
記住我
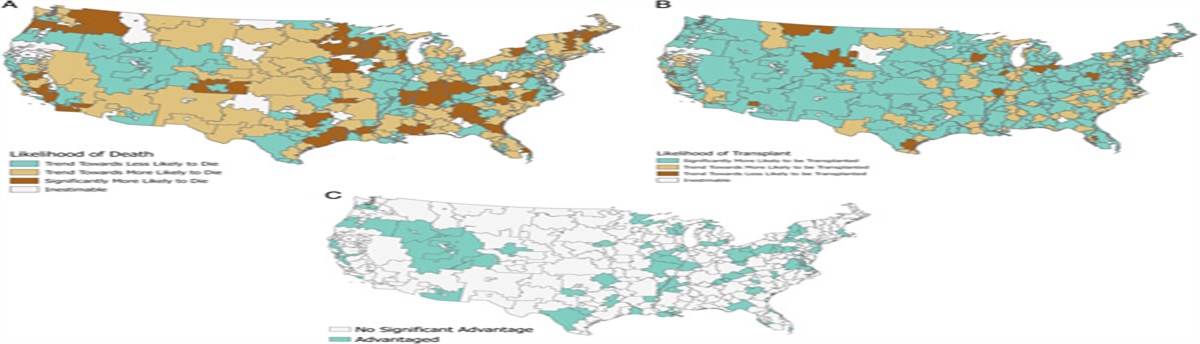


Trauma is one of the leading causes of death and disability in those aged under 44 and exceeds the number of deaths caused by HIV, tuberculosis and malaria combined.1 Globally there are approximately 6 million trauma-related deaths each year. Trauma-associated hemorrhage accounts for almost 40% of all trauma mortalities2 and is a key driver of multiple organ dysfunction (MODS).3 The mechanisms contributing to MODS include a) an excessive systemic inflammatory response secondary to the release of damage-associated molecular patterns from extensive tissue damage and b) ische-mia-reperfusion (I/R) injury.4 There are no specific pharmacological interventions that prevent the onset of MODS associated with HS.
Bruton's tyrosine kinase (BTK) is a cytoplasmic, non-receptor protein tyrosine kinase first discovered in X-linked agammaglobu-linemia.5 All cells of hematopoietic origin except plasma cells, natural killer cells and T-lymphocytes express BTK.6 Whilst BTK was initially known for its critical role in B-lymphocyte development and, thus, adaptive immunity, more recent studies point to a pivotal role for BTK in innate immunity.7
Trauma leads to a so-called ‘genomic storm’ that results in a change in >80% of cellular functions and pathways8 including an increased expression of B-lymphocyte receptor signaling. Inhibition of BTK activity reduces multiple organ injury/dysfunction, activation of NF-κB and NLRP3 inflammasome as well as the release of cytokines and chemokines caused by cecum-ligation and puncture (CLP).9 Mice with inactive BTK (X-Linked immunodeficient) are protected from sepsis-induced MODS.10 Prevention of NF-κB activation reduces the MODS and improves survival rate in rodent models of septic shock11,12 and HS.13
Driven by the COVID-19 pandemic, there has been a significant focus on repurposing strategies that dampen the cytokine storm and pulmonary injury linked to severe acute respiratory syndrome corona-virus 2 (SARS–CoV–2). Interestingly, there is a positive correlation between disease severity and BTK activity following SARS–CoV–2 infection.14–16 Thus, BTK inhibitors (BTKi) appear to reduce systemic inflammation and ongoing clinical trials will assess the potential impact of this repurposing strategy on outcome in COVID–19 patients (ClinicalTrials.gov Identifier: NCT04382586, NCT04665115, NCT04439006, NCT04375397, NCT04528667 and NCT04440007).
BTKi are commonly used in patients with B-lymphocyte malignancies (chronic lymphocytic leukemia, mantle cell lymphoma) and have received approval by the FDA for patients with marginal zone lymphoma, small lymphocytic lymphoma, Waldenstrom’s macroglobulinemia and chronic graft versus host disease.17 Given the evident protective effects of BTKi administration in sepsis and COVID-19, we wished to explore the potential of repurposing BTKi in trauma-hemorrhage. Currently, there is limited information about the role of BTK in trauma.18
Methods BTK Gene Expression in Human Whole BloodOriginal data was obtained under Gene Expression Omnibus (GEO) accession GSE36809, published by Xiao and colleagues.8 RNA was extracted from whole blood leukocytes of severe blunt trauma patients (n = 167) over the course of 28 days and healthy controls (n = 37) and hybridized onto an HU133 Plus 2.0 GeneChip (Affymetrix) according to the manufacturer’s recommendations. The dataset was reanalyzed for BTK gene expression.
