
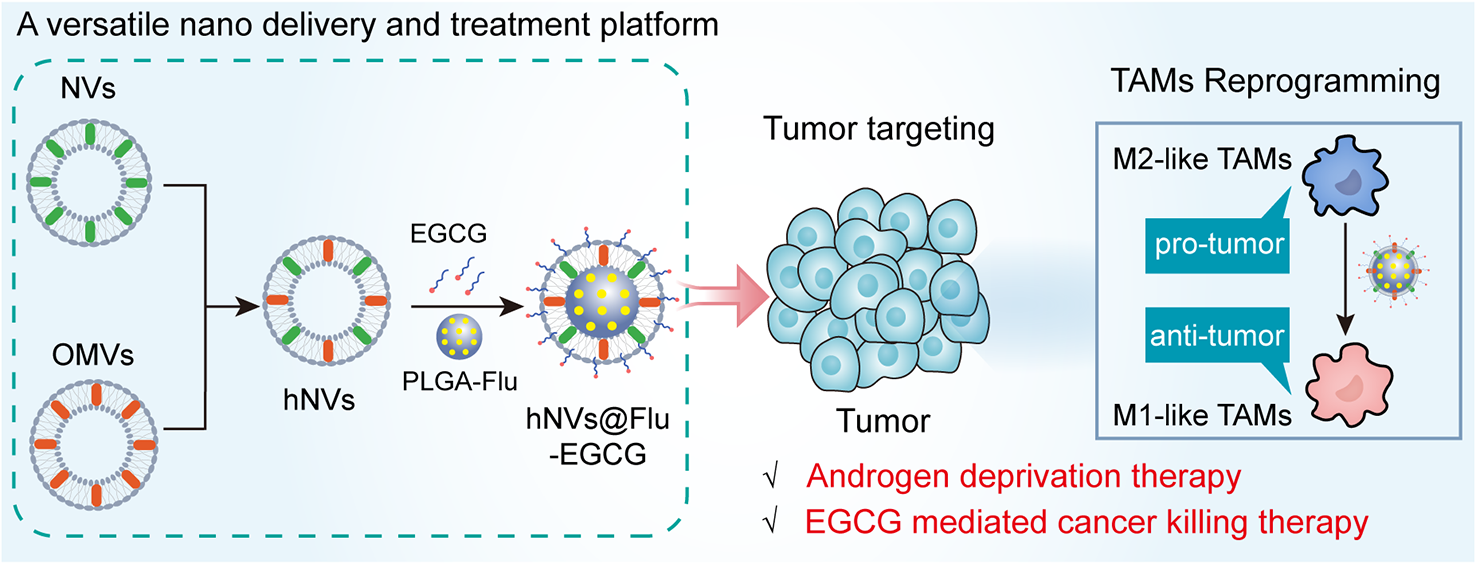
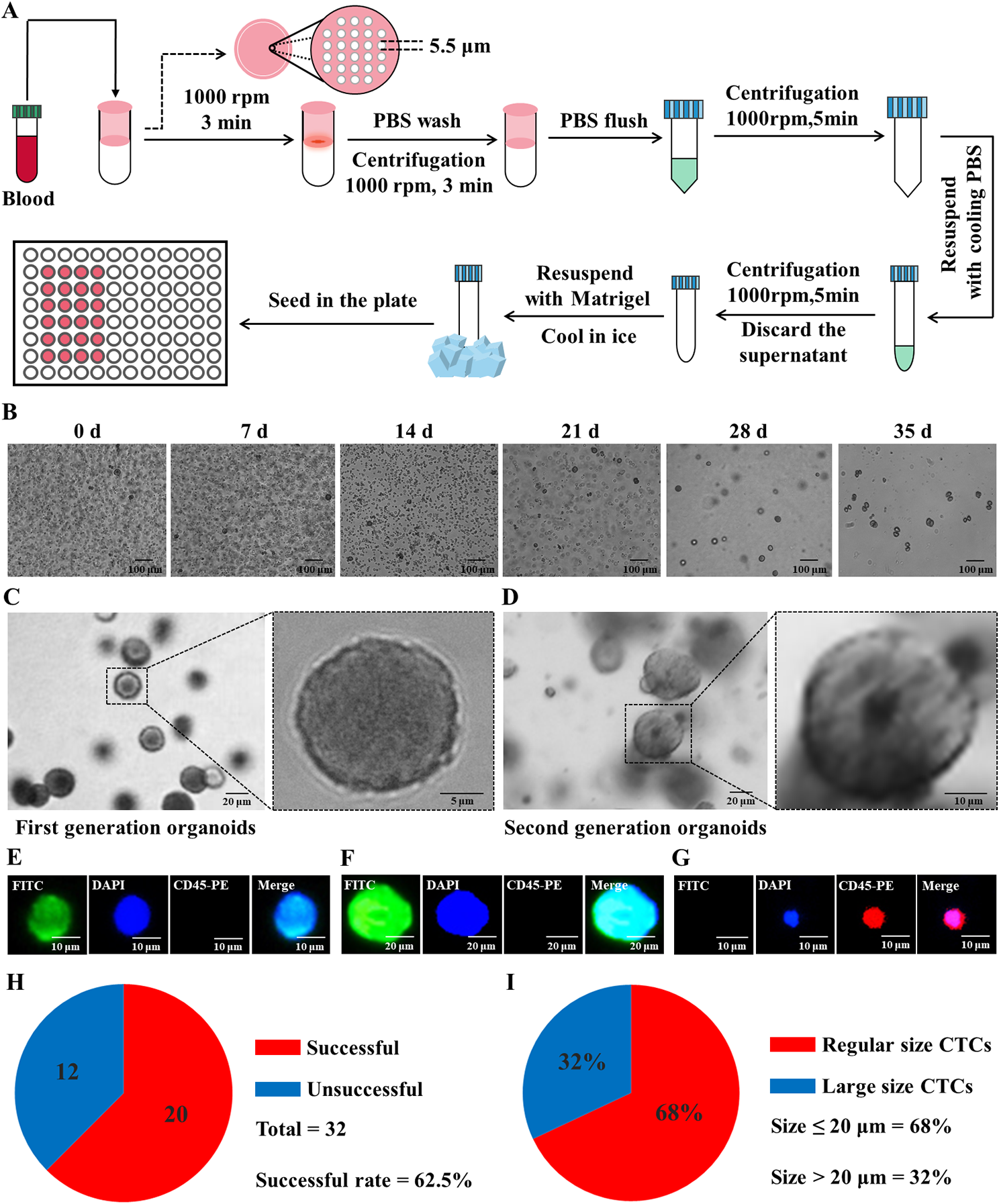
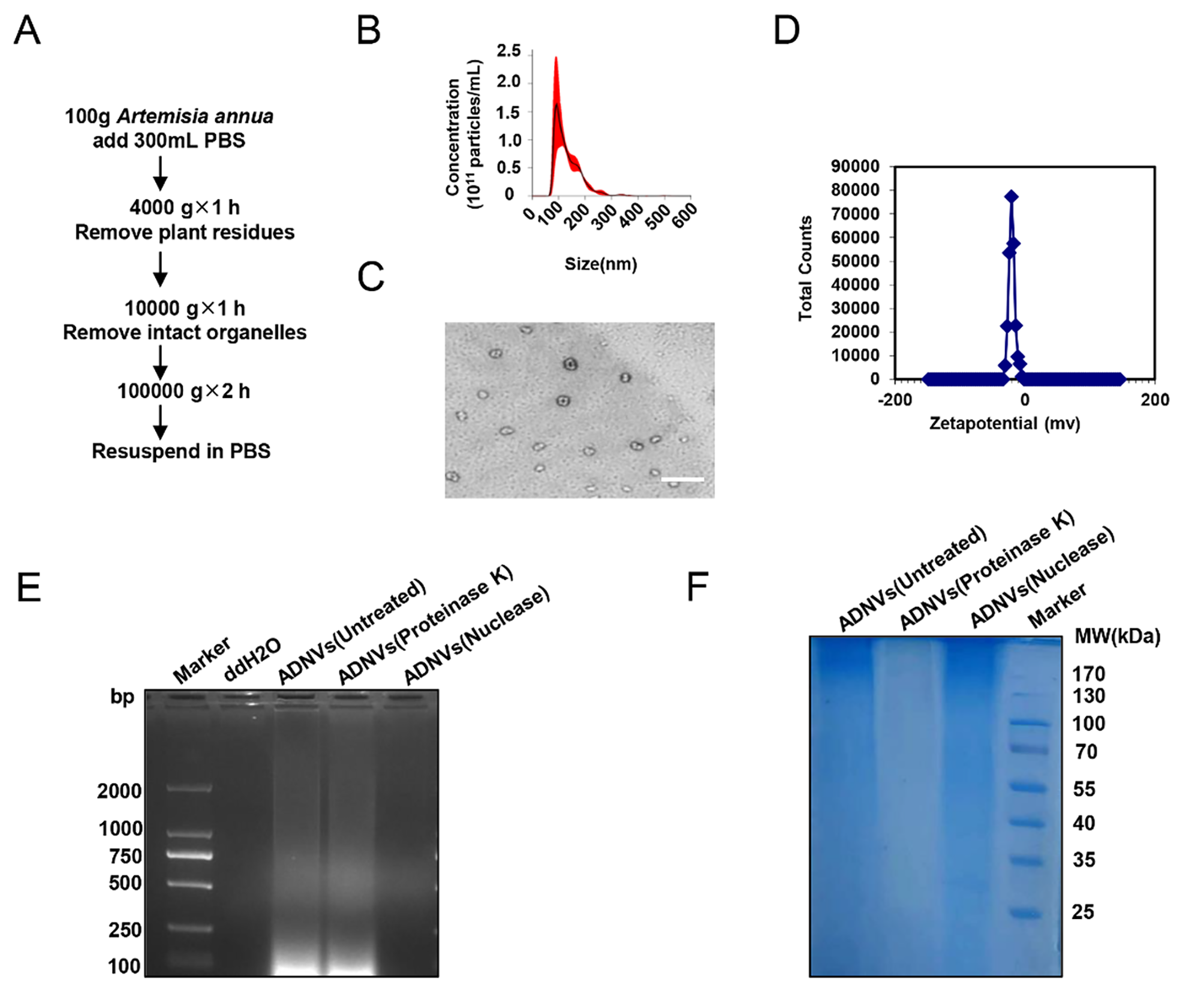
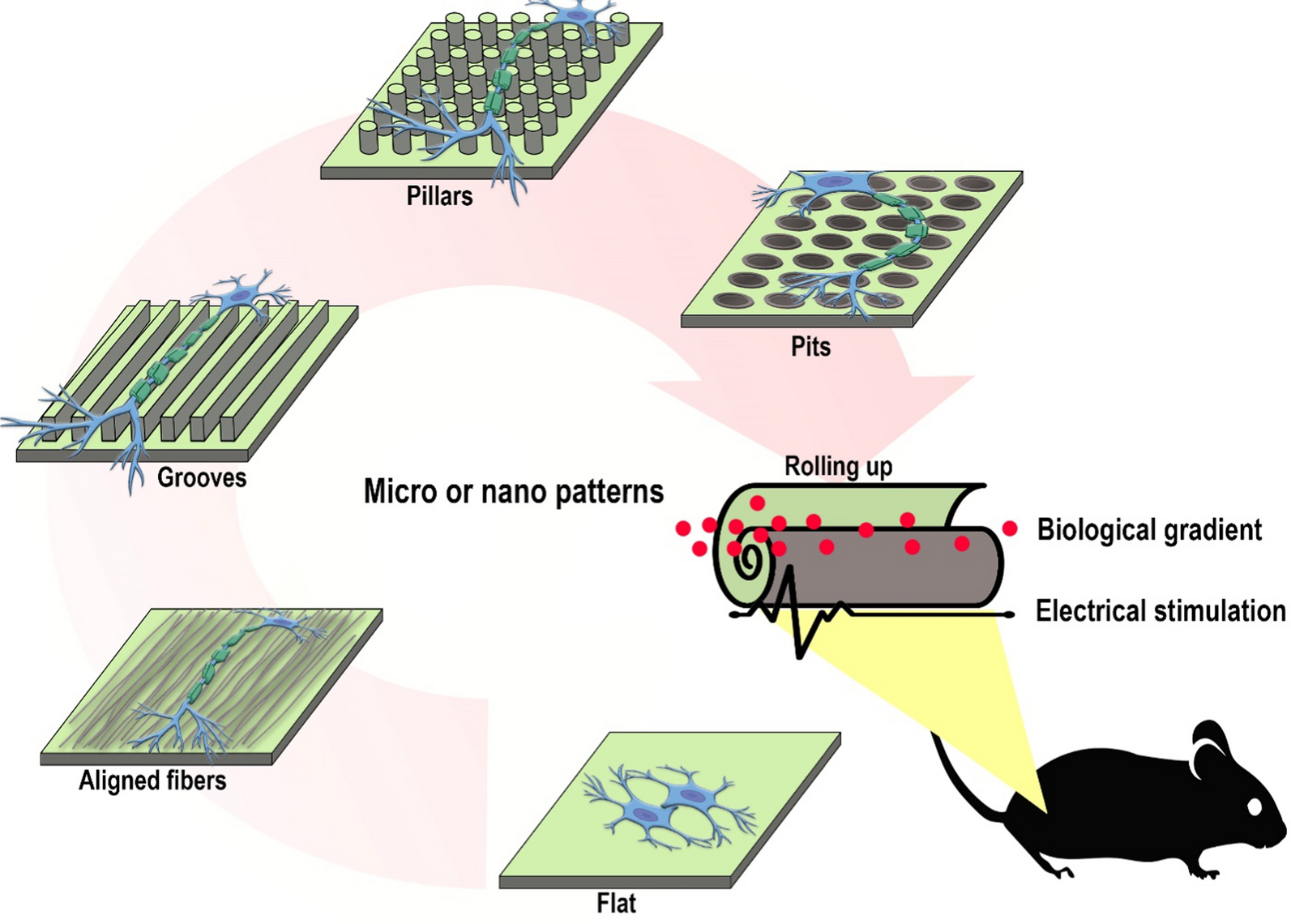
記住我
The drug loading rate of BBR-loaded PLGA nanoparticles (BBR@PLGA NPs), as determined by liquid chromatography-mass spectrometry, was 10.07%, which confirmed that BBR@PLGA NPs were successfully prepared by a double emulsion process. As determined by transmission electron microscopy, BBR@PLGA NPs had a spherical morphology, with an average diameter of approximately 210 nm, BBR@PLGA@PLT NPs had a “core-shell” structured morphology (Fig. 1a), the single outer layer of the PLT “shell” was approximately 10 nm thick, consistent with the previously reported thickness of PLTs [30]. Dynamic light scattering analysis indicated that the peak hydrodynamic diameter of BBR@PLGA NP was 215 nm, with a polydispersity index of 0.161 and a zeta potential of − 10.8 mV (Fig. 1b, c). Compared with uncoated BBR@PLGA NP, the peak hydrodynamic diameter of BBR@PLGA@PLT NP was increased from 215 to 235 nm, which was attributed to the PLTs. Additionally, the absolute