
記住我


By using endogenous AGO1, the in vitro RISC assay can eliminate the limitation of plant material; however, a good-quality α-AGO1 antibody that does not affect AGO1 function would determine whether the in vitro RISC assay can be successfully performed. Although commercial AGO1 is available, we made a homemade α-AGO1 antibody to pull down a large amount of AGO1 for the assay. Moreover, the generation knowledge can be applied to producing other AGO antibodies, e.g., AGO2.
The amino acid alignments of AGO1, AGO2, AGO4, and AGO10 showed that the 1-to-256-aa region is variable, with 12.2%-30.5% identity (Fig. 1a). Thus, the N-terminus of AGO1 (1–240 aa) was selected for recombinant protein production and purified as an antigen to the AGO1 antibody (Fig. 1a, box). Western blot analysis showed that the α-AGO1 antibody could recognize endogenous AGO1 (117 kDa) in Col-0 plants. In contrast, the AGO1 signal was absent in the ago1-36 null mutant, suggesting that the homemade α-AGO1 antibody can be tested for its IP ability with an in vitro RISC assay (Fig. 1b).
Fig. 1
α-AGO1 antibody generation. a Amino acid alignment of the N-termini of AGO1, AGO2, AGO4, and AGO10. The gray box indicates the N-terminal region of AGO1 for antigen. b Evaluation of the α-AGO1 antibody for detection of endogenous AGO1 in Col-0 plants. The null ago1-36 mutant serves as the negative control. The asterisk indicates tubulin, which was used as a loading control
To evaluate the in vitro RISC assay with the α-AGO1 antibody, endogenous AGO1 was immunoprecipitated from Col-0 seedlings and ago1-27 mutants and assessed with the RNA substrate MYB33 (MYB33-230), which contained a target site for miR159a and miR159b (Fig. 2a). The AGO1-IP from Col-0 seedlings can target and cleave MYB33-230 (Fig. 2b). The signals of 5'- and 3'-cleaved fragments were increased when the amounts of AGO1-IP were increased, suggesting that AGO1 cleavage is dose-dependent (Fig. 2b). The ago1-27 mutant, which exhibits an Ala992Val weak allele, showed attenuated slicer activity [24]. AGO1-IP from the ago1-27 mutant revealed inefficient MYB33-230 cleavage (Fig. 2b). The amounts of AGO1-IP from Col-0 plants and ago1-27 mutants were comparable (Fig. 2c). The 20 ng AGO1-IP from Col-0 plants resulted in 81% cleavage efficiency, whereas only 37% cleavage efficiency was observed with 20 ng AGO1-IP from ago1-27 mutants (Fig. 2d). We further mutated the target site in the seed region (MYB33mSeed) and center region (MYB33mCenter) (Fig. 2e). The results showed no RISC cleavage of MYB33mSeed or MYB33mCenter substrates, whereas MYB33-230 was cleaved as a positive control (Fig. 2f). These results indicated that the α-AGO1 antibody could be used for the in vitro RISC assay.
Fig. 2
Establishment of an in vitro RISC assay. a Sequence pairing of MYB33 RNA and miR159 isoforms. The mismatched nucleotides are highlighted in red. b The endogenous AGO1-IP products from Col-0 plant and ago1-27 mutant were incubated with MYB33-230 RNA substrate to perform an in vitro RISC assay. The RNA substrate and cleaved RNA fragments are indicated with arrows. c The AGO1-IP amounts from Col-0 plant and ago1-27 mutant were evaluated by western blotting with α-AGO1 IgG for normalization of the RISC cleavage efficiency. In: input; IP: immunoprecipitation; FT: flow through. d Normalized RISC cleavage efficiency for AGO1-IP from Col-0 plant and ago1-27 mutant. e Sequencing pairing between miR159a and the MYB33mSeed or MYB33mCenter. The mismatched nucleotides are highlighted in red. f In vitro RISC assay for MYB33-230, MYB33mSeed, and MYB33mCenter RNA substrates. The RNA substrate and cleaved RNA fragments are indicated with arrows. (g) In vitro RISC assay for various miRNA-target substrates. h Normalized RISC cleavage efficiency for CSD-230, NAC1-230, and ARF16-230 RNA substrates (i). Normalized RISC cleavage efficiency for ARF10-230 and ARF10-191 RNA substrates (ii). The MYB33-230 RNA substrate served as a positive control
Next, we also examined different RNA substrates, including CSD2-230 (miR398 target), NAC1-230 (miR164 target), ARF16-230 (miR160 target), and ARF10-230 (miR160 targets) (Fig. 2g). All RNA substrates exhibit different levels of cleavage efficiency. For example, MYB33-230 and CSD2-230 have higher cleavage efficiency than NAC1-230, ARF16-230, and ARF10-230 (Fig. 2h, panel i). We designed two ARF10 substrates with 3'-end of different lengths: ARF10-230 (230 nt) and ARF10-191 (191 nt) (Fig. 2g). The data indicated that ARF10-191 exhibited better cleavage efficiency than ARF10-230 (Fig. 2h, panel ii). Thus, we assumed that the secondary structure of the RNA substrate and abundance of miRNA in AGO1 might affect the cleavage efficiency.
