
記住我


According to the search strategy we applied, the literature search provided 594 records, including 151 articles from WOS, 100 from PubMed, and 343 from Scopus. Removing duplicate reports resulted in 337 articles available for the initial assessment. We assessed the titles and abstracts and removed irrelevant articles, limiting our data to 73 articles for full-text review. Out of 73, 32 were excluded based on inaccessibility and exclusion criteria. Eventually, data were collected from 41 articles presenting the role of Nrf2 in melanoma initiation, progression, and metastasis (Fig. 1).
Fig. 1
This flow diagram presents the process of study selection included in the qualitative systematic review
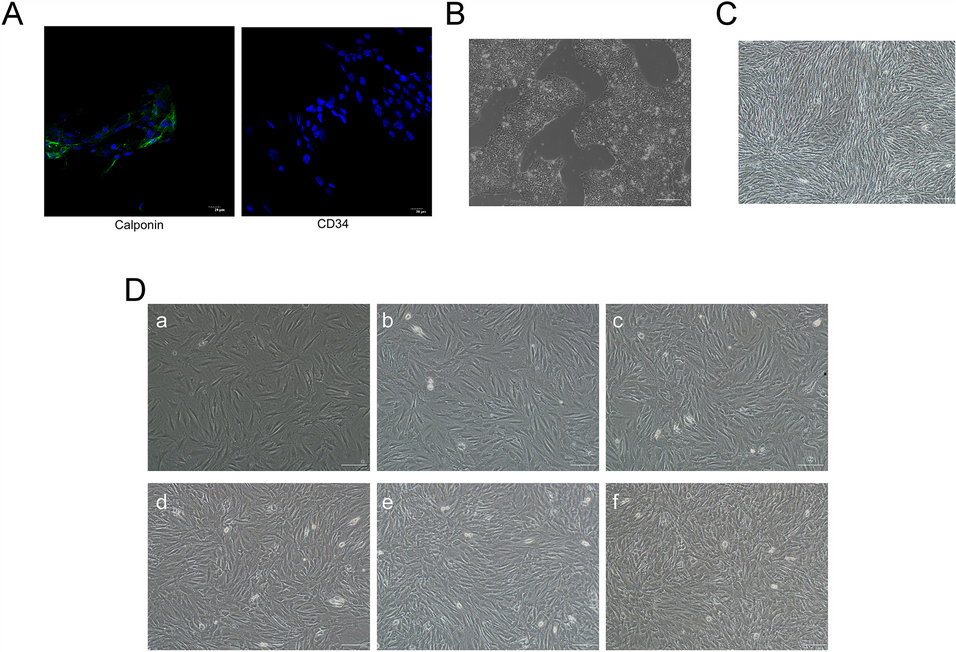
TabulationAll data extracted from the final articles are summarized in Table 1. All of the articles chosen for this study were published after 2014. Studies are mainly performed on human cancer cell lines, whereas others have focused on murine melanoma cell lines, mice, or human tissue samples.
Expression and activation of Nrf2 in melanomaDue to UV radiation and melanin production, melanocytes create more ROS than surrounding keratinocytes in the skin. Evidence shows that oxidative stress, increased ROS level, and redox imbalance are critical parameters in melanogenesis. Indeed, the entire process of melanoma formation relies on oxidative stress and antioxidant response. Because of their more significant metabolic needs, melanoma cells exhibit even higher ROS and antioxidant responses than normal melanocytes [21, 56]. Melanocytes use adaptive strategies to withstand oxidative stress by expressing antioxidant scavenging activity. As a result, it has been proposed that targeting melanoma cells’ addiction to greater redox capability could be a suitable and effective treatment method. The master stress response transcription factor Nrf2, which regulates the expression of antioxidant enzymes such as HO-1, SOD, CAT, and GPX, is primarily responsible for redox homeostasis in melanoma [57]. Investigations on the expression of Nrf2 and its target genes have produced conflicting results. Even some studies reported no significant differences in Nrf2 expression between normal and melanoma cells [16].