Use of Experimental Animals - Ethical StatementFor the short-term follow-up acute HS model, all animal procedures were approved by the Animal Welfare Ethics Review Board of Queen Mary University of London and by the Home Office (License number PC5F29685). For the long-term follow-up acute HS model, all animal procedures were approved by the Universidade Federal de Santa Catarina Institutional Committee for Animal Use in Research (License number 7396250219) in accordance with the Brazilian Government Guidelines for Animal Use in Research.
Experimental DesignMale Wistar rats (for short-term follow-up acute model: Charles River Laboratories Ltd., UK; for long-term follow-up acute model: Universidade Federal de Santa Catarina, Brazil) weighing 250–350 g were kept under standard laboratory conditions and received a chow diet and water ad libitum. Acalabrutinib (3 mg/ kg; Insight Biotechnology, UK) and fenebrutinib (3 mg/kg; Insight Biotechnology, UK) were separately diluted in 5% DMSO + 95% Ringer's Lactate (vehicle) and rats were treated (i.v. in short-term follow-up and i.p. in long-term follow-up) upon resuscitation. Further information about acalabrutinib and fenebrutinib can be found in the supplemental, https://links.lww.com/SLA/D600.
Acute Hemorrhagic Shock Model (Short-Term Follow-up)The short-term follow-up acute HS model was performed as previously described in this journal.19–21 Briefly, fifty-four rats were anesthetized with sodium thiopentone (120 mg/kg i.p. initially and 10 mg/kg i.v. for maintenance as needed) and randomized into six groups: Sham + vehicle (n = 9); Sham + acalabrutinib (3 mg/kg; n = 8), Sham + fenebrutinib (3 mg/kg; n = 8), HS + vehicle (n = 9); HS + acalabrutinib (3 mg/kg; n = 10); HS + fenebrutinib (3 mg/kg; n = 10). Blood was withdrawn to achieve a fall in mean arterial pressure (MAP) to 35 ± 5 mmHg, which was maintained for 90 minutes. At 90 minutes after initiation of hemorrhage (or when 25% of the shed blood had to be reinjected to sustain MAP), resuscitation was performed. At 4 hours post-resuscitation, blood was collected for the measurement of biomarkers of organ injury/dysfunction (MRC Harwell Institute, Oxfordshire, UK). Sham-operated rats were used as control and underwent identical surgical procedures, but without hemorrhage or resuscitation. Detailed description of the short-term follow-up model can be found in the supplemental (Supplemental Digital Content Figure 1A, https://links.lww.com/SLA/D600).
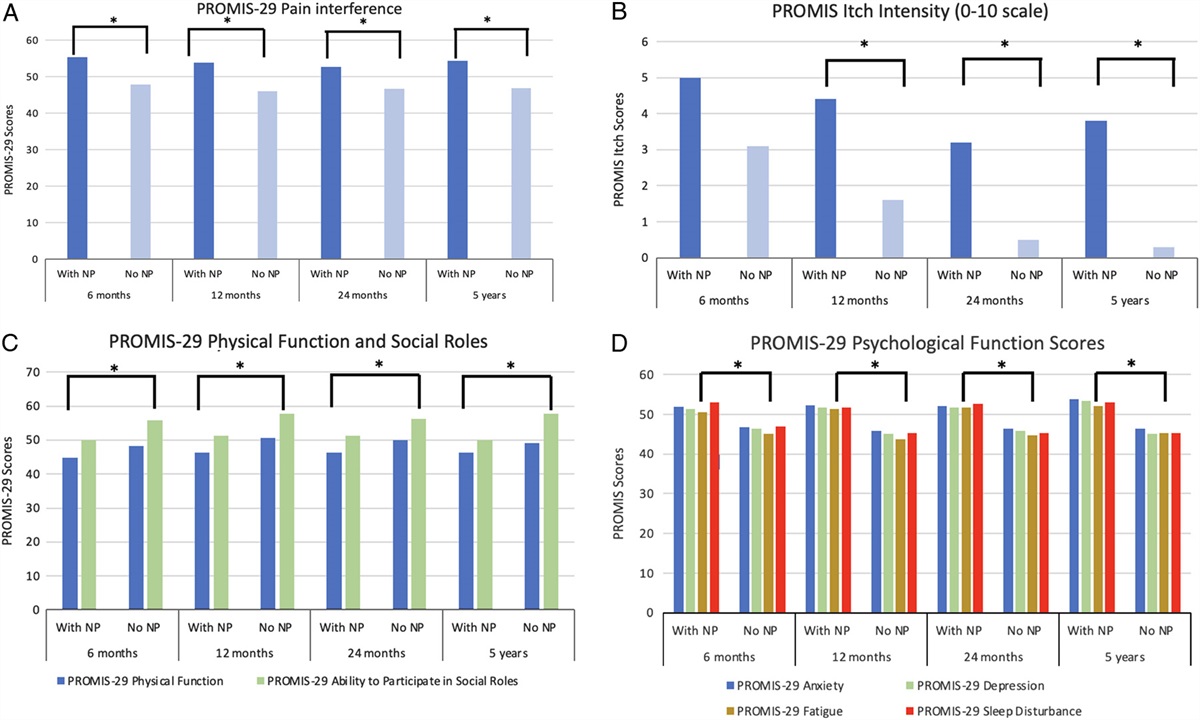
 Figure 1:
Figure 1: Treatment with BTKi improves HS-induced circulatory failure in a short-term follow-up acute HS model. (A) Mean arterial pressure (MAP) was measured from the completion of surgery to the termination of the experiment for vehicle and BTKi treated (acalabrutinib, ACA;fenebrutinib, FEN) rats. (B) MAP values at the end of the resuscitation period (330 min). Data are expressed as mean ± SEM of 8–10 animals per group. Statistical analysis was performed using two-way ANOVA followed by a Bonferroni’s post-hoc test. *P < 0.05 Sham + vehicle vs. HS + vehicle; #P < 0.05 HS + vehicle vs. HS + BTKi (ACA or FEN).