value of zeta potential of BBR@PLGA@PLT NP (− 11.4 mV) was higher than that of unmodified BBR@PLGA NP (Fig. 1b, c). Subsequently, the stability of BBR@PLGA@PLT NPs was tested, and no significant change was found in the zeta potential or nanoparticle size within eight days (Fig. 1d).
Fig. 1
Characterization of the platelet (PLT) membrane-coated biomimetic nanoparticles and ex vivo fluorescence imaging of isolated major organs. a Transmission electron microscopy images of BBR@PLGA and BBR@PLGA@PLT NPs (scale bar = 200 nm). b The sizes and c zeta potentials of BBR@PLGA and BBR@PLGA@PLT NPs. d The trends of sizes and zeta potentials of BBR@PLGA@PLT NPs over time. e In vitro drug release profiles of BBR@PLGA and BBR@PLGA@PLT NPs. f Ex vivo fluorescence imaging of the heart and the other major organs (liver, kidney, spleen, and lung) of MI rats 24 h after DSPE-PEG2000-Cy7-labelled BBR@PLGA@PLT administration. All bars represent as means ± SD (n = 3)
The in vitro release kinetics of BBR@PLGA and BBR@PLGA@PLT NPs are shown in Fig. 1e. PLT-coated nanoparticles had a smoother release curve. In the BBR@PLGA group, the drug was markedly released during the first four days of the experiment, and 79.83% ± 3.67% of BBR was released from day 0 to day 10. In contrast, drug release was relatively stable in the BBR@PLGA@PLT group, with BBR release of 34.90% ± 3.69% from day 0 to day 10. From the 10th to the 20th day, the drug release ability of the BBR@PLGA group was significantly decreased, with a drug release amount of 16.01% ± 1.04%. In contrast, in the BBR@PLGA@PLT group, 37.42% ± 0.68% of the drug was released. After 20 days of incubation, the BBR@PLGA and BBR@PLGA@PLT groups released 95.83% ± 3.09% and 74.32% ± 5.32% of the BBR, respectively. One of the disadvantages of PLGA nanoparticles is that initial drug burst release is common [31]. Both membrane-like nanoparticles and PLGA nanoparticles exhibit sustained drug release properties in vitro [32]. In the BBR@PLGA@PLT system, BBR@PLGA is coated by PLT membrane with a double sustained release effect, and the drug released from BBR@PLGA nanoparticles remains in the PLT membrane first and does not rapidly release outside of BBR@PLGA@PLT, which may be the reason why the drug release curve of BBR@PLGA@PLT is smoother than that of BBR@PLGA. This indicates PLTs could help in the delivery of drugs released early from BBR@PLGA nanoparticles to the infarcted myocardium, which may have a beneficial impact on treatment.
Distearoyl phosphatidylethanolamine (DSPE), as a hydrophobic group, can be inserted into the platelet membrane because of its excellent biological scalability. Therefore, DSPE-PEG2000-Cy7 was used to label BBR@PLGA@PLT fluorescently. Figure 1f shows the distribution of BBR@PLGA@PLT in the major organs of rats with acute MI 24 h after intravenous injection of nanoparticles. The fluorescence intensity in the infarcted myocardium is significantly higher than that in the non-infarcted area. As a control experiment, Cy7-encapsulated PLGA nanoparticles were administered to MI rats via the caudal vein. PLGA nanoparticles without platelet encapsulation had no immune evasion ability, so the liver captured most PLGA nanoparticles, and there was no difference in fluorescence intensity between the infarcted myocardium and the background (Additional file 1: Fig. S1).