Taken together, it seems that lower Nrf2 expression favor melanogenesis, and is appeared in early stages of melanoma. Nrf2 mRNA and protein levels are diminished during the early phases of melanoma formation. Even though Nrf2 is expressed in normal skin cells, it is considerably reduced in skin malignancies such as malignant melanoma, indicating that Nrf2 expression is linked to the incidence of skin cancers. However, higher Nrf2 expression in cancerous cells is linked to more aggressive and resistant melanomas, and poor prognosis [15, 39]. It is hypothesized that Nrf2 contributes in melanoma phenotypic switch between a proliferative and an invasive phenotype. The expression of genes linked to the melanoma progression change subsequent of Nrf2 expression. For example, Nrf2 inhibits the MITF (microphthalmia-associated transcription factor), the primary transcriptional regulator of the phenotypic transition in melanoma, and its expression changes through the melanoma progression. MITF down regulation causes invasive dedifferentiated melanoma, while overexpression causes a proliferative phenotype. It is plausible that Nrf2 expression is required during periods of fast development but is repressed once an invasive phenotype has emerged [11].
When melanoma advances, Nrf2 expression rises. Melanoma redox capacity is assumed to represent a spectrum ranging from low in normal cells to moderate in drug-sensitive melanomas and to high in drug-resistant melanomas [57]. Consequently, the expression of Nrf2 changes in benign lesions and progresses to primary and metastatic melanomas [39]. Although Nrf2 expression is higher in metastatic lesions than in primary sites, cytoplasmic Keap1 expression is lower, indicating that Nrf2 accumulates in the nucleus, leading to up-regulation of Nrf2 target genes [28]. For example, HO-1 expression has been found to be increased in B16-F10 murine melanoma cells as well as in-vivo melanoma tumor models, which has been linked to the higher proliferation rate [57].
BRAF (B-rapidly accelerated fibrosarcoma) and NRAS (N-Rat sarcoma virus) mutations, which are found in 50 and 20% of cases, respectively, are commonly responsible for dysregulation of the MAPK (mitogen-activated protein kinase)-proliferation signaling pathway in melanomas [58]. A direct link between Nrf2 activation and MAPK signaling has recently been discovered. The activation of the MAPK pathway causes Nrf2 to be expressed. BRAF/NRAS mutant melanoma cells activate the Nrf2 signaling pathway to protect cells against oxidative stress [20, 49]. BRAF inhibitors (BRAFi), like vemurafenib, suppress Nrf2 expression in BRAF mutant melanoma cells, although the effect is not significant, suggesting that other oncogenes also trigger the Nrf2 signaling pathway [20]. This is worth noting that BRAFi-resistant cells have higher Nrf2 expression than non-resistant cells [57]. However, not all Nrf2-dependent genes react the same way to BRAFV600E expression [43].
UV irradiation is also a determining factor in the expression of Nrf2 and subsequent antioxidant target genes. UV exposure enhances the Nrf2 expression and nuclear accumulation in a dose-dependent manner [17, 30, 59]. However, the effect of UV radiation on Nrf2 expression is still a matter of contention; some studies have reported no effect or even a negative effect [29]. GSH (glutathione) depletion and ROS accumulation caused by UVA (ultraviolet A) irradiation may trigger melanogenesis in people with defective Nrf2. Although Nrf2 silencing did not affect melanogenesis in the absence of UVA exposure, Nrf2-knockout cells exhibit higher tyrosinase activity and melanin content [21].
Nrf2 in melanoma progressionNrf2, a crucial activator of antioxidant and phase two defenses in cells, has a dual role in melanoma. Although regular Nrf2 expression has a cytoprotective role against carcinogenesis, Nrf2 overexpression boosts antioxidant responses and changes the cellular redox state. In this aspect, antioxidants have been demonstrated to aid melanoma cell survival, proliferation, and tumor growth [44]. Nrf2 activity in transformed cells lowers ROS and inhibits tumor growth in vivo by mechanisms such as apoptosis sensitization and a reduced angiogenic/hypoxic response via HIF-1(hypoxia-inducible factor-1) instability and VEGFA (vascular endothelial growth factor A) suppression [16]. Nrf2 activators like cynaropicrin can reduce the clonogenic ability of metastatic melanomas in a dose- and time-dependent manner while not affecting normal melanocytes. Anti-apoptotic proteins such as XIAP (X-linked inhibitor of apoptosis protein) and Bcl-2 (B cell lymphoma-2) are also reduced in response to cynaropicrin treatment [45]. Another study discovered that delphinidin, an Nrf2 epigenetic enhancer, suppresses JB6P+ cell transformation, implying that delphinidin could be employed as a chemopreventive medication [40]. However, Nrf2 activators or inhibitors should be utilized carefully and in line with the stage of the disease in order to be effective.