Acute Hemorrhagic Shock Model (Long-Term Follow-Up)Thirty-eight rats were administered analgesia with tramadol (10 mg/kg i.p.) 15 minutes prior to anesthesia induction with ketamine and xylazine (100 mg/kg and 10 mg/kg i.m. respectively) and randomized into four groups: Sham + vehicle (n = 9); Sham + acalabrutinib (3 mg/kg; n = 10); HS + vehicle (n = 10); HS + acalabrutinib (3 mg/kg; n = 9). Blood was withdrawn to achieve a fall in MAP to 40 ± 2 mm Hg, which was maintained for 90 minutes. At 90 minutes after initiation of hemorrhage (or when 25% of the shed blood had to be reinjected to sustain MAP), resuscitation was performed. At 24 hours post-resuscitation, blood was collected for the measurement of organ injury/dysfunction parameters. Sham-operated rats were used as control and underwent identical surgical procedures, but without hemorrhage or resuscitation. Detailed description of the long-term follow-up model can be found in the supplemental (Supplemental Digital Content Figure 1B, https://links.lww.com/SLA/D600).
Western Blot AnalysisSemi-quantitative immunoblot analysis was carried out in kidney samples as previously described.22 Detailed description of the method can be found in the Supplemental Digital Content, https://links.lww.com/SLA/D600.
Quantification of Myeloperoxidase ActivityDetermination of myeloperoxidase activity in lung and liver tissue samples was performed. Detailed description of the method can be found in the Supplemental Digital Content, http://link-s.lww.com/SLA/D600.
Statistical AnalysisAll figures are expressed as mean ± SEM of n observations, where n represents the number of animals/experiments/subjects studied. Measurements obtained from the patient groups and vehicle and BTKi treated animal groups were analyzed by one-way ANOVA followed by a Bonferroni’s post-hoc test on GraphPad Prism 8.0 (GraphPad Software, Inc., La Jolla, CA). The distribution of the data was verified by Shapiro-Wilk normality test, and the homogeneity of variances by Bartlett test. When necessary, values were transformed into logarithmic values to achieve normality and homogeneity of variances. P < 0.05 was considered statistically significant.
Results BTK Gene Expression is Elevated in Trauma PatientsXiao and colleagues8 compared genome-wide expression in leukocytes from trauma patients against matched healthy controls. We reanalyzed this dataset for BTK expression. When compared to healthy controls, BTK expression was significantly elevated at all time points except 12 hours (P < 0.05; Supplemental Digital Content Figure 2, https://links.lww.com/SLA/D600). An initial peak was observed at Day 1 followed by a gradual decrease, however, BTK expression remained elevated at Day 28.
 Figure 2:
Figure 2: Treatment with BTKi attenuates HS-induced organ damage in a short-term follow-up acute HS model. Rats were subjected to hemorrhagic shock (HS) and 4 h after resuscitation, levels of serum (A) urea, (B) creatinine, (C) alanine aminotransferase (ALT), (D) aspartate aminotransferase (AST), (E) creatine kinase (CK) and (F) lactate dehydrogenase (LDH) were determined in vehicle and BTKi treated (acalabrutinib, ACA;fenebrutinib, FEN) rats. Sham-operated rats were used as control. Data are expressed as mean ± SEM of 8–10 animals per group. Statistical analysis was performed using one-way ANOVA followed by a Bonferroni‘s post-hoc test. *P < 0.05 denoted statistical significance.