The drug release kinetics and biodistribution results suggest that BBR@PLGA@PLT NPs have ability to remain in circulation for a long time and reduce the potential to be trapped in the liver, which allows more nanoparticles to be enriched in the infarcted myocardium. The PLT coating technology provides BBR@PLGA NPs with the ability to escape the immune response and become enriched in damaged vessels and inflammatory lesions [33]. In addition, PLTs have good biological expansibility and can be further modified. BBR@PLGA@PLT NPs can facilitate the targeted delivery of anti-inflammatory drugs such as BBR into infarcted myocardium.
Network pharmacologic analysis of the effect of BBR on MIIn traditional Chinese medicine, because the therapeutic mechanism of natural drugs is extremely complex, compounds composed of natural drugs are often used to treat patients through network interactions [34]. The most important mechanism of BBR in the treatment of MI is difficult to confirm by quantitative analysis in animal experiments. Network pharmacology can be used to predict the mechanism, toxicity, and metabolic characteristics of traditional Chinese medicines [35]. Therefore, we conducted a network pharmacological analysis of the mechanism of BBR action in acute MI. To determine their interactions, 289 target genes associated with BBR and 1837 target genes associated with acute MI were analyzed. A protein-protein interaction network was constructed using 200 different genes considered key targets in acute MI treatment (Fig. 2a), which indicated that the core mechanisms of BBR effect on MI involve centrally located interleukin (IL)-1β, TNF, and nuclear factor kappa-B, etc.
Fig. 2
Interaction network of the overlapping targets. a Protein–protein interaction network of the overlapping targets. b Gene Ontology enrichment analysis of the overlapping targets
Gene Ontology enrichment analysis was performed to reveal the biological characteristics of 200 intersecting target genes. Detailed network pharmacology data are provided in Additional file 1: Fig. S2–S4. The top 20 terms significantly enriched for biological processes (Fig. 2b) included negative regulation of the apoptotic process, positive regulation of the apoptotic process, positive regulation of cell proliferation, inflammatory response, and response to hypoxia, which were all closely associated with the treatment of MI. Therefore, we speculated that BBR exerted a protective effect on the myocardium through the above biological processes. Because the effects of natural drugs are often multifaceted, we excluded terms that were in conflict and those appeared as outcomes, we concluded that the therapeutic effect of BBR on MI mainly involves the inflammatory response and hypoxia response. The enrichment score of the inflammatory response was much higher than that of the hypoxia response; therefore, we primarily verified the effect of BBR@PLGA@PLT NPs on the inflammatory response.
Ability of BBR@PLGA@PLT NPs to regulate inflammation in vitro and in vivoSubsequently, we evaluated the in vitro biological effects of BBR@PLGA@PLT NPs. The cytotoxicity of BBR@PLGA@PLT NPs was investigated in macrophages (Fig. 3a). In both the BBR@PLGA group and BBR@PLGA@PLT group, BBR was coated with nanoparticles and entered the macrophage environment in a sustained release manner. Therefore, the BBR concentration in the cell environment was relatively low in the BBR@PLGA and BBR@PLGA@PLT groups compared with that in the BBR aqueous solution group. The cell viability decreased only slightly with increasing BBR concentration. Only the BBR aqueous solution groups that contained 50 μg/mL and 100 μg/mL of BBR showed significant reductions in the viability of RAW 264.7 cells (P = 0.0304, P = 0.0062). These results demonstrated that the sustained release ability of BBR@PLGA and BBR@PLGA@PLT NPs reduced the toxicity of BBR and that PLGA and PLTs are biocompatible.
Fig. 3
The in vitro and in vivo effects of BBR@PLGA@PLT NPs on macrophages. a Cell viability of macrophages after incubation with various concentrations of BBR, BBR@PLGA, and BBR@PLGA@PLT NPs in solution for 12 h. b Confocal microscopic images of macrophage markers after incubation in 10 μM BBR solution and saline for 12 h. c Inflammatory cytokines secreted by RAW 264.7 cells. d Serum levels of inflammatory cytokines in rats on day three after MI. e Representative immunofluorescence images of TUNEL, CD206, and CD86 in the border zone of infarcted hearts on day three after MI. TUNEL-stained (green) cells indicate apoptosis-positive cells, and CD86 (red) and CD206 (green) staining shows M1 and M2-type macrophages, respectively. f Statistical analysis of the percentage of TUNEL-, CD206-, and CD86-positive cells. All bars represent as means ± SD (n = 3). *P < 0.05 and **P < 0.01, ***P < 0.001
As shown in Fig. 3b, the BBR aqueous solution group showed significant up-regulation of ARG1 and CD206 (M2 subtype marker) in RAW 264.7 cells compared with the saline control group. We demonstrated that 10 μM BBR did not affect macrophage viability (Fig. 3a), suggesting that BBR exerts its effect on macrophages mainly by acting on macrophage M2-subtype polarization rather than directly inhibiting macrophage activity.