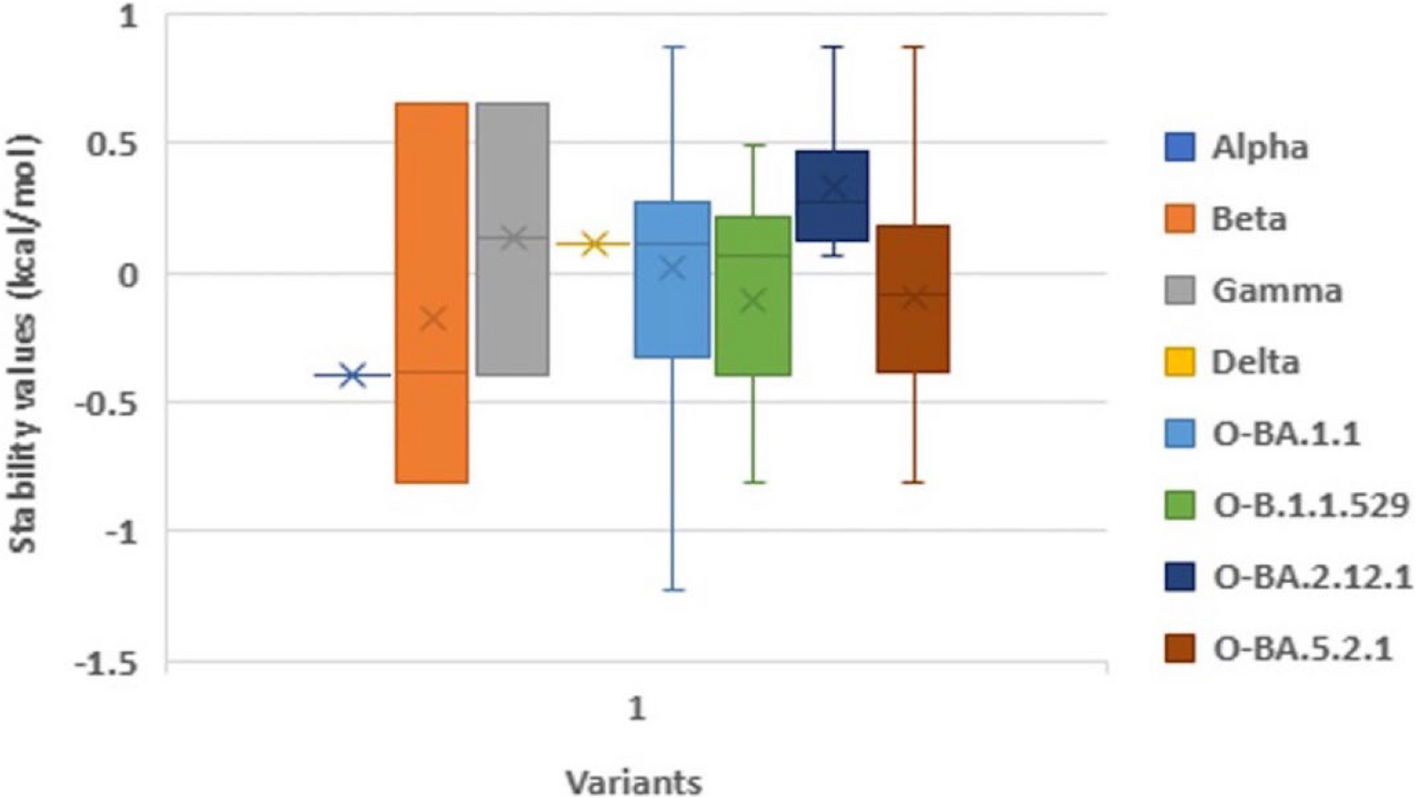
Evaluating endogenous RISC activity in P1/HC-Pro plantsOur previous study generated three viral species of P1/HC-Pro plants, including P1/HC-ProTu, P1/HC-ProZy, P1/HC-ProTe, and P1/HC-ProTu−K plants (Fig. 3a) [4, 8]. The P1/HC-ProTu, P1/HC-ProZy, and P1/HC-ProTe plants showed identical elliptical cotyledons (Fig. 3a, arrowheads). However, the cotyledons of P1/HC-ProTu−K plants were round in shape similar to those of Col-0 (Fig. 3a). We applied NGS to obtain the genomic sequences from these P1/HC-Pro plants. We also developed T-DNA Finder software to identify the T-DNA insertion based on the genomic sequence profiles. Two T-DNAs were identified on chromosome 1: P1/HC-ProTe (at the 16,780,881th nt) and P1/HC-ProTu (at the 28,814,559th nt) (Fig. 3b). In contrast, P1/HC-ProTu−K was inserted on chromosome 3 (at the 21,554,188th nt), whereas P1/HC-ProZy was inserted on chromosome 4 (at the 10,075,981th nt) (Fig. 3b). According to the TAIR database, several critical RNA silencing component genes, e.g., AGOs, DCLs, RDRs, HEN1, HYL1, SE, and ATG8a, were also labeled on diagrammatic chromosomes with these T-DNA insertions, which is helpful for the generation of relative mutations in these P1/HC-Pro plants by crossing or gene editing (Fig. 3b).
Fig. 3
The functional studies of HC-Pro for AGO1 regulation. a Phenotypes of P1/HC-Pro plants. The photographs were obtained from 14-day-old seedlings. Bar, 0.2 cm. b T-DNA insertions of various P1/HC-Pro plants. The diagrammatic chromosomes were labeled with the T-DNA insertion positions and critical RNA silencing component genes. Bar, 100,000 bp. c AGO1 levels in various P1/HC-Pro plants. RUBISCO (asterisk) and tubulin were used as loading controls. d Immunoprecipitation for evaluating the interactions of AGO1 with various HC-Pros. The asterisks indicate the heavy chain of IgG. Tubulin was used as a loading control
