
記住我



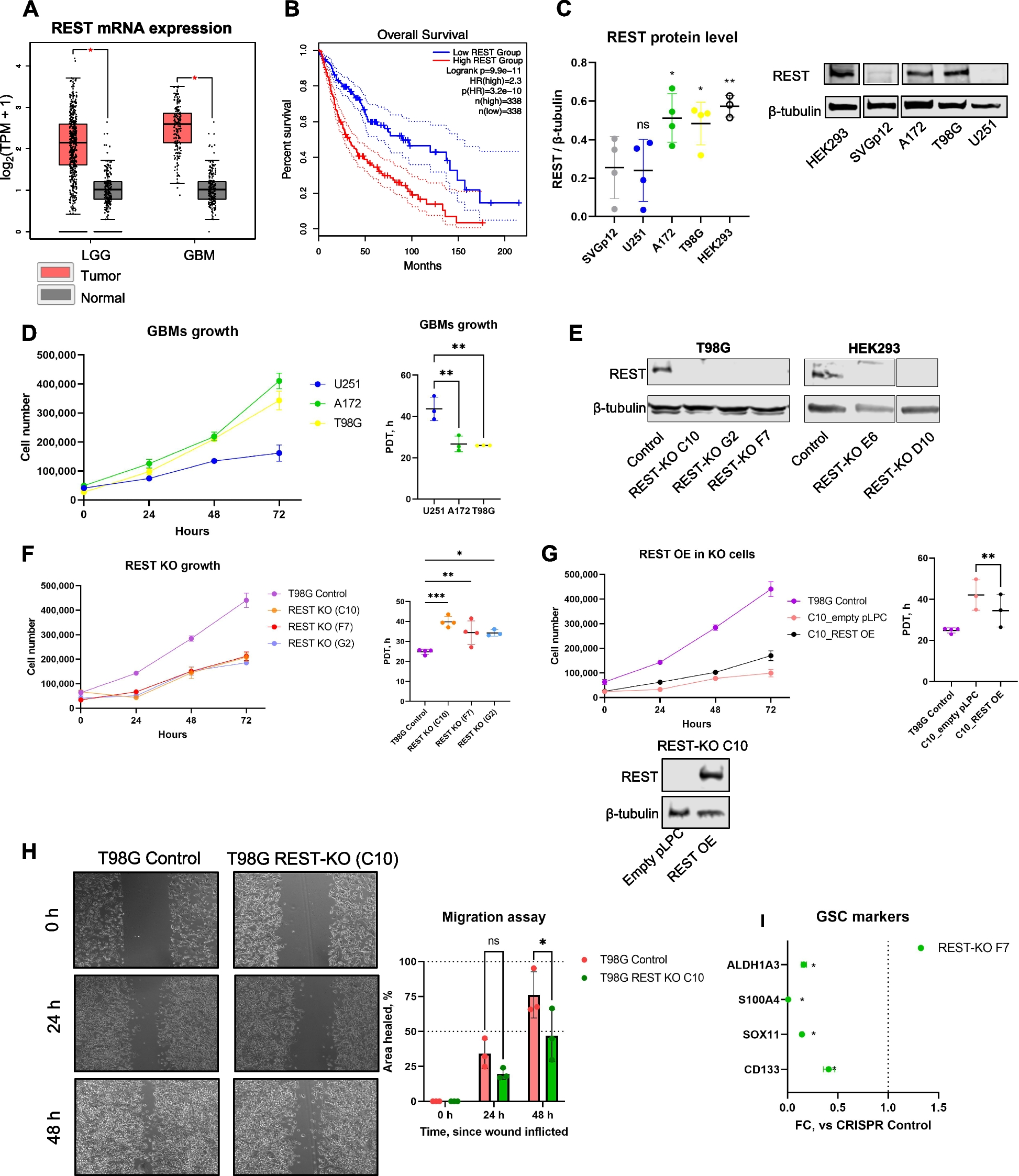
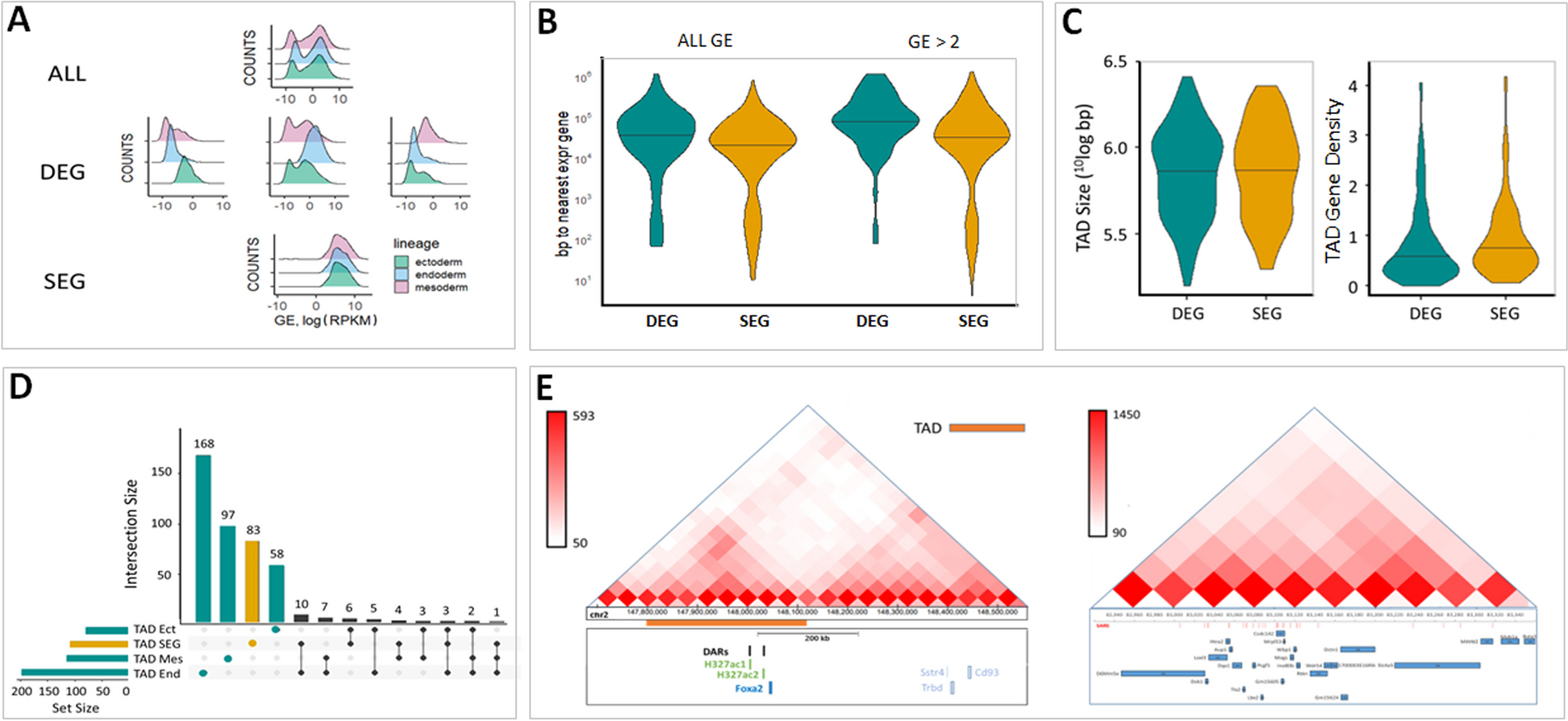
Prior to the detection of erythrocytes, lipid profiling of sera from 3 healthy controls and 4 SCA patients was performed. Compared with healthy subjects, SCA patients had lower concentrations of three major lipids including total cholesterol (104.2 ± 9.7 mg/dL for SCA vs. 164.6 ± 10.8 mg/dL for controls), phospholipids (221.0 ± 22.5 mg/dL for SCA vs. 302.1 ± 62.0 mg/dL for controls), and triglyceride (65.0 ± 22.0 mg/dL for SCA vs. 76.7 ± 12.9 mg/dL for controls) although the latter two did not reach a statistical significance (Fig. 1A).
Fig. 1
Lipid profiles, the masses of apoA-I and apoB in serum of sickle cell anemia (SCA) patients compared with healthy human subjects (the controls). A Lipid profiles of sera including total cholesterol, phospholipid, and triglyceride. B Lipid profiles of lipoproteins (non-HDL and HDL) isolated from sera via FPLC. C Representative western blots of ApoA-I and ApoB (The molecular weights of human ApoA-I and ApoB-100 are ~28.3 kDa and ~510 kDa, respectively). D Quantitative analyses of the western blot data. In A and D, n = 3 for healthy subjects and n = 4 for SCA patients. Statistical significance is indicated as follows: ***P < 0.001
Lipids in human blood are delivered mainly via plasma lipoproteins including ApoA-I-containing lipoproteins (e.g., high-density lipoprotein or HDL) and ApoB-containing lipoproteins (e.g., non-HDL including very low-density lipoprotein or VLDL, low-density lipoprotein or LDL, etc.). To detect the lipid profiles in plasma lipoproteins, aliquots of sera from 3 healthy controls or 