The normalized levels of inflammatory cytokines secreted by RAW 264.7 cells are shown in Fig. 3c (the non-normalized data are shown in Additional file 1: Fig. S5). Only the BBR solution group showed significant differences, with down-regulation of IL-6 (a proinflammatory factor) and TNF-α (a major cytokine that mediates cardiomyocyte apoptosis by inflammation) secretion and up-regulation of IL-10 (an anti-inflammatory factor) secretion. These data again demonstrated that BBR was released from BBR@PLGA and BBR@PLGA@PLT NPs in a sustained manner, and PLGA and PLTs did not produce changes in macrophage secretion in vitro.
In vivo experiments, the serum levels of IL-10, IL-6, and TNF-α were increased on day four after MI, and the IL-10 level was further increased after BBR@PLGA and BBR@PLGA@PLT treatment, while the TNF-α and IL-6 levels were decreased. Moreover, the BBR@PLGA@PLT group had the highest serum IL-10 level and the lowest IL-6 and TNF levels (all P < 0.05) (Fig. 3d, the non-normalized data are shown in Additional file 1: Fig. S5).
Thus, we reasoned that the most likely mechanism of action of BBR@PLGA@PLT NPs on MI involved the cardiac enrichment capacity of the PLT coating, the sustained BBR release capacity of PLGA nanoparticles, and the ability of BBR to promote M2 polarization in macrophages.
We demonstrated that BBR@PLGA@PLT NPs could promote M2 polarization of macrophages in infarcted myocardium by measuring the expression of markers, including TUNEL (apoptotic marker), CD206 (M2 macrophage surface marker), and CD86 (M1 macrophage surface marker) on day four post-MI (Fig. 3e). There was no significant difference in the percentage of TUNEL-positive cells (apoptotic cardiomyocytes) and CD206-positive cells between the saline group and BBR@PLGA group (P = 0.0683, P = 0.2088). The only significant difference between these groups was in the percentage of CD86-positive cells (P = 0.0328). The BBR@PLGA@PLT group showed significant differences in all markers compared with the BBR@PLGA group. Apoptotic cardiomyocytes (P = 0.0018) and CD86-positive cells (P = 0.0025) were significantly reduced, and CD206-positive cells were increased significantly in the BBR@PLGA@PLT group compared with the BBR@PLGA group (P = 0.0062) (Fig. 3f).
Evaluation of cardiac function by echocardiographyCardiac function is a direct response to the therapeutic effect of drugs on MI, and was measured by echocardiography in rat models on day 28 after MI. Representative echocardiograms are shown in Fig. 4a. Compared with the sham group, the MI rats treated with saline exhibited typical characteristics of heart failure with a significantly increased left ventricular internal diameter at end-diastole (LVIDd), left ventricular inner diameter at end-systole (LVIDs), end-systolic volume (ESV), and end-diastolic volume (EDV), as well as reductions in left ventricle fractional shortening (FS) and left ventricular ejection fraction (LVEF) (all P < 0.05) (Fig. 4b–g). BBR@PLGA NPs demonstrated a limited protective effect on heart function with increased EF and FS and decreased in LVIDd, LVIDs, EDV, and ESV compared with saline (all P < 0.05). The BBR@PLGA@PLT group had the highest EF and FS and the lowest LVIDd, LVIDs, EDV, and ESV among all MI groups (all P < 0.05) (Fig. 4b–g).
Fig. 4
Cardiac function was evaluated by echocardiography on day 28 after MI. a Representative echocardiography images of different groups. b LVIDs. c LVIDd. d FS. e ESV. f EDV. g LVEF. All bars represent as means ± SD (n = 6). *P < 0.05 and **P < 0.01, ***P < 0.001
Evaluation of fibrosis by pathological staining and western blottingMyocardial fibrosis is an important pathological process that leads to adverse cardiac remodeling and fatal heart failure [36, 37]. The pathological manifestation of fibrosis is that excess collagen deposition replaces normal tissue. Systematic assessment of myocardial fibrosis on day 28 showed that BBR@PLGA@PLT NPs reduced collagen accumulation and improved collagen composition (Fig. 5).