We performed western blotting to evaluate the endogenous AGO1 levels in various transgenic plants, including the L1 line and P1/HC-ProTu plants (Fig. 3c, panel i). Notably, the L1 line is a β-glucuronidase (GUS)-transgenic Arabidopsis plant that underwent sense-posttranscriptional gene silencing, and the T-DNA inserted on chromosome 3 (at the 2,350,388th nt) (Fig. 3b) [25]. The transgene of P1/HC-ProTu was transferred into the L1 line to generate P1/HC-ProTu plant [8]. The Col-0 and L1 lines showed identical endogenous AGO1 amounts, whereas P1/HC-ProTu plants showed lower levels of AGO1 while HC-Pro was presented (Fig. 3c, panel i).
Moreover, Hu et al. demonstrated that various P1/HC-Pro plants have different AGO1 levels in vivo [4]. Indeed, P1/HC-ProTu plants exhibited 0.5-fold AGO1 levels than Col-0 plants, whereas P1/HC-ProTu−K plants had 0.7-fold AGO1 levels, suggesting that HC-ProTu−K has a partial ability to trigger AGO1 degradation (Fig. 3c, panel ii). Notably, Hu et al. and Wei et al. demonstrated that low AGO1 protein levels are caused by autophagy instead of miR168-mediated RNA silencing. However, the AGO1 levels of the P1/HC-ProZy and P1/HC-ProTe plants were 0.7- to 0.9-fold of those found in the wild-type plants, suggesting that various species of HC-Pros have different AGO1 degradation efficiency (Fig. 3c, panel ii) [4, 11]. Therefore, we aimed to understand whether endogenous AGO1 activity could be affected by VSR through in vitro RISC evaluation.
Evaluation of the AGO1-HC-Pro interactionA conserved WG pair, which localizes near the FRNK motif, is identified in 113 potyviruses [26]. Pollari et al. demonstrated that HC-Pro of potato virus A (HC-ProPVA) physically interacts with AGO1 through transient expression with a cross-linked purification approach [26]. However, we performed AGO1-IP from various P1/HC-Pro plants with the α-AGO1 antibody. The IP results indicated that AGO1 could be detected in three P1/HC-Pro and Col-0 plants at the input and AGO1-IP samples (Fig. 3d). The immunoassay failed to detect HC-Pro in AGO1-IP samples from three P1/HC-Pro plants, suggesting that HC-Pro and AGO1 might not have a direct interaction but might exist a bridge protein to mediate an indirect interaction between HC-Pro and AGO1 (Fig. 3d).