4 SCA patients were mixed, and approximately 36 fractions including HDL and non-HDL lipoproteins were separated by a fast protein liquid chromatography (FPLC) which were subjected to lipid profiling via fluorescence spectrometry. The data show that the three lipids (total cholesterol, phospholipids, and triglyceride) in both HDL and particularly non-HDL lipoproteins of SCA patients decreased in comparison with those in the healthy controls (Fig. 1B).
Interestingly, The distribution of phospholipid contents in HDL and non-HDL lipoproteins changed. In the healthy controls, the phospholipid content in HDL is slightly lower than that in non-HDL (the gray curve in the middle panel of Fig. 1B); in the SCA patients, however, the phospholipid level in HDL is slightly higher than that in non-HDL (the black curve in the middle panel of Fig. 1B).
Lower ApoB level whereas similar ApoA-I level in sera of SCA patients compared with the controlsThen, the masses of the major apolipoproteins constructing HDL and non-HDL lipoproteins were evaluated via western blotting. Both the representative western blot bands (Fig. 1C) and the quantitative analyses (Fig. 1D) of ApoA-I and ApoB masses in sera of 3 healthy controls and 4 SCA patients show that ApoB level in SCA patients was much lower, although not statistically significant due to small sample size (3–4 samples), than that in healthy controls (p = 0.0731 in the left panel of Fig. 1D; a patient/control ratio of 0.723 ± 0.276 in the right panel of Fig. 1D) whereas ApoA-I level was similar (p = 0.8586 in left panel of Fig. 1D; a patient/control ratio of 0.997 ± 0.129 in the right panel of Fig. 1D).