BTK Gene Expression does not Differ Between Uncomplicated and Complicated Recovery Patient GroupsXiao and colleagues8 also stratified their trauma patient cohort into uncomplicated (recovery in <5 days) and complicated (recovery after 14 days, no recovery by Day 28 or death) to further identify genotypic differences. We reanalyzed this dataset for BTK expression using this stratification. When comparing uncomplicated and complicated patients, there were no significant differences at any of the timepoints measured (P > 0.05; Supplemental Digital Content Figure 3, https://links.lww.com/SLA/D600).
 Figure 3:
Figure 3: Treatment with BTKi attenuates BTK, NF-κB and NLRP3 activation in a short-term follow-up acute HS model. (A) The phosphorylation of BTK at Tyr223, (B) the phosphorylation of IKKa/β at Ser176/180, (C) the phosphorylation of IkBα at Ser32/36, (D) the nuclear translocation of p65, (E) the activation of NLRP3 and (F) the cleaved (activated) form of caspase 1 of vehicle and BTKi treated (acalabru-tinib, ACA; fenebrutinib, FEN) rats were determined by western blotting in the kidney. Protein expression was measured as relative optical density (O.D.) and normalized to the sham band. Data are expressed as mean ± SEM of five animals per group. Statistical analysis was performed using one-way ANOVA followed by a Bon-ferroni’s post-hoc test. *P<0.05 denoted statistical significance.
Treatment with BTKi Improves HS-Induced Circulatory Failure in a Short-Term Follow-Up Acute HS ModelTo investigate the effects of the BTK inhibitors (BTKi) acalabrutinib and fenebrutinib on circulatory failure, MAP was measured from the completion of surgery to the termination of the experiment. Baseline MAP values were similar amongst all six groups. When compared to sham-operated rats, HS-rats treated with vehicle exhibited a more pronounced decrease in MAP over time post-resuscitation (Fig. 1A). The MAP of HS-rats treated with either BTKi was significantly higher than that of vehicle treated HS-rats at the end of the resuscitation period (P < 0.05; Fig. 1B). No significant differences were observed between HS-rats treated with either BTKi (P > 0.05; Fig. 1A). Administration of either BTKi to sham-operated rats had no significant effect on MAP (P > 0.05; Fig. 1A). Whilst there were no statistically significant differences in MAP between vehicle treated and BTKi treated sham-operated rats, the BTKi treated sham rats presented with a slightly lower average pressure throughout the experiment. We do not believe this is due to BTKi administration (further information in the Supplement Digital Content, https://links.lww.com/SLA/D600).
Treatment with BTKi Attenuates HS-Induced Organ Damage in a Short-Term Follow-Up Acute HS ModelHere we explored whether pharmacological intervention with BTKi attenuates the MODS associated with HS in rats. When compared to sham-operated rats, rats subjected to HS and treated with vehicle displayed increases in serum urea (P < 0.05; Fig. 2A) and creatinine (P < 0.05; Fig. 2B); indicating the development of renal dysfunction. When compared to sham-operated rats, vehicle treated HS-rats exhibited significant increases in ALT (P < 0.05; Fig. 2C) and AST (P < 0.05; Fig. 2D) indicating the development of hepatic injury, while the increases in CK (P < 0.05; Fig. 2E) and amylase (P < 0.05; not shown) denote neuromuscular and pancreatic injury, respectively. The significant increase in LDH (P < 0.05; Fig. 2F) in vehicle treated HS-rats confirms tissue injury. Treatment of HS-rats with either BTKi significantly attenuated the renal dysfunction, hepatic injury, neuro-muscular injury and general tissue damage caused by HS as shown by the reduction in serum parameter values (all P < 0.05; Figs. 2A–F). Treatment with either BTKi had no significant effect on pancreatic injury (P > 0.05; not shown). No significant differences were observed between HS-rats treated with either BTKi (P > 0.05; Fig. 2). Administration of either BTKi to sham-operated rats had no significant effect on any of the parameters measured (P > 0.05; Fig. 2).
