In this study, we investigated how cortical tau tangles propagate in MTL, inferior, and middle temporal cortices in AD. We found that initial levels of Aβ and tau tangles were linked differently to the longitudinal tau accumulation inside and outside of MTL. Specifically, baseline Aβ plaques and entorhinal tau tangles were related to faster tau accumulation in the amygdala, entorhinal, and parahippocampal gyrus included in the temporal meta-ROI in A+/T− individuals. While greater initial entorhinal tau deposition predicted faster tau accumulations in MTL and further propagations into inferior and middle temporal cortices, higher baseline Aβ plaques were only poorly associated with faster FTP SUVR increases in fusiform, inferior temporal, and middle temporal regions in A+/T+ individuals. Furthermore, we found that early-life elderly adults had faster Aβ-dependent but slower tau-dependent longitudinal tau accumulations in fusiform and inferior temporal areas than late-life elderly adults. Additionally, we found that KL-VShet+ attenuated longitudinal tau accumulations in fusiform and inferior temporal lobe via modulating initial entorhinal tau-associated rather than Aβ-associated tau propagation. These findings are crucial for understanding how tau tangles propagate in the early Braak stages of AD, providing novel insights to design future AD clinical trials.
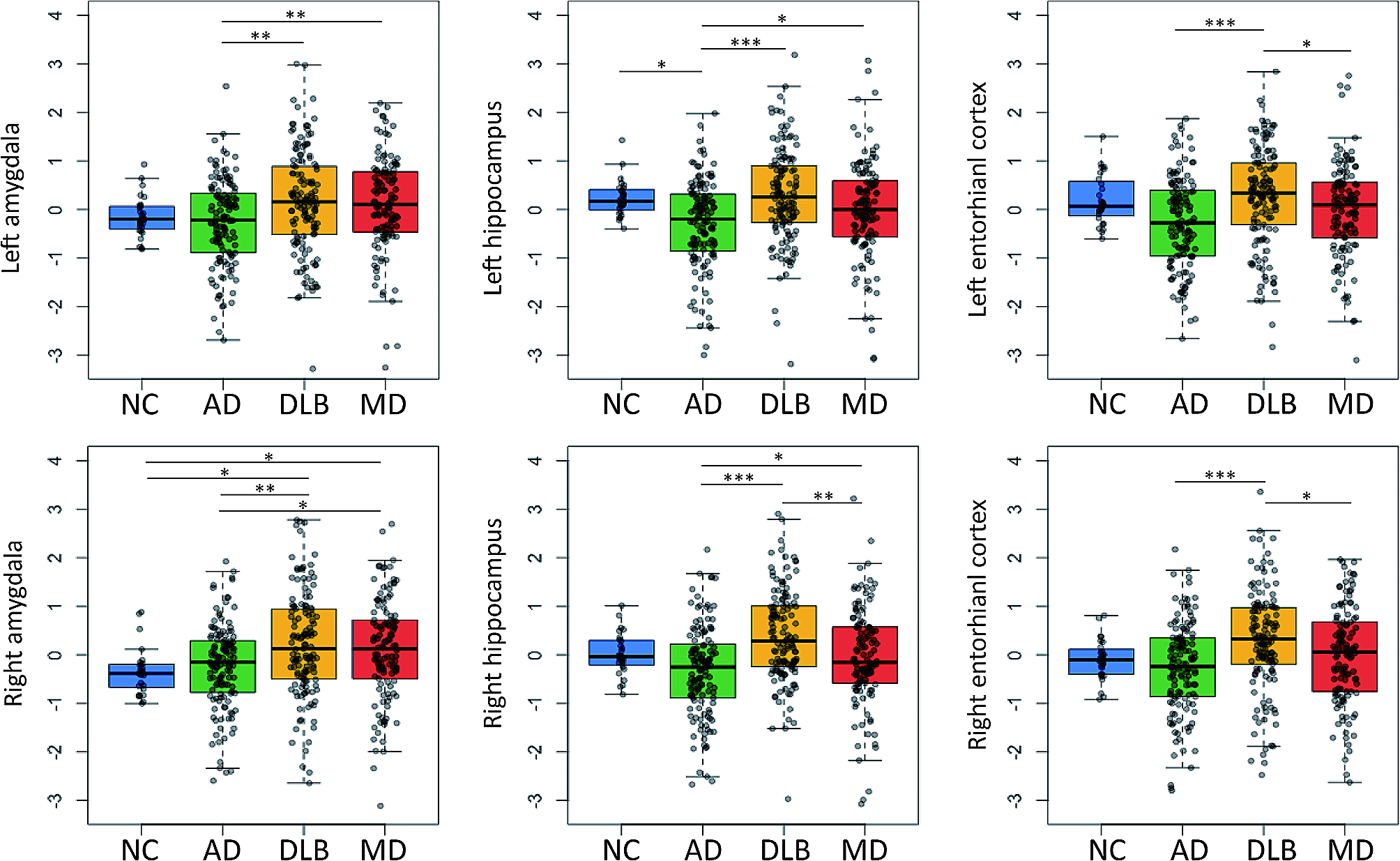
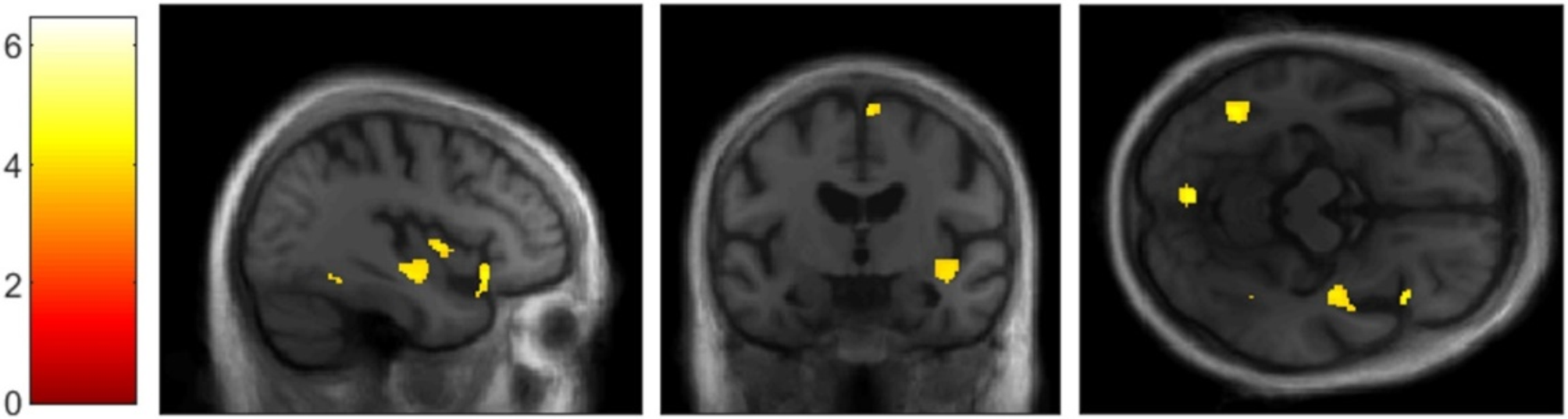
We first determined the early tau-deposited regions in individuals with abnormal AD summary cortical Aβ PET SUVR but normal temporal meta-ROI tau PET SUVR (A+/T−). Focusing on this group, we found that A+/T− individuals had significantly or marginally higher baseline FTP SUVRs and faster rates of FTP SUVR increases in the amygdala, entorhinal, and parahippocampal gyrus compared to A−/T−/N− individuals. The partial correlation analysis further revealed a potential cascading order of tau spread from parahippocampal gyrus to fusiform to inferior temporal to middle temporal cortices. In contrast, the propagating sequence was unambiguous in the entorhinal and amygdala. Early tau deposition likely occurs in the entorhinal and amygdala in the early stages of AD. Consistently, one recent study also observed the most considerable tau deposited in the amygdala in both A−/T+ and A+/T+ individuals [38], and numerous studies [11, 13, 39,40,41] have reported the entorhinal cortex being the earliest site of neurofibrillary tau tangle formation.