UV-induced aggregation of Nrf2 in B16-F10 cells was found to diminish cell viability and increase caspase-3 expression, a key indicator of apoptosis. Surprisingly, ionizing radiation in Nrf2 silenced cells synergistically affected melanoma cell viability, elevating apoptosis [30]. This dual behavior may be due to the complex web of interactions around Nrf2. As Arslanbaeva et al. indicated, in vitro and in vivo, the combination of brusatol, a potent Nrf2 inhibitor from the plant Brucea javanica, and UVA irradiation could reduce the melanoma cell proliferation and induce apoptosis in melanoma cells [57].
In melanoma, the activity of receptor tyrosine kinases (RTKs) is normally restricted. Some RTK family members like EGFR may be expressed in MITF-low melanoma cells, and under this circumstance, the EGFR expression is linked to a pro-invasive and pro-metastatic melanoma phenotype with resistance to BRAF/MEK inhibitors. Stress-induced Nrf2 mediates the expression and activity of EGFR by increasing the EGFR levels and its ligands like EGF and TGFα. A canonical ARE is found in the EGF promoter, and Nrf2 binds to it directly, leading to a higher level of EGF for AKT activation. Furthermore, Nrf2 blocks the MITF activity, the melanocytic lineage factor responsible for suppression of EGFR and TGFA. Hence, Nrf2 indirectly derepresses the EGFR, contributing in the maintenance of EGFR-expressing melanomas [46]. Reversely, EGFR stimulates Nrf2, providing a positive feedback loop. In vivo Nrf2 silencing has been shown the low EGFR expression and proliferation rate, preventing melanoma growth, and increasing tumor-free survival. However, Nrf2-induced suppression of MITF activity also contributes to development of dedifferentiated, invasive phenotype in another ways. The antigenicity of melanoma cells is significantly influenced by the proteins expressed by MITF target genes. Nrf2-mediated down regulation of pigmentation markers (melanoma antigens) due to MITF inhibition promote a dedifferentiated melanoma cell that can escape from immune recognition [44]. Another study has demonstrated that inhibiting Nrf2 by pharmacologic activation of the Nrf2’s upstream gene reduces proliferation, accelerates cell death, and slows melanoma progression [42]. Inhibiting either Nrf2 or its target gene, HMOX-1 (heme oxygenase 1 gene) prevents melanosphere formation, an indicator of cell tumorigenic potential, in melanoma cell lines. Moreover, HMOX-1 overexpression in A375 melanoma cells increases tumor volume, whereas silencing HMOX-1 reduces tumor growth in mouse models [43].
The cellular redox status has been shown to alter the innate immune responses. Albeit this has yet to be determined, Nrf2 may cooperatively modulate the innate immune response, repressing the pro-inflammatory mediators’ expression by mediating the transactivation of antioxidant and other cytoprotective genes [60]. Nrf2 depletion can increase the susceptibility to infection and inflammatory diseases due to impaired cellular stress response and exacerbated immune-mediated hypersensitivity and autoimmunity [61]. Nrf2 can also aid anti-tumor immunity by inducing immune-dependent cascades such as IL-17D-dependent NK (natural killer) cell recruitment early in carcinogenesis before the pro-tumor action of Nrf2 manifests. However, induction of IL-17D at advanced stages is not always associated with a good prognosis [23].