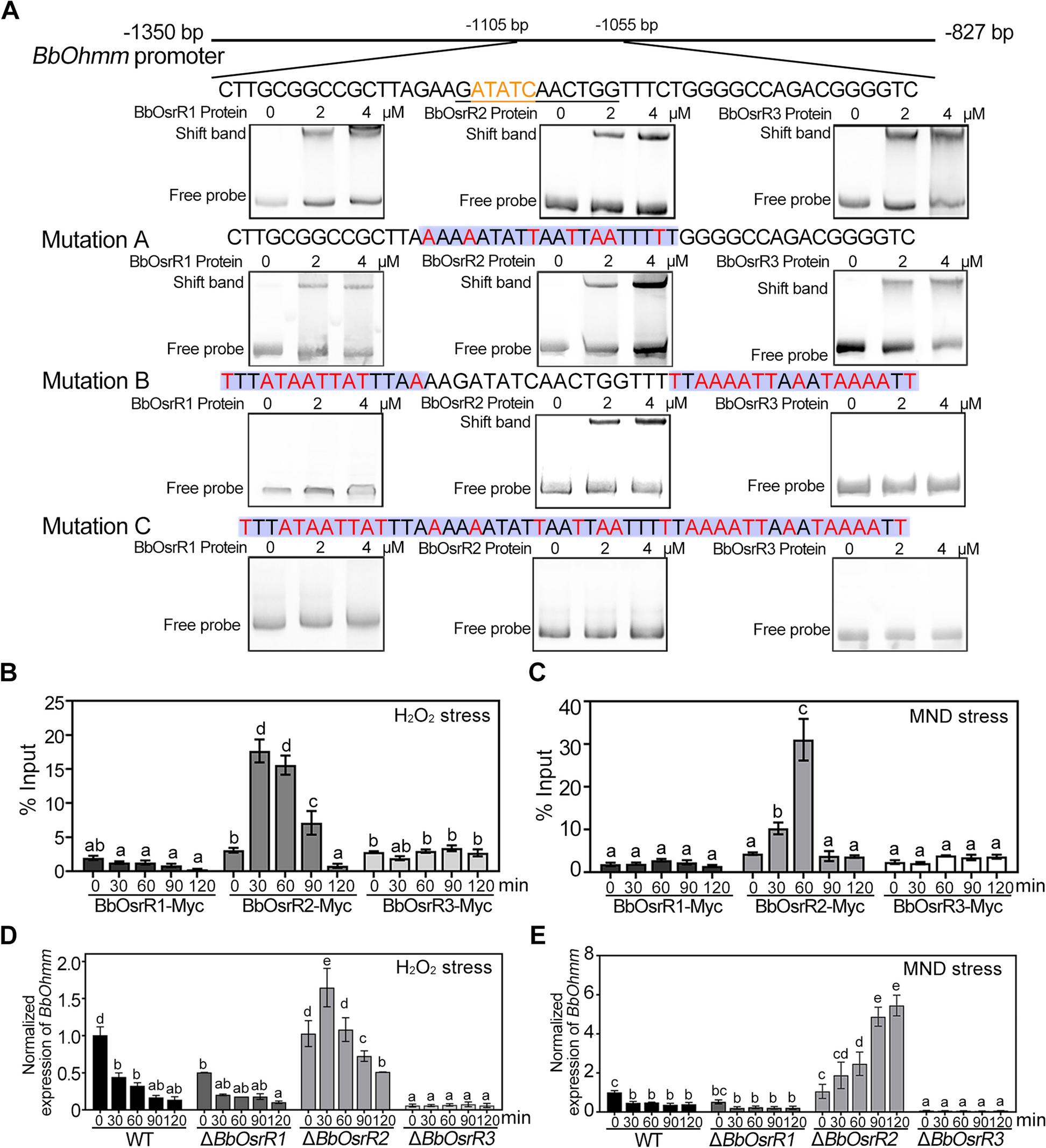
Sickle erythrocytes have a rougher surface than normal erythrocytes detected by AFMNext, red blood cells (RBCs or erythrocytes) isolated from healthy subjects and SCA patients were observed with confocal microscopy prior to the observation by atomic force microscopy (AFM). The typical biconcave erythrocytes and sickle cells were observed in healthy controls and SCA patients, respectively (Fig. 2A). Subsequently, AFM studies of erythrocytes were performed. The typical biconcave erythrocytes and sickle erythrocytes from healthy subjects and SCA patients, respectively, were also observed by AFM (left and middle panels of Fig. 2B, C). The topographical AFM images of local surfaces of individual cells (right panels of Fig. 2B, C) and the height profiles of cross sections across the local surfaces (Fig. 2D) display a relatively rougher surface of sickle erythrocytes compared with normal erythrocytes. The quantitative analysis (Fig. 2E) further confirms the statistically significant increase in surface roughness of sickle erythrocytes (the average roughness is 1.8 ± 0.3 nm and 2.6 ± 0.8 nm for normal and sickle erythrocytes, respectively).
Fig. 2
AFM topographical images of erythrocytes fixed by glutaraldehyde. A Morphology of individual normal erythrocytes (left) and sickle erythrocytes (right) observed by optical microscopy. B AFM images of normal erythrocytes. Left: multiple erythrocytes; middle: single representative erythrocyte; right: local structures on a single normal erythrocyte. C AFM images of sickle erythrocytes from sickle cell anemia patients. Left: multiple erythrocytes; middle: single representative erythrocyte; right: local structures on a single sickle erythrocyte. D High profiles of the cross sections across the local structures on a single normal (left) or sickle (right) erythrocyte. E Roughness quantification of the local structures on single normal or sickle erythrocyte detected by AFM (n = 25; ***P < 0.001)