The Harvard group [11] recently observed that higher baseline entorhinal tau and Aβ burden would trigger the greatest tau spreading into the inferior temporal region. Another recent study also highlighted the significant interaction of Aβ plaques and tau tangles in promoting the onset and acceleration of longitudinal tau accumulation [23]. Congruently, our findings showed that for A+/T− individuals, both baseline Aβ plaques and entorhinal tau tangles were associated with faster tau accumulation in early tau-deposited ROIs (amygdala, entorhinal, and parahippocampal) of temporal meta-ROI. These findings provide further evidence to explain how initial Aβ burden and regional tau deposition are associated with longitudinal tau accumulation in MTL in Aβ-positive individuals without abnormal temporal meta-ROI tau. Unlike the initial entorhinal tau level, initial amygdala and parahippocampal tau levels in A+/T− individuals only predicted faster tau accumulation in amygdala, and the tau levels in late tau-deposited ROIs (fusiform, inferior temporal, and middle temporal cortices) of temporal meta-ROI did not predict any longitudinal tau accumulation in early tau-deposited regions. These findings suggest that the faster tau accumulations in amygdala, entorhinal, and parahippocampal gyrus are probably associated with existing cortical Aβ plaques and entorhinal tau deposition in the A+/T− stage of AD.
Once individuals had widespread tau tangles in temporal meta-ROI (A+/T+), unlike tau levels in other individual ROIs in temporal meta-ROI, higher initial entorhinal tau level was positively correlated with faster tau accumulations in all individual ROIs. Such findings suggest that entorhinal tau might be a crucial predictor of the subsequent accumulation of tau aggregates in mesial and basal temporal neocortices [11]. Notably, for A+/T+ individuals, the baseline tau levels and longitudinal tau accumulations were strongly linked to each other in the relatively late tau-deposited ROIs (parahippocampal, fusiform, inferior temporal, and middle temporal regions). However, the tau levels in relatively late tau-deposited ROIs would unlikely affect tau accumulations in early tau-deposited regions, whether in A+/T− or A+/T+ groups. The prediction effect of faster tau accumulation in fusiform, inferior temporal, and middle temporal areas by a more considerable Aβ burden became minimal no matter the tau level of which late tau-deposited ROI was included in our models.
Notably, our findings are not suggesting that existing Aβ plaques are unimportant for longitudinal tau accumulation. Recently, Lee and colleagues [23] observed a significant local interaction of Aβ plaques and tau tangles in the inferior temporal region, sparking the most significant acceleration of tauopathy. Differences in modeling methodology might underlie such discrepancies. While the local Aβ and tau interaction (Aβ×tau) within the same region was modeled in their study [23], we used a summary cortical Aβ PET SUVR of several brain areas [32] to represent cortical Aβ burden. The amount of Aβ plaques and regional tau tangles were modeled separately to distinguish their respective contributions in the downstream tau accumulation. Furthermore, many studies reported that abnormal tau tangles outside the entorhinal cortex were rarely observed in the absence of substantial cortical Aβ burden [9, 20, 21, 25, 42], and larger Aβ burden was strongly related to more rapid tau accumulation over time [5, 10, 11, 20]. Altogether, it might be possible that the existing tau tangles contribute more to longitudinal tau accumulations in late tau-deposited ROIs of temporal meta-ROI than cortical Aβ plaques when Aβ plaques and tau tangles are already widespread in neocortex and temporal meta-ROI.
Our findings may have potential implications for AD clinical trials (e.g., anti-Aβ or anti-tau treatments): For Aβ+ individuals without abnormal temporal meta-ROI tau, reducing cortical Aβ plaques might be a promising therapeutic strategy to decelerate the further spreading of tau aggregates, and prevent subsequent tau-associated neurodegeneration and cognitive decline. However, for individuals who already have widespread cortical Aβ plaques and tau tangles, sole Aβ-lowering therapy might be insufficient. Directly targeting the local tau aggregates might be more effective in preventing AD progression.
Recent literature has demonstrated that females have more tau depositions [30] and faster tau accumulations [24, 29] than males. One cross-sectional study reported that the tau tangles of females in the entorhinal and inferior temporal lobe were more positively associated with Aβ burden than those of males [43]. Similarly, our work further manifested that females may exhibit faster initial tau-associated rather than Aβ-associated longitudinal tau increases than males. Both cross-sectional [44] and longitudinal studies [24, 26, 29] have found that larger amounts of Aβ plaques were associated with faster tau accumulation in Aβ+ individuals under 65 years old. Congruently, we also observed more rapid Aβ-associated longitudinal tau increases in the fusiform, inferior temporal, and middle temporal cortices in early-life elderly adults (age < 65 or age < 72.7 years) than in late-life elderly adults (age ≥ 65 or age ≥ 72.7 years). Unexpectedly, participants of older ages showed faster tau-related longitudinal tau increases in parahippocampal, fusiform, inferior temporal, and middle temporal cortices compared to those of younger ages. To the best of our knowledge, this primary finding has not been thoroughly discussed in the literature. In line with our result, a recent animal study found that older animals had more tau spreading in the hippocampus and adjacent cortices than younger ones [45].