There is evidence that Nrf2 has a significantly greater tendency to suppress immunity, mostly in advanced stages. BRAF wild-type melanomas have been treated with monoclonal antibodies that target immune checkpoint proteins such as anti-PD-1 (programmed cell death protein-1) (like pembrolizumab or nivolumab). Nrf2 inhibits the recruitment of interferon-secreting immune cells, reducing PD-L1 (programmed death-ligand 1) expression in the process. Given that increased PD-L1 expression has been linked to enhanced anti-PD-1 therapeutic effectiveness, this might explain why Nrf2 overexpression is linked to immunotherapy resistance [11]. Nrf2 has also been found to have a direct impact on PD-L1 expression. Zhu et al. found that Nrf2 stimulates PD-L1 transcription, and targeted Nrf2 inhibition is an alternative way of suppressing PD-1/PD-L1 to trigger tumor infiltration by CD4+ and CD8+ T lymphocytes and consequentially restrict melanoma growth [34].
Cytosolic DNA induced from nuclear/mitochondrial DNA damage in cancer triggers the activation of cGAS-STING (cyclic GMP-AMP synthase-stimulator of interferon genes) pathway. The recruitment of cGAS-STING pathway plays an important role in tumor immunogenicity, and Its activation has been demonstrated to reduce mice’s resistance to PD-1 blockage [62, 63]. Evidence exists to show the suppressive role of Nfr2 on cGAS-STING pathway, providing an immune-cold microenvironment [44, 64]. Moreover, Nrf2 is demonstrated to be a potent inducer of COX2, the enzyme that facilitate the production of PGE2. Following this, the PGE2 contributes to the suppression of the innate immune response and the attenuation of T cell receptors, establishing an immune-evasive tumor microenvironment [64, 65]. Nrf2 inhibitors are thus expected to function in conjunction with checkpoint inhibitors in the treatment of melanoma. Such a combination is presently not possible due to the lack of potent Nrf2 inhibitors. Nevertheless, preclinical studies have shown that COX inhibitors and STING agonists work in conjunction with anti-PD-1 immunological therapy, indicating that targeting Nrf2 downstream indicators may be beneficial [42, 44].
Overall, it can be concluded that the onco-promoter/onco-suppressor function of Nrf2 varies by cell stage and is context-dependent. Furthermore, whereas transient Nrf2 activity protects against cancer, constitutive Nrf2 activation caused by Keap1 or Nrf2 genetic variations may promote cancer progression and tumor growth (Fig. 2).
Fig. 2
Nrf2 plays a dual function in melanoma, depending on the cell stage. The standard expression of Nrf2 has a protective role against oxidative stress and cell damage, maintaining normal melanocytes. Nrf2 depletion and ROS accumulation make cells susceptible to transformation and cancer initiation. Evidence exists to show that reduced Nrf2 expression is associated with the incidence of melanoma. On the contrary, aberrant overexpression or constitutive activation of Nrf2 leads to invasive, metastatic, and radio/chemoresistant melanomas with poor prognosis. Indeed, after cell transformation has occurred, Nrf2 can become a pathway for cancer progression
Nrf2, and melanoma metastasisThere is evidence that Nrf2 activation enhances cancer cell metastasis in melanoma, suggesting that Nrf2 has an onco-promoter role [34, 47, 59]. In fact, as long as the cell is intact, Nrf2 and its lower genes try to save the cell from becoming cancerous by reducing oxidative stress. However, if the situation progresses in such a way that cell transformation occurs, the cancer cell uses Nrf2 in favor of itself, and higher expression of Nrf2 makes melanoma cells resistant to high levels of ROS and oxidative stress produced from cell metabolism. In this regard, Nrf2 is associated with invasiveness and pro-metastatic features. Studies have shown that the ROS or ROS-enhancing chemicals at this phase might be effective in cancer treatment, whereas antioxidants provide a more desirable environment for melanoma progression and metastasis [32, 45, 57, 66].