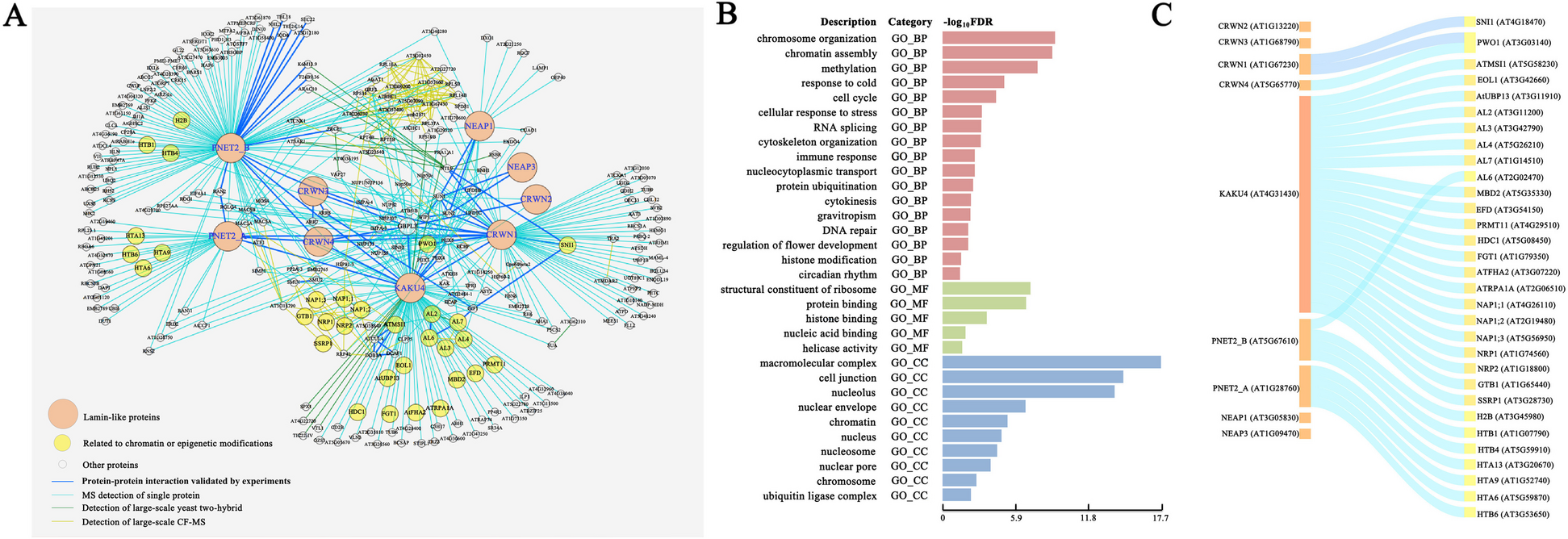
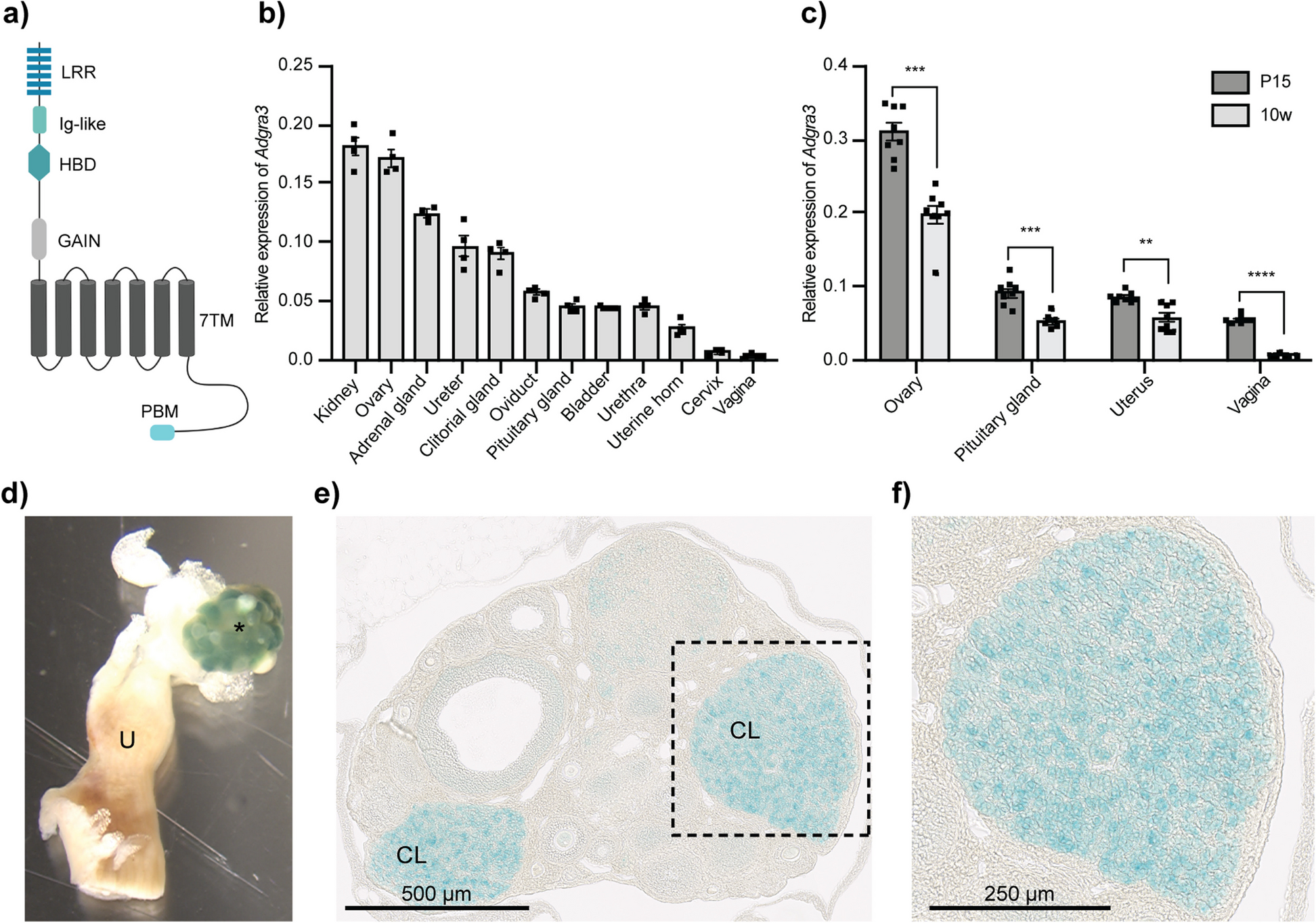
Sickle cell membrane ghosts contain a higher hemoglobin level than normal ghosts detected by SDS-PAGEThe entire cell envelope (generally called “membrane ghosts” or “ghosts”) of erythrocytes can be easily prepared/isolated from erythrocytes via hypotonic treatment and repeated centrifugation for specific studies on the erythrocyte membrane. Prior to AFM detection, the major proteins in isolated membrane ghosts and in the supernatants after centrifugation of membrane ghosts were separated by SDS-PAGE, transferred to nitrocellulose membrane, and visualized after ponceau S staining (Fig. 3A). The supernatants after hypotonic treatment and the first-round centrifugation contained most globin proteins (including monomer, dimer, and tetramer/hemoglobin due to SDS treatment for gel electrophoresis), after the 7th round of centrifugation a few globin monomers still could be detected in the supernatants (lanes 8–18 in Fig. 3A). There were no obvious differences in protein type/content of supernatants between normal and sickle samples. The membrane ghosts (hypotonic treatment and 7 rounds of centrifugation) from all samples contained multiple proteins/bands (some specific proteins including α-/β-spectrin, ankyrin, band 3, globin, and others are indicated in the image based on their molecular weights and the locations of their bands corresponding to the marker bands), among which the ghosts from sickle erythrocytes exhibited a higher level of hemoglobin in comparison with the ghosts from normal erythrocytes (lanes 2–7 in Fig. 3A) which was further confirmed by the quantitative analysis (Fig. 3B).