Treatment with BTKi Abolishes Renal BTK Activation in a Short-Term Follow-Up Acute HS ModelUsing western blot analysis, we examined whether HS leads to the activation of BTK in the kidney; given that treatment with either BTKi significantly attenuated HS-associated renal dysfunction. The activation of BTK and subsequent BTK-associated signaling pathways consists of the phosphorylation of BTK at Tyr223 as the initial stage of the BTK-signaling cascade. When compared to sham-operated rats, HS-rats treated with vehicle displayed significant increases in the phosphorylation of BTK at Tyr223, indicating that BTK is activated in injured kidneys (P < 0.05; Fig. 3A). Treatment with BTKi in HS-rats significantly abolished these increases (P < 0.05; Fig. 3A). No significant differences were observed between HS-rats treated with either BTKi (P > 0.05; Fig. 3A). These data demonstrate that both BTKi abolish the activation of BTK caused by HS.
Treatment with BTKi Abolishes Renal NF-κB Activation in a Short-Term Follow-Up Acute HS ModelThe effect of BTK inhibition on the activation of the signaling events leading to the activation of NF-κB, were investigated in the kidney. When compared to sham-operated rats, HS-rats treated with vehicle had significant increases in the phosphorylation of IKKα/β at Ser176/180 (P < 0.05; Fig. 3B), phosphorylation of IκBα at Ser32/36(P < 0.05; Fig. 3C) and the translocation of p65 to the nucleus (P < 0.05; Fig. 3D). Treatment of HS-rats with either BTKi significantly abolished these increases (P < 0.05; Figs. 3B-D). No significant differences were observed between HS-rats treated with either BTKi (P > 0.05; Figs. 3B-D). These data illustrate that both BTKi abolish the activation of NF-κB caused by HS.
Treatment with BTKi Abolishes Renal NLRP3 and Caspase 1 Activation in a Short-Term Follow-Up Acute HS ModelHaving discovered that BTKi significantly reduced the activation of NF-κB in the kidney of rats subjected to HS, we next analyzed the potential involvement of the NLRP3 inflammasome complex. When compared to sham-operated rats, HS-rats treated with vehicle exhibited a significantly increased expression of the NLRP3 inflammasome (P < 0.05; Fig. 3E) and of the cleaved (activated) form of caspase 1 (P < 0.05; Fig. 3F). Treatment of HS-rats with either BTKi significantly inhibited these increases (P < 0.05; Figs. 3E,F). No significant differences were observed in the degree of expression of the NLRP3 inflammasome and cleaved caspase 1 between HS-rats treated with either BTKi (P > 0.05; Figs. 3E,F). These data demonstrate that both BTKi abolish the activation of the NLRP3 inflammasome and caspase 1 and the subsequent formation of IL–1 β.
BTK Activation Correlates with Renal Dysfunction, NF-κB and NLRP3 Activation in a Short-Term Follow-Up Acute HS ModelCorrelation analysis was performed to determine whether the degree of activation of BTK correlates with changes in renal function, NF-κB and NLRP3 activation. Significant positive correlations were found between BTK activation and all parameters investigated (except urine creatinine; Supplemental Digital Content Figure 4, https://links.lww.com/SLA/D600).
 Figure 4:
Figure 4: Treatment with acalabrutinib improves HS-induced circulatory failure in a long-term follow-up acute HS model. (A) Mean arterial pressure (MAP) and (B) heart rate (HR) were measured 24 h post resuscitation for vehicle and acalabrutinib (ACA) treated rats. Sham-operated rats were used as control. Data are expressed as mean ± SEM of 9–10 animals per group. Statistical analysis was performed using one-way ANOVA followed by a Bonferroni's post-hoc test. *P < 0.05 denoted statistical significance.
Treatment with Acalabrutinib Improves HS-Induced Circulatory Failure in a Long-Term Follow-up Acute HS ModelWhen comparing acalabrutinib and fenebrutinib, both inhibitors were equally efficacious in the short-term follow-up model but acalabrutinib has the advantage of being FDA approved; hence was investigated in a long-term follow-up acute HS model. Having demonstrated treatment with acalabrutinib improved blood pressure in a short-term follow-up model, we wished to determine whether acalabrutinib would still be effective in a model in which the resuscitation period is prolonged to 24hours. When compared to sham-operated rats, HS-rats treated with vehicle had significantly lower MAP values recorded 24hours post-resuscitation (P < 0.05; Fig. 4A); highlighting that either cardiac dysfunction or excessive hypotension23 was still present. In contrast, MAP values of HS-rats treated with acalabrutinib upon resuscitation were significantly higher even at 24 hours post-resuscitation [when compared with those of vehicle treated rats (P < 0.05; Fig. 4A]. Administration of acalabrutinib to sham-operated rats had no significant effect on MAP (P > 0.05; Fig. 4A). There were no significant differences in HR between any of the groups (P > 0.05; Fig. 4B).