Previous findings have found that KL-VShet+ carriers have decreased Aβ and tau pathologies [46]. For instance, in one recent study, KL-VShet+ carriers attenuated Aβ-associated tau increases cross-sectionally and longitudinally [28]. Similarly, for KL-VShet+ carriers, our results exhibited marginally decreased Aβ levels but prominently decreased baseline tau levels and rates of tau accumulation (e.g., KL-VShet+/KL-VShet− ratio as 0.551 in the parahippocampal gyrus). This reduction effect might indicate a notable lowering effect of KL-VShet+ genotyping on subsequent tau accumulations. More specifically, in our current study, we observed both Aβ-associated and entorhinal tau-associated decreased longitudinal tau accumulation in fusiform and inferior temporal cortices in KL-VShet+ carriers, suggesting that KL-VShet+ carriers might slow down both Aβ-associated and tau-associated longitudinal tau accumulation in AD. Furthermore, mediation analyses verified that only the pathway from KL-VShet to initial entorhinal tau, then to longitudinal tau accumulation, was statistically significant when we included baseline levels of Aβ plaques and tau aggregates into the models. This verified pathway demonstrates that the slower rate of tau accumulation in KL-VShet+ carriers was probably mediated via decreased entorhinal tau-associated rather than Aβ-associated pathway. The findings reached by the current study extended our knowledge regarding the potential protective mechanism of KL-VShet+ genotyping and might have implications for the selection of at-risk individuals for clinical trials.
Notably, the influences of sex, age, and KL-VShet on longitudinal tau accumulation are predominantly tau-dependent, and the brain regions involved are mainly located in the late tau-deposited ROIs of temporal meta-ROI. Our preceding findings demonstrated that both Aβ burden and tau tangles act in the early stage of tauopathy, while subsequent accumulation of tau aggregates may strongly associate with baseline levels of tau aggregates at a more advanced stage of AD. Therefore, we infer that sex, age, and KL-VShet mainly modulate the relationship between Aβ burden, tau tangles, and longitudinal tau accumulation at a stage when tau tangles accumulate rapidly and when initial symptoms of AD start to emerge [23, 47, 48].