Fig. 3
Major proteins in isolated membrane ghosts and in supernatants after centrifugation of membrane ghosts of normal and sickle erythrocytes. The samples were separated by SDS-PAGE, transferred to nitrocellulose membrane, and stained with ponceau S (A). Lane 1: protein marker; lanes 2–4: isolated membrane ghosts of the normal erythrocytes from three healthy subjects; lanes 5-7: isolated membrane ghosts of the sickle erythrocytes from three sickle cell anemia patients; lanes 8–12: the supernatants after 1st, 4th, and 7th centrifugation of membrane ghosts of normal erythrocytes from two healthy subjects; lanes 13–18: the supernatants after 1st, 4th, and 7th centrifugation of membrane ghosts of sickle erythrocytes from two patients. B Quantitative analysis of the hemoglobin bands indicated by the white boxes in A (***P < 0.001)
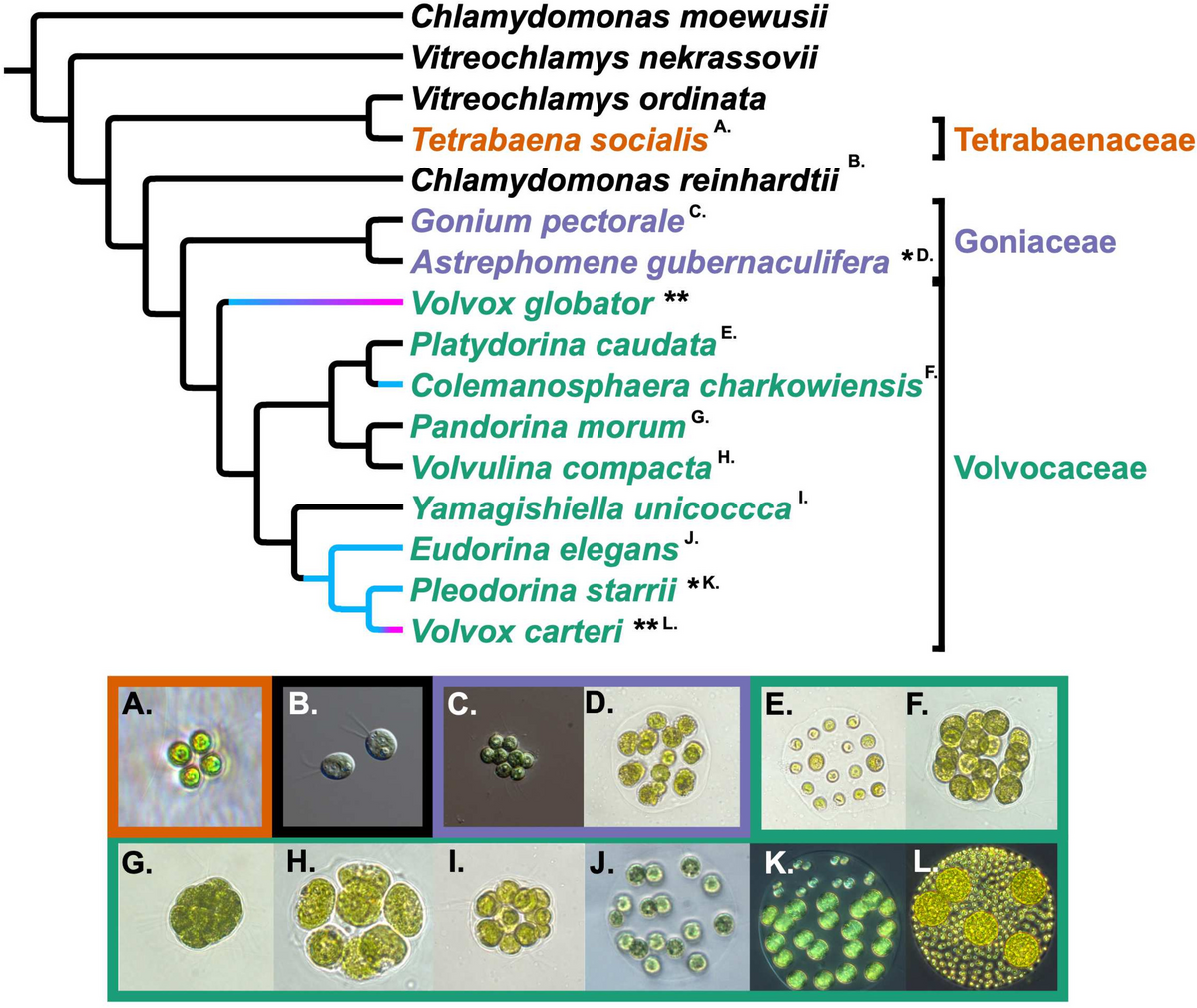
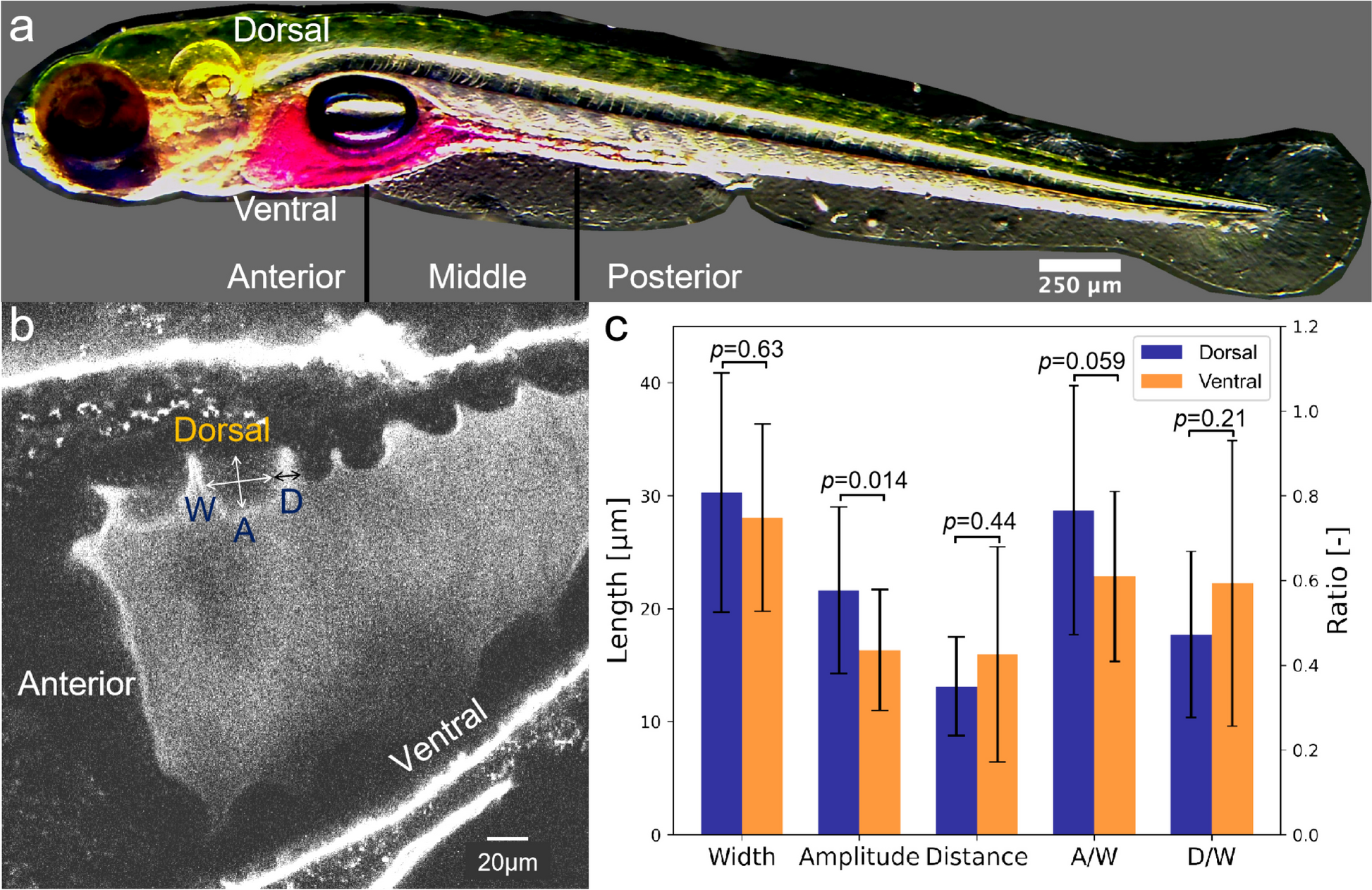
Sickle erythrocyte membrane ghosts are thicker and rougher than normal ghosts detected by AFMReal-time optical observation shows that erythrocytes became transparent after the removal of intracellular hemoglobin (Fig. 4A–C). It was observed that the transformation of most sickle erythrocytes into membrane ghosts lagged behind in comparison with that of normal erythrocytes (Fig. 4A) and some sickle cells temporarily maintained a sickle shape after the removal of intracellular hemoglobin (Fig. 4B). Eventually, however, almost all sickle erythrocytes became membrane ghosts with a wafer-shaped (or circular pie-shaped) morphology similar to those of normal erythrocytes as observed by optical microscope (Fig. 4C) and AFM (Fig. 4D).
Fig. 4
Optical and AFM imaging of entire erythrocyte membrane ghosts. A, B Time-lapse optical observation of the transformation from whole cells (dark) to membrane ghosts (ghosts; transparent) in hypotonic solution. The yellow asterisks indicate the sickle-shaped cells/ghosts. C Optical observation of membrane ghosts from normal (top) and sickle (bottom) erythrocytes. D AFM topographical images of glutaraldehyde-fixed ghosts from normal erythrocytes (left; the inset is enlarged from the white