
記住我


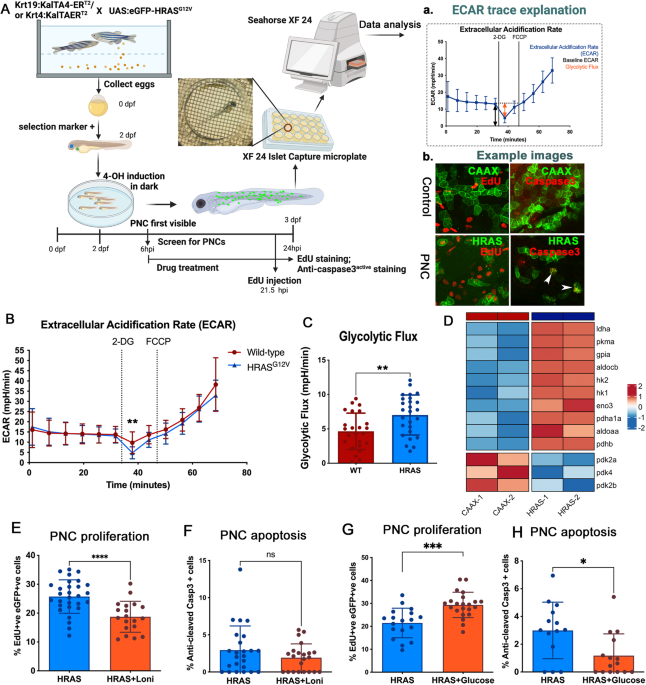
LMNA::NTRK1 fusion is the most common genetic event driving mesenchymal tumors with a lipofibromatosis-like neural phenotype [3, 6]. The LMNA and NTRK1 genes are located on 1q21.2 locus (0.7 Mb apart) with the same direction of transcription. Based on prior targeted RNA/DNA sequencing and FISH studies, it was suggested that the leading mechanism of LMNA::NTRK1 fusion in sarcomas is through a 0.7 Mb interstitial deletion (ID) [6, 7], rather than a reciprocal t(1;1) translocation. We first applied a similar strategy designed for selection of reciprocal inter-chromosomal translocations [8]. Briefly, a donor template with a splice acceptor (SA) site was used resulting in the expression of a selectable marker (hygromycin or puromycin) from the LMNA promoter upon correct integration (Fig. 1A). This gene trap strategy allows the expression of the selectable marker while blocking the fusion expression that can be conditionally induced by Cre recombinase-mediated removal of the marker. Integration of the donor template is promoted by induction of double strand breaks (DSBs) in the participating loci. Relevant to the most common fusion transcript reported in human tumors [7, 9,10,11], we designed sgRNAs to induce DSBs in intron 2–3 of LMNA and intron 9-10 of NTRK1, adjacent to the sequences homologous to the donor (shaded boxes), to generate the LMNA (exon2)-NTRK1 (exon10) isoform (Fig. 1A). The LMNA intron 2–3 and the NTRK1 intron 9-10 sequences were screened for the presence of interspersed repeats and low complexity DNA sequences that could impair targeting efficiency (www.repeatmasker.org), and the sgRNAs were selected using the Guidescan website [12].
Fig. 1: Generation of hES cell lines with conditional LMNA::NTRK1 fusion expression.
A Strategy for conditional LMNA::NTRK1 fusion gene expression. A donor template with homology arms (shaded boxes) and a promoter-less selectable marker (hygromycin or puromycin) is introduced at the DSB sites (scissors) by HDR. A splice acceptor (SA) sequence allows the expression of the marker from the LMNA promoter. Induction of the fusion occurs after removal of the selectable marker by expression of the CRE recombinase. B Dual color FISH analysis showing the NTRK1 locus and the rearranged allele in clones 1 (hygromycin positive; red, centromeric; green, telomeric) and 3 (puromycin positive; green, centromeric; red, telomeric) after loss of the centromeric probe either by t(1;1) or ID. C RT-PCR analysis in hES cells showing the LMNA::NTRK1 fusion level over a period of 30 days after expression of the CRE recombinase in clones 1 and 3. D Dual color FISH analysis on clone 13, (from NHEJ strategy, see S1D) showing loss of 5′ NTRK1 probe (red, centromeric) and RT-PCR confirming the presence of the LMNA::NTRK1 fusion in two distinct clones. E Western blot analysis showing the LMNA::TRKA fusion protein in clones isolated after removal of the marker (clones 1.14 and 3.3) and clone 13. Phosphorylation of TRKA is not detected.