HMOX1 is one of the driving forces in melanoma invasion and migration. Silencing Nrf2, which causes HO-1 to be down-regulated, prevents melanoma cells from invasion and migration [43]. UV irradiation also has a significant role in regulating the apoptosis, migration, and invasion of melanoma cells in the Nrf2-dependent manner. Gao et al. found that downregulation of Nrf2 reduces the ability of radiation-exposed B16-F10 cells to invade and migrate. Surprisingly, however, overexpression of Nrf2 exhibited similar effects on cell survival, invasion, and migration, demonstrating Nrf2’s dual function in melanoma [30]. It has been discovered that inhibiting the function of Nrf2 by pharmacologically activating the STING pathway accelerates cell death and decreases melanoma cell migration, and has a synergistic effect with BRARi in cell death [42].
It has been shown that FAM129B, an Nrf2 inducer, also makes melanoma cells more susceptible to metastasis. Intriguingly, knocking down FAM129B or reducing its phosphorylation reduces melanoma cell invasion and migration in an Nrf2-dependent way [49]. In metastatic melanomas, mGPDH (mitochondrial glycerol-3-phosphate dehydrogenase) activity is reduced. Li et al. showed that in A375 melanoma cells, knocking down mGPDH activates the Nrf2 pathway, causing melanoma cell metastasis, whereas co-inhibiting Nrf2 prevents melanoma cell metastasis in vivo and in vitro. These findings show that via inhibiting the Nrf2 signaling pathway, mGDPH suppresses melanoma cell motility and invasion [47]. P62, an autophagy-degrading factor, is overexpressed in malignant and metastatic melanomas regardless of BRAF/NRAS mutant status. Overexpression of P62 has been shown to promote tumor growth and metastasis in various malignancies, including melanoma. P62 accumulates in melanocytes and melanoma cells due to different stimuli, including UVA [33, 49]. P62 interacts with Keap1, preventing it from inhibiting Nrf2 and boosting Nrf2. Surprisingly, p62 is a direct target of Nrf2, and the two are linked in a positive feedback loop. UVA-induced p62 upregulation is inhibited when Nrf2 is silenced. P62 knockdown, on the other hand, hinders Nrf2 overexpression. As a result, p62’s oncogenic activity may be influenced by Nrf2 antioxidant responses [49, 67].
Nrf2 has also been linked to establishing the metastatic niche and stromal progression in various studies. In the pre-metastatic liver of melanoma-bearing mice, a reduction in Nrf2 expression was discovered, leading to oxidative stress and acidosis. This state is consistent with the tumor microenvironment, enhances melanoma cell survival, and facilitates metastasis [36]. These findings imply that redox imbalance changes the environment in favor of metastasis and that the Nrf2 antioxidant response is critical in this process.
There is also evidence of the preventive role of Nrf2 in metastasis. The phytochemical cynaropicrin enhances the Nrf2 activity in melanoma cells. Cicco et al. proved that cynaropicrin reduces the capacity of human melanoma cells to migrate, invade, and proliferate in vitro. However, this phytochemical can also suppress MAPK/ERK (extracellular signal-regulated kinase) and NFκB (nuclear factor kappa-light-chain-enhancer of activated B cells) pathways in human melanoma cells. Thus we cannot consider the anticancer effects of cynaropicrin to be attributed only to Nrf2 stimulation [45]. Zhu et al. reported that B16-F10 melanoma cells lacking Nrf2 are more prone to tumor development and lung metastasis [24]. Weitzenböck et al. found that Nrf2 suppression leads to the expression of CD44 and the development of epithelial-mesenchymal transition phenotype. Of note, treatment of Nrf2 depleted cells with BRAF or MYC inhibitor results in a higher survival rate of melanoma cells [50]. This shows that antioxidant intervention could be a game-changing strategy for improving melanoma patient survival.
Taken together, it can be concluded that despite the contradictory findings regarding the role of Nrf2 in metastasis, most studies indicate the facilitative role of Nrf2 in this subject. Further investigation is necessary to find a clear conclusion as to whether or not nrf2 activity is metastatic.
Clinical role of Nrf2 in melanomaMelanoma is a poorly responsive malignancy with a 5-year survival of about 15% based on conventional chemotherapy [68]. In recent years, novel targeted treatments and immunotherapy have drastically altered the general approach to melanoma treatment. BRAF and NRAS mutations in the MAPK signaling pathway are found in about half of all melanoma cases [58], resulting in constitutive activation of the MAPK pathway.