square showing the overlapping of two ghosts indicated by the white asterisks) and sickle erythrocytes (right). E Height profiles of the cross sections of individual ghosts across the dashed lines in C (the black asterisk in D corresponds to the white asterisks in C). F Quantitative analysis of average diameter (left) and average height (right) of ghosts (n = ~50; ***P < 0.001)
The height profiles of the cross sections across multiple entire ghosts in AFM topographical images show that individual ghosts from sickle erythrocytes were higher than those from normal erythrocytes although they have a similar diameter (Fig. 4E; the black and red dashed lines indicate the approximate heights of normal and sickle cell membrane ghosts, respectively). Quantitative analyses find that the average ghost diameters from normal and sickle erythrocytes are 9.09 ± 0.57 μm and 9.54 ± 1.04 μm, respectively and that the average ghost heights from normal and sickle erythrocytes are 21.3 ± 2.8 nm and 26.4 ± 4.0 nm, respectively (Fig. 4F), which actually contain two layers of membrane ghost.
When single membrane ghosts were observed by AFM and local structures were highlighted, it was found that the surface of membrane ghosts from normal erythrocytes (Fig. 5A) seems much smoother than that from sickle erythrocytes (Fig. 5B). It is worth noticing that many small bumps appear on the surface of membrane ghosts from sickle erythrocytes (Fig. 5B). The height profiles of the cross sections across single entire ghosts confirm that compared with normal erythrocytes sickle cells have a membrane ghost with similar diameter, higher thickness (Fig. 5C; the black and red dashed lines indicate the approximate heights of normal and sickle cell ghosts, respectively), and rougher surface. The quantitative analysis (Fig. 5D) reveals the statistically significant increase in average surface roughness of membrane ghosts from sickle erythrocytes (1.9 ± 0.8 nm and 2.9 ± 1.2 nm for the membrane ghosts from normal and sickle erythrocytes, respectively). The average surface roughness values of membrane ghosts are similar to those of entire erythrocytes (Fig. 2E).