Generating LMNA::NTRK1 fusions in hES cells at improved efficiencySelection of hygromycin positive clones proved to be inefficient compared to prior targeting experiments modeling other sarcoma-related gene fusions in hES cells [13], with <10 colonies/nucleofection of irregular shape and reduced plating efficiency after the first passage. Out of the surviving colonies only one clone (clone 1) showed correct integration of the donor template by PCR analysis. Fluorescence In Situ Hybridization (FISH) assays using custom BAC probes flanking both NTRK1 and LMNA genes (centromeric, red probe; telomeric, green probe) showed a signal pattern consistent with a t(1;1) translocation rather than an ID (Figs. 1B and S1A). The presence of the reciprocal chromosome was confirmed by PCR and sequencing of the breakpoint junction between LMNA intron 2–3 and NTRK1 intron 9–10 (Fig. S1B).
The donor template was modified to include a puromycin selectable marker and a longer LMNA homology arm (461 bp instead of 372 bp). After 10-day selection, 175 colonies were recovered, with 11 clones showing correct integration of the donor template at both participating loci. The puromycin resistant colonies revealed a uniform, rounded shape, and grew after several passages. One clone (clone 3) analyzed by FISH showed an unbalanced rearrangement in keeping with an ID. However, it is possible that the event occurred through a translocation involving multiple DSBs, leading to the imbalance (Fig. 1B). Moreover, the junction of the reciprocal translocation could not be detected, in contrast to the t(1;1) translocation-associated clone 1 (Fig. S1B).
LMNA::NTRK1 fusion resulting from either t(1;1) translocation or interstitial deletion (ID) does not affect hES cell proliferation and viabilityIn contrast to what was observed in hES cells expressing EWSR1-CREB fusions [13], LMNA::NTRK1 transcript levels in both clones (clones 1 and 3) remained stable over a 30-day time course in a pool of cells after Cre recombinase (Fig. 1C). This result suggests that the LMNA::NTRK1 fusion does not impair cell proliferation and/or viability of hES cells, unlike EWSR1-CREB fusions. We then isolated clones that constitutively express the LMNA::NTRK1 fusion after removal of the selectable marker by Cre recombinase. Two clones from each cell line were tested showing comparable LMNA::NTRK1 fusion transcript levels by RT-PCR and qRT-PCR (Fig. S1C). One clone from each original cell line (1.14 from clone 1 and 3.3 from clone 3) was selected for further characterization.
LMNA::NTRK1 fusions by nonhomologous end joining (NHEJ) repair of DSBsGiven the viability of cells expressing the fusion, we also attempted to directly model the fusion by generating two DSBs in the intronic regions of LMNA and NTRK1 in the absence of a homologous donor to allow repair of DNA ends by NHEJ (Fig. S1D). PCR screening by breakpoint junction amplification confirmed the presence of deletions in two clones from a 96-well plate (Fig. S1D). FISH analysis showed the same pattern as observed for clone 3, with loss of the centromeric portion of the NTRK1 gene, and RT-PCR showed similar expression of the LMNA::NTRK1 fusion in both clones (Fig. 1D). One clone (clone 13) was selected for comparison with clones 1.14 and 3.3 from the previous targeting strategies.
Induction of the LMNA::NTRK1 fusion in Retinal Pigment Epithelium (RPE) cell lineAs the LMNA::NTRK1 fusion has been identified in different tumor types including carcinoma, we modeled the fusion in epithelial cells and evaluated the effect on cell proliferation/viability. Retinal Pigment Epithelium (RPE) cells were transfected with sgRNA for LMNA and NTRK1 and the fusion transcript resulting from NHEJ repair was monitored in a time course experiment in a pool of transfected cells. The fusion product was detected up to 18 days, before reduction at day 21 and disappearance by day 25 (Fig. S1E). We also attempted single-cell plating from transfected cells to select single clones expressing the fusion but none of the 112 clones analyzed by PCR showed amplification at the breakpoint junction. These results suggest that the LMNA::NTRK1 fusion affects differently the cell proliferation and/or viability depending on the cellular background.