Fig. 5
Evaluation and quantitative analysis of fibrosis by pathological staining and western blotting on day 28 after MI. a Representative images of heart sections stained with Masson trichrome, collagen is stained blue. b Representative images of heart sections stained with Sirius Red, the red and yellow tissues are considered type I collagen, while the green tissues are type III collagen. Quantitative analysis of the c ventricular wall thickness, d collagen content, and e collagen I/III ratio in different groups was performed by visual analysis. f Western blot results, g quantification of collagen content, and h collagen I/III ratio. All bars represent as means ± SD (n = 3). *P < 0.05 and **P < 0.01, ***P < 0.001
The saline group had a large amount of collagen deposited in the ventricle, which was stained blue by Masson staining (Fig. 5a). Stiff collagen lacks the contractile capacity of cardiomyocytes and reduces ventricular thickness during the long-term passive stretch (Fig. 5c). Compared with the saline group, the BBR@PLGA group had greater ventricular thickness and less collagen deposition. The ventricles of the BBR@PLGA@PLT group had the thickest ventricular wall and the least collagen deposition among all MI groups (Fig. 5d).
The collagen fibers observed by Masson staining were composed of multiple components. Type III collagen has good elasticity, and type I collagen is stiff [37]. The collagen I/III ratio can reflect the passive expansion ability of collagen fibers in scar tissue. In this study, type I and type III collagen were observed using Sirius Red staining. The visual analysis results showed that the BBR@PLGA group had a lower collagen I/III ratio than the saline group (P = 0.0267), and the BBR@PLGA@PLT group had a significantly lower collagen I/III ratio than the BBR@PLGA group (P = 0.022) (Fig. 5e). To ensure the reliability of this conclusion, we also performed western blotting analysis of collagen in the infarcted myocardium (Fig. 5f). The densitometry results are presented as arbitrary units and normalized ratios. Semi-quantitative analysis showed that treatment with BBR@PLGA and BBR@PLGA@PLT NPs significantly reduced the content of type I and type III collagen in the scar tissue on day 28 (all P < 0.05). The BBR@PLGA@PLT group exhibited a greater reduction in type I collagen than the BBR@PLGA group (P = 0.0003), but there was no significant difference in the type III collagen content between the two groups (P = 0.8078) (Fig. 5g). The BBR@PLGA@PLT group had the lowest type I/III collagen ratio, which was significantly lower than that in the BBR@PLGA group (P = 0.0031) (Fig. 5h). Thus, we again verified the effect of BBR@PLGA@PLT NPs on improving the composition of scar tissue by western blot analysis. The elasticity of collagen fibers in scar tissue plays an important role in the protection of cardiac function.
Fibroblasts secrete collagen in the myocardium and other organs. Studies [37, 38] have shown that the secretion of collagen by fibroblasts in infarcted myocardium is mainly regulated by inflammatory factors, such as IL-10. IL-10 increases collagen synthesis in the early phase of MI, but inhibits collagen synthesis and regulates collagen composition in the later phase of MI, leading to the up-regulation of type I collagen in scar tissue. Type I collagen, which has greater elasticity than type III collagen, can reduce fibrotic myocardial stiffness and directly affect cardiac function. Importantly, cardiac stiffness is one of the leading causes of decompensation in heart failure [39]. Therefore, we suggest that BBR@PLGA@PLT NPs can be enriched in the infarcted myocardium, and the released BBR can regulate the secretory phenotype of macrophages to reduce fibrosis of the infarcted myocardium and stiffness of cardiac scar tissue.
Assessment of cardiac structuresCardiac structures, including electrical signal transmission networks, vascular networks, and cardiomyocytes [40], were assessed on day 28 by immunofluorescence analysis.
Myocardial structure damage was detected by measuring α-actinin and CX43 protein levels. α-actinin is a vital protein of the myocardial skeleton, and CX43 mediates electrical signal transmission between cardiomyocytes. As shown in Fig. 6a, MI operation significantly changed the myocardium structure. CX43 and α-actinin were significantly down-regulated in the infarcted myocardium of the saline group. Additionally, the location of CX43 in the tissue was altered; CX43 was absent from viable myocardial tissue and mainly distributed around the nuclei of cardiomyocytes. Compared with the saline group, both the BBR@PLGA and BBR@PLGA@PLT groups had CX43 in the correct location. Furthermore, CX43 and α-actinin protein levels were up-regulated in the BBR@PLGA and BBR@PLGA@PLT groups, with greater up-regulation in the BBR@PLGA@PLT group than that in the BBR@PLGA group (all P < 0.0001) (Fig. 6b).