Fig. 5
AFM topographical images of local structures of individual erythrocyte membrane ghosts fixed by glutaraldehyde. A A representative membrane ghost of normal erythrocytes. B A representative membrane ghost of erythrocytes from sickle cell anemia patients. From left to right panels: Entire membrane ghosts, local structures (3 μm × 3 μm), and smaller local structures (1 μm × 1 μm), respectively. C Height profiles of the cross sections of individual normal (left) and sickle (right) membrane ghosts. D Quantitative analysis of average roughness of the local surfaces (n = 30; ***P < 0.001)
To visualize the inner surface of membrane ghosts, most of the upper layer of membrane ghosts was peeled by shear force to expose the inner/cytoplasmic surface of the bottom layer of membrane ghosts remaining a small part of the upper layer of membrane ghosts with its outer surface detectable by AFM (Fig. 6A). The height profiles of cross sections (Fig. 6B) show that the heights of the membrane ghost in one layer from normal and sickle cells are approximately 10 nm and 15 nm, respectively. the quantitative analysis (Fig. 6C) shows the average thickness of the bottom layers of membrane ghosts (11.3 ± 1.2 nm for normal erythrocyte membrane vs. 13.9 ± 1.6 nm for sickle cell membrane) which are around half of the heights of the intact membrane ghost in two layers (Fig. 4F). The data confirms that the ghosts in one layer from sickle erythrocytes are thicker than those from normal erythrocytes. Figure 6D displays the representative AFM topographical images of the outer and inner surfaces of membrane ghosts from normal and sickle erythrocytes. It is worth noticing that the inner/cytoplasmic surfaces of both normal and sickle cell ghosts display a meshwork-like structure which probably is based on the spectrin network beneath the plasma membrane. Both the topographical images and the quantitative analysis of average surface roughness (Fig. 6E) show that the inner surface is rougher than the outer surface for both normal and sickle ghosts and that sickle ghosts are rougher than normal ghosts for both outer and inner surfaces.
Fig. 6
AFM topographical images of outer and inner/cytoplasmic surfaces of individual erythrocyte membrane ghosts. A Representative membrane ghosts of normal (left) and sickle (right) erythrocytes. B Height profiles of the cross sections of individual normal (left) and sickle (right) ghosts showing the heights of the inner and outer surfaces which are indicated by the black and red dashed lines, respectively. C Quantification of the average thickness of the bottom layers of membrane ghosts (n = 15; ***P < 0.001). D Representative topographical images showing the local ultrastructures of the outer and inner surfaces of normal and sickle erythrocytes. The meshwork-like structures with a mesh size of less than 200 nm are evident in the inner surfaces of both normal and sickle cell ghosts. E Quantitative analysis of average roughness of the local surfaces (n = 15; **P < 0.01, ***P < 0.001). The marks N and S represent the normal and sickle cell samples, respectively
Cholesterol depletion by MβCD induces surface roughening at relatively low concentration and surface damage or even cell degradation at high concentrations for both normal and sickle erythrocytesMethyl-beta-cyclodextrin (MβCD) is a widely used reagent for depleting the cholesterol in the plasma membrane of cells. Here, the effects of MβCD at different concentrations on the number of erythrocytes and the cholesterol level in the plasma membrane of erythrocytes were evaluated. Clearly, MβCD could cause the degradation of erythrocytes in a concentration-dependent manner and reach a statistically significant decrease at a concentration of larger than 1.6 mM (Fig. 7A). Flow cytometric quantification found that MβCD could induce a concentration-dependent (statistically significant at 0.8 and 1.6 mM) decrease in cholesterol of the plasma membranes of both normal (Fig. 7B) and sickle (Fig. 7C) erythrocytes as well as in cholesterol of membrane ghosts of both normal and sickle erythrocytes (Fig. 7D), confirming the cholesterol-depleting effect of MβCD.
Fig. 7
Effects of MβCD at different concentrations on cell viability and cholesterol depletion. A Effect of MβCD on the number of erythrocytes (n = 6; **P < 0.01). B–D The representative flow cytometric data show the fluorescence intensity of filipin-stained normal erythrocytes (B), sickle erythrocytes (C), and their membrane ghosts (D), respectively, treated with MβCD at indicated concentrations for 1h at 37 °C. Insets show the quantitative analyses (n = 3; **P < 0.01, ****P < 0.0001)
The following phenomena are observed in AFM topographical images of entire erythrocytes treated with MβCD: (a) With the increasing of MβCD concentration (from 0 mM to 12.8 mM) the number of erythrocytes decreased gradually (Fig. 8A); (b) more and more biconcave erythrocyte transformed into spherical erythrocytes (Fig. 8A) whereas most erythrocytes treated with 0.8 mM MβCD still remained in a biconcave shape (panel 2 of Fig. 8A); (c) Large pits (indicated by arrows in Fig. 8B) appeared in the surfaces of many erythrocytes treated by MβCD at a relatively high concentration (e.g., 6.4 mM); (d) 0.8 mM MβCD treatment did induce significant changes in shape of erythrocytes (the length to width ratio: 1.019 ± 0.124 vs. 1.004 ± 0.131 for untreated and MβCD-treated normal cells, respectively and 1.236 ± 0.287 vs. 1.207 ± 0.220 for untreated and MβCD-treated sickle erythrocytes, respectively; n = 50, P > 0.05); (e) 0.8 mM MβCD treatment also caused the increase in cell-surface roughness for both normal and sickle erythrocytes as showed by AFM images of local surfaces of erythrocytes (Fig. 8C) and quantitative analysis (Fig. 8D; the average cell-surface roughness is 1.8 ± 0.3 nm and 2.5 ± 0.6 nm for normal and sickle cells without treatments, respectively, and 2.6 ± 0.8 nm and 3.3 ± 1.1 nm for normal and sickle cells treated with 0.8 mM MβCD, respectively). MβCD-induced cholesterol depletion was responsible for the cell-surface roughening and membrane damage (e.g., the large pits) which further contributed to the shape change and cell degradation probably due to the increase of membrane permeability. Since 0.8 mM MβCD could induce a statistically significant cholesterol depletion but no statistically significant cell degradation and no obvious changes in the entire morphology/shape of erythrocyte, this concentration (0.8 mM) of MβCD was used in the following AFM experiments.