Detection of the LMNA::TRKA fusion protein in hES clonesWestern blot analysis using an antibody against the C-terminal, catalytic portion of TRKA protein confirmed the presence of the oncogenic fusion protein in clones expressing the fusion transcript (clones 1.14, 3.3 and 13), but not in the parental clones not treated with Cre recombinase (clones 1 and 3) (Fig. 1E). The TRKA kinase domain encompasses exon 12 to 15 with exon 13 coding for the ATP-binding pocket and exon 14 for the activation loop. While we confirmed by Sanger sequencing that the exons are included in the fusion transcripts of the clones 1.14, 3.3 and 13 (Fig. S2), western blot analysis using phospho-TRKA antibodies shows that the TRKA residues Tyr680/681, typically activated in fusion positive tumors, were not phosphorylated in hES cells (Fig. 1E).
LMNA::NTRK1 fusion expression and protein localization in hES-cell derived mesenchymal progenitorsThe use of hES-MP to model cancer relevant translocations and to study the role of fusions in oncogenesis is hampered by their limited passages. We surmounted these limitations by using hES cells that can grow almost indefinitely, form colonies from single cells and differentiate into multiple cell lines. To functionally characterize the role of the LMNA::NTRK1 fusion in tumor development, hES cell models were differentiated to mesenchymal progenitors (hES-MP), the putative cell of origin of different mesenchymal tumor entities expressing this fusion. Cre recombinase was expressed in hES-MP by either transfection or infection, and fusion transcript level was monitored for 30 days. As with hES cells, the LMNA::NTRK1 transcript levels were constant over time in hES-MP cells, without perturbation of cell proliferation and with the NTRK1 portion expressed from the fusion transcript and not from the endogenous allele (Fig. 2A). However, distinct from what was observed in hES cells, phosphorylation of TRKA was detected in these differentiated mesenchymal progenitors (Fig. 2B), emphasizing the importance of the appropriate cellular background when modeling cancer relevant alterations. Immunohistochemistry experiments demonstrated that the cytoplasmic/perinuclear positivity of the LMNA::TRKA fusion protein in a variety of tumor types, including carcinoma and sarcoma, is an effective diagnostic tool [14, 15]. The immunohistochemistry with pan-TRK antibodies on hES-MP cells expressing the LMNA::NTRK1 fusion (clone 3.3) confirmed the cytoplasmic/perinuclear localization of the fusion protein (Fig. S3A).
Fig. 2: Expression of LMNA::NTRK1 fusion in human mesenchymal progenitors (hES-MP).
A Diagram of LMNA::NTRK1 transcript with primers position for RT-PCR. Time course (days) experiment for LMNA::NTRK1 fusion and NTRK1 expression in hES-MP (clones 1 and 3) after Cre expression. B Western blot showing LMNA::TRKA product and phosphorylation of TRKA in hES-MP cells expressing the fusion transcript. C Histogram plot from RNAseq analysis showing NTRK1, NESTIN, S100A and SOX10 RNA levels in hES and hES-MP expressing the LMNA::NTRK1 fusion compared to wild-type isogenic cell line and colorectal cancer cell line KM12 expressing the TPM3::NTRK1 fusion. D GSEA analysis for cell lines expressing the LMNA::NTRK1 fusion (clones 13 and 1.14 (hES-MP) and 13 (hES)). GSEA shows an enrichment of pathways involved in Ewing sarcoma and soft tissue sarcoma.