Treatment with Acalabrutinib Attenuates HS-Induced Organ Damage in a Long-term Follow-up Acute HS ModelHaving shown that treatment with acalabrutinib ameliorated the MODS associated with HS in a short-term follow-up model, we examined whether this effect was sustained when the resuscitation period was extended to 24hours. As with the short-term follow-up model, when compared to sham-operated rats, rats subjected to HS with long-term follow-up and treated with vehicle displayed significant increases in serum urea (P < 0.05; Fig. 5A) and creatinine (P < 0.05; Fig. 5B); indicating the development of renal dysfunction. When compared to sham-operated rats, vehicle treated HS-rats exhibited significant increases in ALT (P < 0.05; Fig. 5C) and AST (P < 0.05; Fig. 5D) indicating the development of hepatic injury, whilst the significant increase in LDH (P < 0.05; Fig. 5E) confirmed tissue injury. Treatment of HS-rats with acalabrutinib significantly attenuated the hepatic and general tissue injury caused by HS as shown by the decrease in serum parameter values (P < 0.05; Figs. 5C–E). Treatment with acalabrutinib had no significant effect on renal dysfunction (P > 0.05; Figs. 5A,B). Administration of acalabrutinib to sham-operated rats had no significant effect on any of the parameters measured (P > 0.05; Figs. 5A–E).
 Figure 5:
Figure 5: Treatment with acalabrutinib attenuates HS-induced organ damage and myeloperoxidase activity in a long-term follow-up acute HS model. Rats were subjected to hemorrhagic shock (HS) and 24 h after resuscitation, levels of serum (A) urea, (B) creatinine, (C) alanine aminotransferase (ALT), (D) aspartate aminotransferase (AST) and (E) lactate dehydrogenase (LDH) and mye-loperoxidase (MPO) activity in the (F) lung and (G) liver were determined for vehicle and acalabrutinib (ACA) treated rats. Sham-operated rats were used as control. Data are expressed as mean ± SEM of 9–10 animals per group. Statistical analysis was performed using oneway ANOVA followed by a Bonferroni’s post-hoc test. *P < 0.05 denoted statistical significance.
Treatment with Acalabrutinib Reduces Myeloperoxidase Activity in a Long-Term Follow-Up Acute HS ModelWe next determined myeloperoxidase (MPO) activity in the lung and liver as an indicator of neutrophil infiltration. When compared to sham-operated rats, HS-rats treated with vehicle showed a significant increase in MPO activity in the lung (P < 0.05; Fig. 5F) and liver (P < 0.05; Fig. 5G). Treatment with acalabrutinib in HS-rats significantly attenuated these rises in MPO activity (P < 0.05; Figs. 5F,G), suggesting reduced neutrophil recruitment and inflammation. Administration of acalabrutinib to sham-operated rats had no significant effect on pulmonary and hepatic MPO activity (P > 0.05; Figs. 5F,G).
DiscussionThis study reports that inhibition of BTK activity attenuates organ injury/dysfunction and circulatory failure in acute short-term follow-up (Figs. 1 and 2) and long-term follow-up (Figs. 4 and 5) rat models of HS. Having shown that BTK gene expression is significantly elevated in leukocytes of trauma patients (Supplemental Digital Content Figure 2, https://links.lww.com/SLA/D600), we used a reverse translational approach to investigate whether pharmacological intervention with acalabrutinib or fenebrutinib ameliorates the MODS associated with HS in a well-established rat model. Either irreversible (acalabrutinib) or reversible (fenebrutinib) inhibition of BTK activity significantly attenuated the fall in blood pressure (Fig. 1 short-term follow-up and 4 long-term follow-up) and, hence, the delayed vascular decompensation caused by HS.23 Moreover, BTKi significantly attenuated the renal dysfunction, hepatic injury and neuromuscular injury caused by HS (Fig. 2 short-term follow-up and 5 long-term follow-up). Similarly, BTKi also reduce disease severity in animal models of lupus nephritis,24,25 hepatic ischemia-reperfusion,26 acute lung injury27–29 and sepsis.9
What, then, are the mechanisms by which BTKi attenuate HS-associated organ injury/dysfunction? HS resulted in a significant increase in BTK activity in the kidney (Fig. 3), which correlated with the rise in serum creatinine and urea (Supplemental Digital Content Figure 4, https://links.lww.com/SLA/D600). Indeed, inhibition of BTK activity with acalabrutinib or fenebrutinib in the kidney of HS-rats attenuated the renal dysfunction, suggesting that activation of BTK plays a pivotal role in the pathophysiology. A positive correlation between BTK expression and creatinine also occurs in patients with IgA nephropathy30 and diabetic nephropathy.31
There is good evidence that BTK activation precedes the activation of NF-κB through TLR signaling32 and trauma results in elevated NF-κB translocation to the nucleus.19–22 Inhibition of BTK activity with acalabrutinib or fenebrutinib reduced NF-κB activation in the kidney of HS-rats (Fig. 3). We also found a significant positive correlation between the activation of BTK and the phosphorylation of IKKa/β at Ser176/180, phosphorylation of IkBα at Ser32/36 and translocation of p65 (Supplemental Digital Content Figure 4, https://links.lww.com/SLA/D600). This may suggest that inhibiting the activation of NF-κB contributes to the observed beneficial effects of BTKi in HS. Activation of NF-κB drives the formation of several pro- and anti-inflammatory mediators which include cytokines, chemokines and enzymes.33 As part of a positive feedback loop, these mediators can activate NF-κB and its upstream signaling components, further amplifying and perpetuating the inflammatory responses mediated by NF-κB which can lead to increased endothelial permeability, tissue hypoperfusion/hypoxia, tissue injury and ultimately MODS.11