In patients with advanced mutant BRAF melanoma diseases, BRAFi like as vemurafenib or dabrafenib have been used successfully in conjunction with MEK (mitogen-activated protein kinase) inhibitors such as trametinib. Moreover, for BRAF wild-type melanomas, current recommendations advocate using monoclonal antibodies (i.e., ipilimumab) that target immunological checkpoint proteins such as anti-PD-1 or CTLA-4 (cytotoxic T-lymphocyte antigen 4) in combination with anti-PD-1 therapy [69].
Despite the fact that new agents for melanoma have improved survival compared with conventional chemotherapy, the response rate of metastatic melanoma is still low and has little influence on prognosis. Malignant melanoma cells’ extensive resistance typically limits modern treatments’ efficacy. Adaptive processes of melanoma cells allow them to offset the effects of medicines, resulting in a multidrug-resistant phenotype [70]. Melanoma cells rewire their proliferation and survival pathways, putting their inherent resistance to apoptosis and ferroptosis to work, making them ‘bulletproof’ against a wide range of chemotherapeutic medicines [71]. Identifying signaling pathways involved in the regulation and execution of apoptosis and altering them to promote melanoma cell sensitivity has offered a unique method for battling melanoma chemoresistance in recent years. In this context, a mountain of evidence suggests that metabolic modification plays a significant role in melanoma cells’ chemo- and radio-resistance. During melanoma progression, metabolic reprogramming, including redox status, results in constitutive activation of Nrf2 and its downstream enzymes. As a result, intracellular ROS levels drop considerably, conferring chemoresistance [18, 72]. Therefore, Nrf2 is critical for controlling the redox system and fostering melanoma resistance cells through mediating downstream antioxidant enzymes, detoxifying and drug-metabolizing enzymes, drug transporters, and multidrug resistance proteins [73]. Although melanoma cells often create reactive oxygen species, the co-evolution of adaptive systems, specifically Nrf2 as a master regulator, makes them more sensitive to emerging chemoresistant cells’ ROS tolerance. In vitro studies have shown that suppressing Nrf2 can make melanoma cells more sensitive to anticancer medicines. As expected, Nrf2 devoid cells have increased ROS levels, making them more susceptible to anticancer drugs such as dacarbazine (DTIC) or cisplatin [18, 55]. Thus targeting Nrf2 may prevent melanoma from further progression and chemoresistance. Nevertheless, whether targeting Nrf2 alone is more beneficial or detrimental is still debated. If Nrf2 is targeted as a single therapy, melanoma phenotypic change will likely occur. Thus A combinatorial strategy would very certainly be required [11].
Drugs that reduce the corticosterone production or its effect on the receptor indirectly downregulate Nrf2, preventing tumor growth. In contrast, the induction of Nrf2 overexpression reversed this anti-tumor effect [26, 41]. According to the Rocha study, TMZ (temozolomide) promotes Nrf2 activation, which leads to the expression of the GCLM (glutamate-cysteine ligase modifier) and GST (glutathione S-transferase) genes. Melanoma cells eventually develop TMZ resistance due to high levels of GSH. GSH depletion, on the other hand, boosted TMZ sensitivity in cells [25]. As a result, using TMZ in conjunction with Nrf2/GSH inhibitors enhances therapy outcomes.
According to multiple studies, RAF/RAS isoforms activate the Nrf2 pathway, and BRAF/NRAS mutant melanomas express more Nrf2 [57, 74]. Melanoma cells also accumulate more Nrf2 in their nuclei, much higher in MAPKi-resistant cells. Moreover, MAPKi-resistant melanomas exhibit higher oxidative phosphorylation and oxygen consumption in their mitochondria. As a result of the robust activation of the Nrf2 signaling pathway, these cells can modify mitochondrial metabolism, allowing MAPKi-resistant cells to survive. In BRAFi-resistant cells, Nrf2 silencing improves the efficacy of treatments like vemurafenib or PLX4032 [31,
留言 (0)