LMNA::NTRK1-expression in hES-MP cells drives an NTRK1-transcriptional profile with upregulation of neuronal and neuroectodermal signaturesWhole transcriptome sequencing (RNAseq) was performed on clones 1.14 and 13 in both the hES and hES-MP backgrounds. Individual comparisons between the LMNA::NTRK1 positive clones versus wild-type hES and hES-MP cell lines and KM12 colorectal cancer cell line expressing the TPM3::NTRK1 fusion demonstrated differentially expressed genes (DEG) using log2 fold change (FC) > 2 (Fig. 2C). For example, NTRK1 mRNA, presumably from the fusion transcript, was highly upregulated in LMNA::NTRK1 hES-MP compared to wild-type hES-MP cells or to the hES background (Fig. 2C), which was confirmed by RT-PCR for the LMNA::NTRK1 fusion in clones 1.14, 3.3, and 13 (Fig. S3B). Similar to the consistent immunoprofile of LMNA::NTRK1 driven human mesenchymal tumors [3, 6, 16], clone 1.14, although not clone 13, showed mRNA upregulation of S100 but no expression of SOX10 (Fig. 2C). Moreover, NESTIN mRNA levels were upregulated in clones 1.14 and less so in clone 13 compared to wild-type hES-MP cells (Fig. 2C), similar to what has been previously reported in NTRK-fusion positive tumors [7].
The individual DEG and their log2FC from each comparison described above were then subjected to gene set enrichment analysis (GSEA) using Bioconductor cluster Profiler to identify significant pathways. The upregulated genes in hES-MP cells expressing the LMNA::NTRK1 fusion (clone 13 and 1.14) were found to be enriched among the statistically significant pathways involved in neuronal development, neural crest stem cells, Ewing sarcomas and other soft tissue sarcomas (Fig. 2D and Supplementary Table S1).
Modeling the canonical ETV6::NTRK3 gene fusion in hES and hES-MP cellsThe ETV6::NTRK3 fusion represents the canonical fusion (90% frequency) for both infantile fibrosarcoma and mammary analog secretory carcinoma, while being detected at a much lower frequency (5–10%) in a variety of other cancers of various lineages [17]. The ETV6::NTRK3 fusion results from a balanced t(12;15)(p13;q25) translocation, encompassing the entire kinase domain of TRKC expressed by exons 15–18 of NTRK3, with the ATP-binding pocket on exon 16 and the activation loop on exon 17 (Figs. 3A and S2). Employing a similar methodology used to model the desmoplastic small round cell tumor EWSR1::WT1 fusion [8], we aimed to fuse ETV6 exons 1–5 to NTRK3 exons 15-19 in hES cells (Fig. 3B). Briefly, a donor template with homology arms was transfected into hES cells and integrated by HDR at the participating loci. The donor template carries a puromycin selectable marker that is expressed under the control of a PGK promoter and can be excised by Cre recombinase. From the 56 puromycin resistant clones, the PCR analysis showed one clone harboring the translocation (clone G2), which was confirmed by the break-apart FISH analysis (Fig. 3C). The reciprocal translocation (der 12) was not detected in this clone. Cells were then differentiated to hES-MP progenitors, and the fusion transcript levels were monitored in both cell types, remaining stable for a 30-day period (Fig. 3D). Of note, the fusion transcript was detectable in both cell types before Cre expression at consistently lower levels compared to post-Cre cells, likely due to a partial splicing to delete the selectable marker [8]. After Cre expression, hES clones that had removed puromycin marker were isolated (clone G2-6) and differentiated to hES-MP. Immunohistochemistry with pan-TRK antibodies confirmed the nuclear localization of the ETV6::TRKC fusion protein observed in tumor samples [14, 15] (Fig. S3C). Comparison of the fusion transcript levels between the two cellular backgrounds (hES and hES-MP) showed a higher level of ETV6::NTRK3 fusion in mesenchymal progenitors (Fig. S3D), as for LMNA::NTRK1. The induction of the ETV6::TRKC protein and the phosphorylation of the TRKC portion were detected by western blot in hES-MP from clone G2-6 and G2 cells infected with Cre recombinase (Fig. 3E).
Fig. 3: Generation of the ETV6::NTRK3 chromosomal translocation in hES cells and differentiation to hES-MP.
A Breakpoints within ETV6 on chromosome 12 and NTRK3 on chromosome 15 create the ETV6::NTRK3 t(12;15)(p13;q25). B A donor template containing a puromycin selectable marker under the control of a PGK promoter is inserted by HDR at the ETV6 and NTRK3 participating loci after induction of DSBs by sgRNA (scissors). The selectable marker is removed by Cre recombinase allowing the expression of the oncogenic fusion. C Dual color FISH analysis with telomeric (red) and centromeric (green) probes on ETV6 showing the ETV6::NTRK3 translocation on clone G2. D RT-PCR time course (days) for ETV6::NTRK3 fusion after Cre expression in hES and hES-MP cells. The fusion product is detectable in cells before Cre recombination indicating a partial splicing of PGK-Puro sequence. E Western blot analysis showing the ETV6::TRKC fusion protein and the TRKC phosphorylation in hES-MP clone G2-6 and G2 after expression of Cre recombinase. The total ETV6::TRKC protein was detected by pan-TRK antibodies and the TRKC phosphorylation by phospho TRKA/B antibodies (same used for detection of LMNA::pTRKA).