Fig. 6
Assessment of cardiac structures, including electrical signal transmission networks, vascular networks, and cardiomyocytes on day 28 after MI. a Representative immunofluorescence images co-stained for CX43 (green) and α-actinin (red). b Optical density semi-quantitative analysis of CX43 and α-actinin expression. c Representative immunofluorescence images of blood vessels co-stained for CD31 (green) and α-SMA (red). d Quantitative analysis of the neovessels in the infarct regions. e Evaluation of cardiomyocyte apoptosis in the border zone of infarcted hearts by TUNEL staining and f semi-quantitative analysis. g Western blot and h quantification of TNF-α expression in infarcted myocardium. All bars represent as means ± SD (n = 3). *P < 0.05 and **P < 0.01, ***P < 0.001
In infarcted myocardium, cardiomyocytes are more likely to survive when angiogenesis occurs [41]. As shown in Fig. 6c, d, compared with the sham group, the saline group showed angiogenesis induced by MI operation. The vascular area of the infarcted myocardium in the saline group was significantly greater than that of the normal myocardium in the sham group (P = 0.0088). The BBR@PLGA group exhibited more vascularization than the saline group (P = 0.0391), and the angiogenesis of infarcted myocardium in the BBR@PLGA@PLT group was significantly greater than that in the BBR@PLGA group (P = 0.0162).
As shown in Fig. 6e, f, compared with the sham group, all MI groups showed obvious cardiomyocyte apoptosis. The number of apoptotic cells in the BBR@PLGA group was significantly lower than that in the saline group (P = 0.0304), while the number of apoptotic cells in the BBR@PLGA@PLT group was lower than that in the BBR@PLGA group (P = 0.0169). TNF-α is produced predominantly by macrophages, and it is a major cytokine responsible for cardiomyocyte apoptosis [42]. Western blotting was performed to evaluate the TNF-α content in infarcted myocardium (Fig. 6g). The BBR@PLGA@PLT group had the least TNF-α in the infarcted myocardium (all P < 0.05), consistent with TUNEL staining.
BBR@PLGA@PLT NPs can protect the infarcted myocardium through various mechanisms by regulating inflammatory responses, such as down-regulation of TNF-α secreted by macrophages directly involved in reducing cardiomyocyte apoptosis [43]. Moreover, M1-type macrophages have been suggested to mainly promote vessel sprouting, while M2-type macrophages mainly promote the maturation and quiescence of new blood vessels. M1-type macrophages can cause degeneration of previously formed vessels, suggesting that promoting polarization of inflammatory macrophages to the M2 subtype after MI is critical for angiogenesis [4,5,6,7]. The BBR@PLGA@PLT group formed more mature arterioles consisting of CD31-positive cells (endotheliocytes) surrounded by α-smooth muscle actin (α-SMA)-positive cells (smooth muscle cells), demonstrating the ability of BBR@PLGA@PLT NPs to protect cardiomyocytes by resolving inflammation and up-regulating angiogenesis.
Biosafety assessmentThe toxic side effects of nanoparticles on major organs and whole systems are a major concern during drug treatment. Potential side effects were investigated to assess biosafety in vivo. Biochemical analysis showed no significant differences in creatinine (Cr) and blood urea nitrogen (BUN) levels between any two groups, indicating that renal function was not affected by BBR@PLGA@PLT NPs treatment (Fig. 7a, b).
Fig. 7
Preliminary safety evaluation. a–d Biochemical markers relevant to hepatic and kidney function. e Hematoxylin and eosin staining of sections of major organs resected from mice and subjected to treatment with various formulations for one month. All bars represent as means ± SD (n = 6)
The serum alanine aminotransferase (ALT) and aspartate aminotransferase (AST) levels are commonly used to assess the toxicity of a drug to the liver and heart, respectively [44]. Compared with the saline group, the serum ALT level decreased after BBR@PLGA and BBR@PLGA@PLT NPs treatment (P < 0.0001), and the decrease was more obvious in the BBR@PLGA@PLT group (Fig. 7c), indicating that both BBR@PLGA and BBR@PLGA@PLT NPs had a protective effect on MI in rats, but the effect of BBR@PLGA@PLT NPs was superior to that of BBR@PLGA NPs. The BBR@PLGA@PLT group had the lowest serum ALT level among all MI groups (P < 0.0001, P = 0.0025) (Fig. 7d). However, in a comparison of the sham and saline groups, we concluded that MI operation resulted in increased serum AST and ALT levels in rats on day 28 (all P < 0.0001). Myocardial injury also causes an increase in the serum AST level; therefore, it is difficult to evaluate whether the drug injures the liver or has a protective effect via the serum AST level. Consequently, we performed a histological evaluation of the major organs (Fig. 7e). Hematoxylin and eosin staining showed no obvious histologic damage in the hearts, livers, spleens, lungs, or kidneys isolated from the BBR@PLGA@PLT group. TUNEL staining on liver sections showed that hepatocytes in each group exhibited no obvious apoptosis. All of the above results indicated that no obvious side effects occurred in other major organs.
留言 (0)