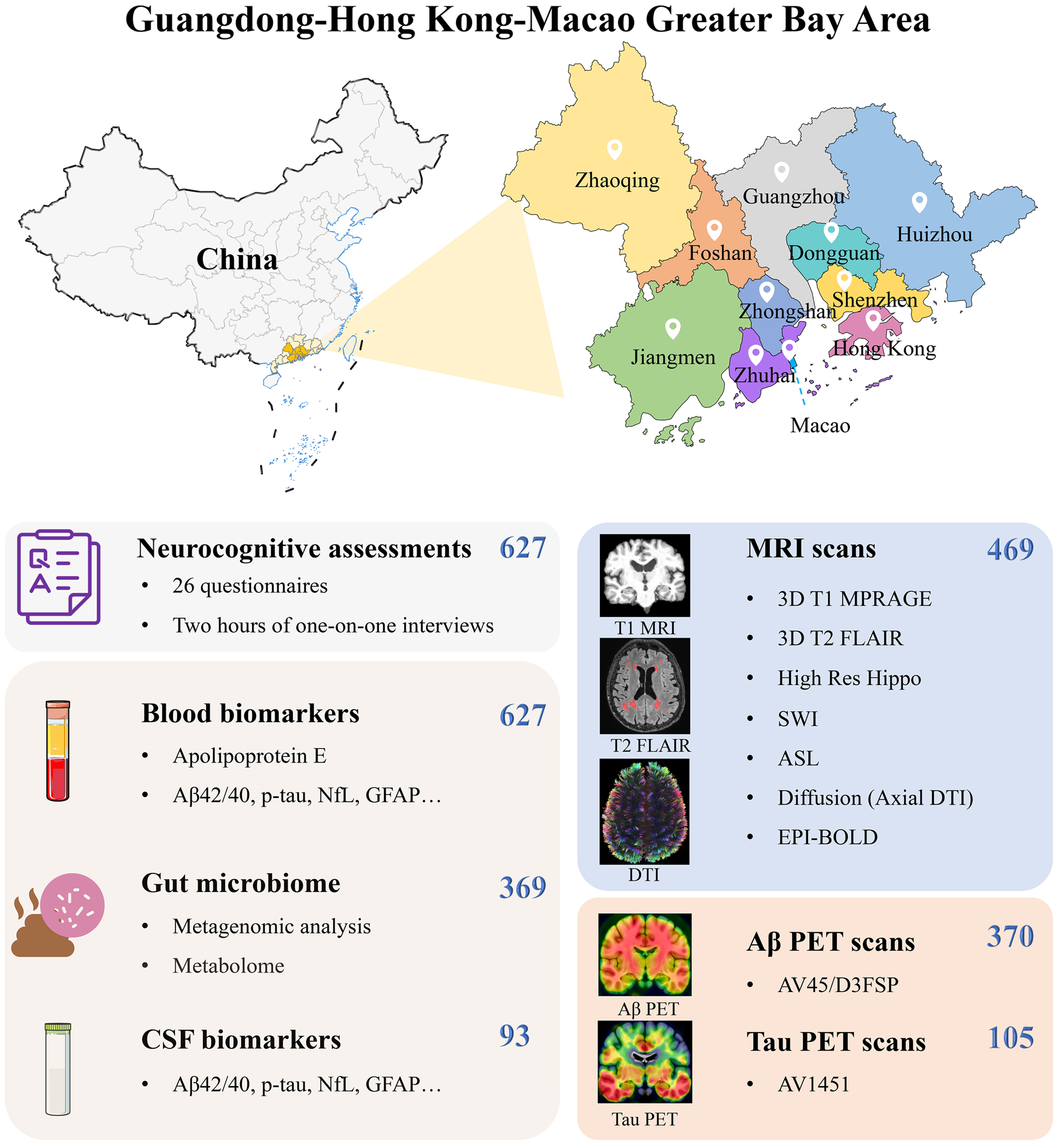
Several limitations should be addressed for our current work. As we know, temporal meta-ROI has been regarded as an early composite region of tau aggregation formation in AD according to post-mortem [13] and neuroimaging [14, 15] studies, and this composite region, therefore, has been commonly used to detect early tau depositions [5, 24, 49, 50]. Nevertheless, several recent studies highlighted the heterogeneity of tau spreading patterns [22, 51,52,53]. Thus, it still requires further investigation to confirm whether the findings in the present study can extend to distinct trajectories of tau spreading in AD. Secondly, the interpretations of the findings related to sex, age, and KL-VShet should be cautious as we used only one observational cohort and may need to be confirmed in other independent cohorts. Additionally, such a limited sample size constrained our capability to investigate the effects of baseline Aβ plaques/tau tangles on longitudinal tau accumulation in A+/T− and A+/T+ profiles, which might also be modulated by sex, age, and genotyping factors. Thus, future studies with a larger sample size of longitudinal tau PET would be beneficial.
Overall, our findings suggest that initial levels of Aβ plaques and tau tangles might be differently linked to the further spreading of tau aggregates in the early Braak stage of AD. Specifically, both initial Aβ plaques and tau tangles may be related to faster tau accumulation in early tau-deposited regions (entorhinal, amygdala, and parahippocampal gyrus) for A+/T− individuals, whereas further longitudinal tau propagation in late tau-deposited regions (fusiform, inferior temporal and middle temporal lobes) is mainly associated with the existing tau tangles for A+/T+ individuals. Furthermore, age, sex, and KL-VShet all play a role in modulating the relationships between baseline levels of Aβ plaques/tau tangles and subsequent tau accumulation. These findings provide novel insights into understanding the spatial and temporal patterns of tau accumulation in AD and highlight the effects of biological stages, Aβ plaques, tau tangles, age, sex, and KL-VShet genotyping on longitudinal tau accumulation.

留言 (0)