There is also good evidence that BTK activation influences the assembly and activation of the NLRP3 inflammasome in rodents and humans.34–36 NLRP3 inflammasome activation drives the production of IL-1 β which plays a crucial role in the systemic inflammation and/or organ dysfunction in trauma.22 Inhibition of BTK activity with acalabrutinib or fenebrutinib reduced both the assembly and subsequent activation of the NLRP3 inflammasome in the kidney (Fig. 3). We also discovered a significant positive correlation between BTK activation and the activation of NLRP3 and caspase 1 (Supplemental Digital Content Figure 4, https://links.lww.com/SLA/D600). This may suggest that inhibiting the activation of the NLRP3 inflammasome contributes to the observed protective effects of BTKi in HS by lowering the pro-inflammatory effects of IL-1β and resulting tissue inflammation.37
The sterile inflammation caused by HS drives leukocyte recruitment to the tissues and is secondary to the activation of NF-κB and NLRP3 and their transcriptional regulation of pro-inflammatory cytokines.38–40 Moreover, the expression of adhesion molecules present on leukocytes and endothelial cells is regulated by NF-κB and permits leukocyte extravasation from the circulation to the site of injury.41 As neutrophils play a key role in HS-associated pulmonary and hepatic inflammation, we evaluated the degree of neutrophil recruitment (measured as MPO activity) in the lung and liver.42,43 HS resulted in a significant increase in pulmonary and hepatic MPO activity which was attenuated by BTKi in HS-rats (Fig. 5, see also Supplement for an extended discussion, https://links.lww.com/SLA/D600). A graphical abstract summarizing the protective effects of BTKi following HS-induced MODS is shown in Fig. 6. Please refer to the Supplement, https://links.lww.com/SLA/ D600 for a discussion of the limitations of this study.
 Figure 6:
Figure 6: Graphical abstract highlighting the role of Bruton’s tyrosine kinase (BTK) in the pathophysiology of traumatic injury. Pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs) are released following trauma which activate signaling pathways leading to NF-kB activation and pro-inflammatory cytokine production. BTK also activates the NLRP3 inflammasome which results in the cleavage of pro IL–1 β to IL–1 β and further pro-inflammatory cytokine production. Leukocyte recruitment is stimulated by chemotactic cytokines. Excessive systemic inflammation from the combination of cytokine release and innate immune cell recruitment contributes to the onset of multiple organ dysfunction syndrome (MODS). Treatment with BTK inhibitors such as acalabrutinib and fenebrutinib can attenuate trauma-induced inflammation and thus MODS to improve clinical outcomes.
Our results and conclusions are supported by findings in patient cohorts with either COVID-19 or sepsis where BTK has been proposed to play a role in disease pathogenesis; both diseases featuring the hallmarks of excessive systemic inflammation similar to those seen in trauma-associated MODS.15,44 The beneficial effects of acalabrutinib in COVID-19 patients, as measured by the determination of biomark-ers of inflammation, oxygenation and clinical status, imply that BTK activation plays a role in the pathology.15 Whilst Parnell and colleagues did not investigate pharmacological intervention with BTKi in patients with sepsis, reanalysis of microarray data (GEO Dataset Number GDS4971) revealed an increased expression (in whole blood) of BTK in septic non-survivors compared to septic survivors; highlighting the potential for BTK to be a predictor of mortality.10
ConclusionsIn conclusion, we report here for the first time that treatment with either the irreversible BTKi acalabrutinib or the reversible BTKi fenebrutinib reduces the organ injury/dysfunction and circulatory failure caused by severe hemorrhage in the rat; highlighting a role of BTK in disease pathogenesis. Moreover, experimental trauma-hemorrhage results in a significant upregulation of BTK in the kidney. Administration of either BTKi subsequently attenuates the degree of BTK activation as well as the activation of NF-κB and the NLRP3 inflammasome (measured in the kidney), both of which are key drivers of local and systemic inflammation. Notably, no significant differences were found between the two structurally and mechanistically different inhibitors, suggesting that the observed beneficial effects in experimental trauma-hemorrhage are most likely due to a drug class related effect. Thus, we propose that BTKi may be repurposed for the use in trauma patients to lower the organ injury and inflammation caused by severe hemorrhage and resuscitation.
