representative flow cytometric analyses of CD11b+Ly6G+ neutrophils among CD45+ cells in lung tissues of isotype control- or TRFK5-treated male mice (left panel) and the percentage of neutrophils among CD45+ cells (right panel). f) Representative flow cytometric analyses of CD11b+Ly6C+ monocytes/macrophages among CD45+ cells in lung tissues of isotype control- or TRFK5-treated male mice (left panel) and the percentage of CD11b+Ly6C+ monocytes/macrophages among CD45+ cells (right panel). g) Representative images of Ly6G and α-SMA immunofluorescent staining of lung sections from isotype control- or TRFK5-treated male mice (left panel) and quantification of Ly6G+ cell numbers per mm2 lung tissue (right panel). h) Representative images of CD68 and α-SMA immunofluorescent staining of lung sections from isotype control- or TRFK5-treated male mice (left panel) and quantification of CD68+ cell numbers per mm2 lung tissue (right panel). Scale bars: 50 μm. Differences between multiple groups were evaluated by two-way ANOVA with Bonferroni's post hoc test. For a–d, control: n=4 per group; sugen/hypoxia (SuHx): n=7 for WT, n=9 for ΔdblGATA. For e–h, control: n=5 for isotype control, n=4 for TRFK5; SuHx: n=8 per group. All data are shown as mean±sem. *: p<0.05; **: p<0.01 for ΔdblGATA versus WT mice, or TRFK5- versus isotype control-treated mice. #: p<0.05; ##: p<0.01 for mice in control versus SuHx groups.
Because IL5 is crucial for EOS maturation and migration [8], depletion of EOS through IL5 neutralisation is applied clinically as a treatment for asthma [30]. Therefore, we depleted EOS by neutralising IL5 in C57BL/6 male mice. After injecting with anti-IL5 antibody (TRFK5), we induced PH in both TRFK5-treated male mice and isotype control-treated male mice (figure 4i). We found that the percentage of EOS was significantly lower in lung tissues of TRFK5-treated mice than in isotype control-treated mice (figure 4j, k). Furthermore, TRFK5 treatment resulted in a higher right ventricular systolic pressure and RV/LV+S in PH mice (figure 4l, m). Consistent with these observations, we also noted that vascular muscularisation was more extensive in EOS-depleted PH mice (figure 4n–p), without affecting pulmonary fibrosis (supplementary figure S5c, d). Because PAH is more predominant in females, we also examined the role of EOS in female mice by TRFK5 injection (supplementary figure S6a). Consistent with the observations in male mice, more severe PH phenotypes were observed in female SuHx-induced PH mice with EOS depletion than in isotype control-treated mice (supplementary figure S6b–g).
Because the rat PH model exhibited more severe progress than the mouse PH model, and shared more similarity with human PAH, we further explored the role of EOS in the SuHx-induced rat PH model. Male rats that received TRFK5 had a decreased blood EOS percentage (supplementary figure S7a, b) compared with the isotype control-treated mice. In the SuHx-induced rat PH model, EOS depletion resulted in more severe PH development and vascular muscularisation, which is consistent with observations in the mouse model (supplementary figure S7c–g).
Taken together, these results show that genetically or chemically induced EOS deficiency aggravates PH development, suggesting that EOS have a protective effect in PH development.
EOS deficiency leads to increased neutrophil and monocyte/macrophage infiltration during PHWe next sought to precisely define the effects of EOS in PH development. Because previous studies have shown that EOS participate in the regulation of immune homeostasis processes, such as Th2 polarisation [20], mast cell infiltration [17] and B-cell immunoglobulin production [18, 19], we examined how EOS deficiency affects different immune cell populations during PH in mice. Using flow cytometry-based assays to quantify the infiltration of different immune cells in ΔdblGATA mice, we observed no significant changes in the percentages of B-cells, T-cells (helper T-cells and cytotoxic T-cells), mast cells or dendritic cells compared with those in WT mice (supplementary figure S8a–e) under PH. ELISA-based detection of immunoglobulin subtype levels indicated that EOS deficiency did not affect IgE, IgG or IgA production in lung tissues of PH mice (supplementary figure S8f–h).
Interestingly, we found increased proportions of CD45+CD11b+Ly6G+ neutrophils and CD45+CD11b+Ly6C+ monocytes/macrophages in lung tissues of EOS-deficient PH mice (figure 5a, b). Immunofluorescent staining further showed more Ly6G+ neutrophils and CD68+ monocytes/macrophages located around remodelled pulmonary vessels in lung tissues of EOS-deficient ΔdblGATA mice under PH (figure 5c, d). EOS depletion in TRFK5-treated mice also led to increased infiltration of neutrophils and monocytes/macrophages into lung tissues compared with that in the isotype control-treated mice with induced PH (figure 5e–h, supplementary figure S6h, i), without affecting other immune cell types (supplementary figure S9).
These observations of enhanced neutrophil and monocyte/macrophage infiltration indicate that EOS deficiency or depletion aggravates the PH-related inflammation of lung tissue in mice.
EOS suppress PASMC proliferation and migrationIncreased pulmonary vascular muscularisation in ΔdblGATA mice during PH suggests a role for EOS in sustaining smooth muscle cells. To test this hypothesis, bone marrow-derived EOS were cultured, and the lysates and supernatants of harvested cells were used to treat PASMCs (figure 6a). After 14 days of culture, the percentage of CD45+CD11b+SiglecF+ EOS in the total cell population reached 95% (figure 6b). PDGFbb-induced PASMC proliferation and migration were effectively suppressed by EOS lysates (figure 6c, d, quantified in 6g) and culture supernatants (figure 6e, f, quantified in 6h) in a dose-dependent manner. These observations suggest a beneficial role of EOS in regulating PASMC function.

留言 (0)