Inhibition of TRK-phosphorylation by pan-TRK inhibitorsTo confirm that the isogenic hES-MP cells expressing the LMNA::NTRK1 and ETV6::NTRK3 fusion are indeed a bona fide model for human tumors driven by the oncogenic activation of the chimeric protein, we monitored TRK phosphorylation in the clones expressing the fusion after treatment with two TRK inhibitors currently used in clinical practice, Entrectinib and Larotrectinib. LMNA::NTRK1 clone 1.14 was initially exposed to increasing concentrations of both inhibitors. Entrectinib was found to reduce phosphorylation at the lowest dose tested, 5 nM, and Larotrectinib at 100 nM (Fig. S3E). Two concentrations were then chosen for each inhibitor, and cells from two LMNA::NTRK1 clones (3.3 and 1.14) and the ETV6::NTRK3 clone G2-6 were treated for 24, 48 and 72 h in a time course experiment. TRKA phosphorylation was reduced in all the conditions tested, while the total fusion protein remained stable (Fig. 4A). Conversely, the stability of ETV6::TRKC was impaired after inhibitor treatment, together with the TRKC phosphorylation [18] (Fig. 4B). To demonstrate the activity of the TRKA-mediated signaling cascade, we tested the phosphorylation of the two well-known downstream effectors ERK1 and ERK2. The phosphorylation of ERK1/2 is constitutively induced in the clone 3.3 expressing the LMNA::TRKA fusion and is reduced to the level of the isogenic clone 3 after a short treatment (1 and 4 h) with Entrectinib and Larotrectinib (Fig. S3F).
Fig. 4: Response to Entrectinib and Larotrectinib inhibition of hES-MP cells expressing LMNA::TRKA and ETV6::TRKC fusion proteins.
A Western blot analysis showing the LMNA-TRKA fusion protein and the loss of TRKA phosphorylation in hES-MP clones 1.14 and 3.3 after 24, 48, and 72 h treatment with the TRK inhibitors Entrectinib (0, 20, and 200 nM) and Larotrectinib (0, 200, and 1000 nM). B Reduced ETV6::TRKC protein stability and TRKC phosphorylation in hES-MP clone G2-6 at different doses of Entrectinib and Larotrectinib.
We measured apoptosis after 72 h treatment with Entrectinib and Larotrectinib and we found that Annexin V positive cells increased in all cell lines tested (Fig. 5A). Finally, cells expressing the LMNA::NTRK1 fusion (clones 1.14 and 3.3) were exposed for 7 days continuously to increasing concentration of both inhibitors showing a dose dependent increase in sensitivity when compared to control isogenic cell lines (clone 1 and 3) (Figs. 5B and S3G).
Fig. 5: Induction of apoptosis and viability after Entrectinib and Larotrectinib treatment in hES-MP cells expressing LMNA::TRKA and ETV6::TRKC fusion proteins.
A hES-MP cells from clones 1 and 1.14, 3 and 3.3 and clone G2 before and after expression of Cre recombinase were exposed for 72 h to two doses of Entrectinib (20 and 200 nM) and Larotrectinib (200 and 1000 nM) and Annexin V was quantified as a measure of apoptosis. Annexin V intensity is represented as the ratio of the percentage of untreated cells. Error bars represent standard deviation from the mean of n ≥ 3 independent experiments. Unpaired t test *p < 0.05, **p < 0.01, ***p < 0.001, if not indicated the difference is not statistically significant. B Cells expressing the LMNA::TRKA protein (clone 1.14) and the corresponding parental cells (clone 1) were treated at increasing concentration of Entrectinib and Larotrectinib for 7 days continuously, before determination of cell viability. Results are presented as the mean of three independent experiments.
留言 (0)