References 1. World Health Organization. Injuries and Violence: The Facts 2014. 2014. 120. www.who.int/healthinfo/global_burden_disease/projections/en/. 2. Curry N, Hopewell S, Dorée C, et al. The acute management of trauma hemorrhage: a systematic review of randomized controlled trials. Crit Care. 2011;15:1–10. 3. Dewar D, Moore FA, Moore EE, et al. Postinjury multiple organ failure. Injury. 2009;40:912–918. 4. Lord JM, Midwinter MJ, Chen YF, et al. The systemic immune response to trauma: an overview of pathophysiology and treatment. Lancet. 2014;384:1455–1465. 5. Vetrie D, Vořechovský I, Sideras P, et al. The gene involved in X-linked agammaglobulinaemia is a member of the src family of protein-tyrosine kinases. Nature. 1993;361:226–233. 6. Mohamed AJ, Yu L, Bäckesjö CM, et al. Bruton's tyrosine kinase (Btk): function, regulation, and transformation with special emphasis on the PH domain. Immunol Rev. 2009;228:58–73. 7. Weber AN, Bittner Z, Liu X, et al. Bruton’s tyrosine kinase: an emerging key player in innate immunity. Front Immunol. 2017;8:1454. 8. Xiao W, Mindrinos MN, Seok J, et al. A genomic storm in critically injured humans. J Exp Med. 2011;208:2581–2590. 9. O’Riordan CE, Purvis GSD, Collotta D, et al. Bruton’s tyrosine kinase inhibition attenuates the cardiac dysfunction caused by cecal ligation and puncture in mice. Front Immunol. 2019;10:2129. 10. O’Riordan CE, Purvis GSD, Collotta D, et al. X-linked immunodeficient mice with no functional Bruton's tyrosine kinase are protected from sepsis-induced multiple organ failure. Front Immunol. 2020;11:581758. 11. Liu SF, Malik AB. NF-κB activation as a pathological mechanism of septic shock and inflammation. Am J Physiol Lung Cell Mol Physiol. 2006; 290:L622–L645. 12. Coldewey SM, Rogazzo M, Collino M, et al. Inhibition of IkB kinase reduces the multiple organ dysfunction caused by sepsis in the mouse. Dis Model Mech. 2013;6:1031. 13. Sordi R, Chiazza F, Johnson FL, et al. Inhibition of IkB kinase attenuates the organ injury and dysfunction associated with hemorrhagic shock. Mol Med. 2015;21:563–575. 14. Nicolson PLR, Welsh JD, Chauhan A, et al. A rationale for blocking thromboin-flammation in COVID-19 with Btk inhibitors. Platelets. 2020;31:685–690. 15. Roschewski M, Lionakis MS, Sharman JP, et al. Inhibition of Bruton tyrosine kinase in patients with severe COVID-19. Sci Immunol. 2020;5:eabd0110. 16. Soresina A, Moratto D, Marco Chiarini, et al. Two X-linked agammaglobu-linemia patients develop pneumonia as COVID-19 manifestation but recover. Pediatr Allergy Immunol. 2020;31:565–569. 17. Burger JA. Bruton tyrosine kinase inhibitors: present and future. Cancer J. 2019;25:386–393. 18. Liu X, Zhang J, Han W, et al. Inhibition of BTK protects lungs from trauma-hemorrhagic shock-induced injury in rats. Mol Med Rep. 2017;16:192–200. 19. Sordi R, Chiazza F, Collotta D, et al. Resolvin D1 Attenuates the Organ Injury Associated With Experimental Hemorrhagic Shock. Ann Surg. 2021;273:1012–1021. 20. Sordi R, Nandra KK, Chiazza F, et al. Artesunate protects against the organ injury and dysfunction induced by severe hemorrhage and resuscitation. Ann Surg. 2017;265:408–417. 21. Yamada N, Martin LB, Zechendorf E, et al. Novel synthetic, host-defense peptide protects against organ injury/dysfunction in a rat model of severe hemorrhagic shock. Ann Surg. 2018;268:348–356. 22. Martin L, Patel NM, et al. The inhibition of macrophage migration inhibito
留言 (0)