Fig. 8
Effects of MβCD on erythrocytes detected by AFM after glutaraldehyde fixation. A AFM topographical images (80 μm × 80 μm) of normal erythrocytes treated with MβCD at 0, 0.8, 6.4, and 12.8 mM, respectively for 1h at 37 °C (from left to right). B AFM topographical images of entire erythrocytes (40 μm × 40 μm) treated with 6.4 mM MβCD and an enlarged area from the dashed square (3 μm × 3 μm). The arrows indicate the large pits in the surface of erythrocytes. C AFM topographical images of local surfaces (0.5 μm × 0.5 μm) of normal and sickle erythrocytes treated with or without 0.8 mM MβCD for 1h at 37 °C, respectively. Insets display the entire cells (10 μm × 10 μm). D Quantitative analysis of the surface roughness of normal and sickle erythrocytes treated with or without MβCD (n = 25; *P < 0.05, **P < 0.01, ***P < 0.001). The marks N and S represent the normal and sickle cell samples, respectively
Cholesterol depletion by MβCD causes an increase in roughness and a decrease in thickness of membrane ghosts for both normal and sickle erythrocytesAfter isolation from erythrocytes, membrane ghosts were also treated with 0.8 mM MβCD and imaged by AFM (Fig. 9). Compared with the relatively smooth surfaces of membrane ghosts from normal erythrocytes (Fig. 9A), the surfaces with many small bumps could be observed on membrane ghosts of sickle erythrocytes (Fig. 9C) as also showed in Fig. 5B. After 0.8 mM MβCD treatment, many creases could be observed on membrane ghosts from both normal and sickle erythrocytes (as indicated by the arrows in left panels of Fig. 9B, D) implying the local detachment of membrane ghosts from the underlying cytoskeleton. Compared with the membrane ghosts without MβCD treatment (Fig. 9A, C), the membrane ghosts treated with MβCD (right panels of Fig. 9B, D) had a larger roughness which was confirmed by the quantitative analysis (Fig. 9G; the average roughness is 2.8 ± 1.0 nm and 4.6 ± 0.7 nm for normal and sickle cells without treatments, respectively and 4.7 ± 0.9 nm and 5.8 ± 2.2 nm for normal and sickle cells treated with 0.8 mM MβCD, respectively). Moreover, after MβCD treatment, relatively big pits (indicated by the white asterisks) appeared in the membrane ghosts of sickle erythrocytes (right panel in Fig. 9D). On the other hand, MβCD treatment induced a ~3.4-nm reduction in average height of normal cell membrane ghosts (21.3 ± 2.8 nm for non-treatment vs. 17.9 ± 1.9 nm for treatment) and a ~4.0-nm reduction in average height of sickle cell membrane ghosts (26.4 ± 4.0 nm for non-treatment vs. 22.4 ± 4.1 nm for treatment), respectively according to the height profiles of cross sections (Fig. 9E) and the quantitative analysis (Fig. 9F).
Fig. 9
Effect of MβCD on membrane ghosts of normal and sickle erythrocytes (fixed by glutaraldehyde after MβCD treatment). A, B Membrane ghosts from normal erythrocytes. C, D Membrane ghosts from sickle erythrocytes. (A) AFM topographical image of a local surface (2 μm × 2 μm) enlarged from an untreated ghost (inset). B AFM topographical images of the MβCD-treated ghosts (left, ~ 12 μm × 12 μm) and a local surface (right, 2 μm × 2 μm). C AFM topographical image of a local surface (2 μm × 2 μm) enlarged from an untreated ghost (inset). D AFM topographical images of the MβCD-treated ghosts (left, ~ 12 μm × 12 μm) and a local surface (right, 2 μm × 2 μm). E Height profiles of the cross sections across entire ghosts from normal or sickle cells with or without treatment. (MβCD). F Quantitative analysis of average height of ghosts in two layers (n = 50). G Quantitative analysis of surface roughness (Sa) of ghosts (n = 25). The ghosts were treated with 0.8 mM MβCD for 1 h at 37 °C. The white arrows in B and D indicate the creases on ghosts treated by MβCD. The white asterisks in D indicate the large pits with a diameter of ~250–500 nm (231.8 ± 20.4 nm; n = 20 cells) in ghosts treated by MβCD. Statistical significance is indicated as follows:*P < 0.05, **P < 0.01, ***P < 0.001. The marks N and S represent the normal and sickle cell samples, respectively in F and G
Sickle erythrocytes are stiffer than normal erythrocytes and cholesterol depletion by MβCD induces a larger decrease in stiffness of sickle cells than that of normal erythrocytes detected by AFMFinally, AFM was recruited to detect/measure the rigidity/stiffness of both normal and sickle erythrocytes treated with or without MβCD. Due to the potential influence of a fixative on cell stiffness, all erythrocytes for stiffness measurement were not fixed by glutaraldehyde in this experiment. By using the force measurement function (force spectroscopy) of AFM, both the topographical mapping and Young’s modulus mapping were performed on the same single erythrocytes from healthy subjects and sickle cell anemia patients (Fig. 10). Compared with the normal erythrocytes (Fig. 10A), sickle erythrocytes (Fig. 10B) had a much higher average Young’s modulus (6.036 ± 2.577 kPa for sickle cells vs. 2.597 ± 1.221 kPa for normal cells; Fig. 10C) implying that sickle cells are ~2.3-fold stiffer than normal erythrocytes. Upon 0.8 mM MβCD treatment, the average Young’s moduli of both normal and sickle erythrocytes significantly decreased to 1.847 ± 0.909 kPa for normal erythrocytes and 4.456 ± 1.659 kPa for sickle cells, respectively (Fig. 10C).
Fig. 10
AFM detection of the stiffness of individual erythrocytes in PBS buffer. A A representative normal erythrocyte. B A representative sickle erythrocyte. Left: topographical mapping; right: Young’s modulus mapping. C Quantitative analysis of Young’s modulus (n = 100; *P < 0.05, ****P < 0.0001). All erythrocytes in this experiment were not fixed by a fixative
留言 (0)