HC-Pro-mediated RISC inhibition assayWe purified comparable amounts of AGO1-IP from the various transgenic plants for in vitro RISC assays to evaluate the status of AGO1 activity with MYB33-230 RNA substrates. The AGO1-IP from Col-0 seedlings exhibited 20% and 29% normalized cleavage efficiency in 10 ng and 20 ng AGO1-IP (Fig. 4a, panel i; and 4b). In contrast, although the AGO1-IP from the P1/HC-ProTu plants still had RISC activity, the activity was reduced to 6% and 13% in 10 ng and 20 ng AGO1-IP, respectively, suggesting interference with RISC regulation (Fig. 4a, panel i; and 4b). Surprisingly, the normalized in vitro RISC activity from P1/HC-ProTu/atg8age plants returned to 20% and 39% efficiency with 10 and 20 ng AGO1-IP, respectively, suggesting the restoration of RISC regulation (Fig. 4a, panel i; and 4b). Notably, the AGO1-IP from Col-0, P1/HC-ProTu, and P1/HC-ProTu/atg8age plants were comparable (Fig. 4a, panel ii).
Fig. 4
Evaluation of various HC-Pros in terms of their efficiency in inhibiting RISC activity in vitro. a In vitro RISC assay (i) and evaluation of the AGO1-IP efficiency (ii) for P1/HC-ProTu and P1/HC-ProTu/atg8age plants. In: input; IP: immunoprecipitation; FT: flow through. b Comparison of the in vitro RISC efficiency among the Col-0, P1/HC-ProTu, and P1/HC-ProTu/atg8age plants. c Represented in vitro RISC assay with the P1/HC-ProTu, P1/HC-ProZy, P1/HC-ProTe, and P1/HC-ProTu−K plants from 3 biological repeats. d Relative RISC cleavage efficiency of the P1/HC-ProTu, P1/HC-ProZy, and P1/HC-ProTe plants (i) and comparison of the RISC efficiency between Col-0 and P1/HC-ProTu−K plants (ii). The bars represent the standard deviations (n = 3). Means were compared by Tukey’s honestly significance test. Different letters above the bars indicate significant differences
To evaluate other HC-Pros inhibit RISC activity, the P1/HC-ProZy and P1/HC-ProTe plants were used to perform their RISC activity to compare with P1/HC-ProTu and Col-0 plants (Fig. 4c). The relative RISC cleavage efficiency of P1/HC-ProZy plants showed no difference from that of Col-0 but differed from that of P1/HC-ProTu and P1/HC-ProTe plants, suggesting various HC-Pro of potyviruses have different RISC cleavage inhibition abilities. (Fig. 4c and 4d, panel i). Notably, the P1/HC-ProTu−K sample did not inhibit RISC activity, suggesting FKNK mutation lost HC-ProTu-specific RISC inhibition ability (Fig. 4c and 4d, panel ii).
TuMV-infected Arabidopsis triggers endogenous AGO1 degradationWe subsequently evaluated the endogenous AGO1 in TuMV-infected Col-0 plants (Fig. 5a). TuGR infection caused symptoms with mosaic and up-curling systemic leaves, whereas TuGK-infected plants showed symptomless (Fig. 5a). Similar to P1/HC-ProTu plants, TuGR-infected Col-0 plants showed lower amounts of endogenous AGO1 than mock or TuGK-infected Col-0 plants (Fig. 5b). High abundances of HC-Pro and CP were detected in TuGR-infected plants, whereas lower amounts of HC-Pro and CP were detected in TuGK-infected Col-0 plants, which demonstrated uneven HC-ProTu and HC-ProTu−K amounts in infected tissues. Similarly, miRNA regulation suppression was also observed in TuGR-infected Col-0 plants. More than twofold higher levels of AGO1 (miR168), ARF16 (miR160), and MYB33 (miR159) were also detected in TuGR-infected Col-0 plants (Fig. 5c). In contrast, TuGK infection induces ARF16 accumulation to approximately 1.5-fold levels, whereas the RNA levels for AGO1 and MYB33 were similar to those obtained with the mock plants (Fig. 5c). These data indicated that miRNA regulation in TuGR-infected plants was also suppressed.
Fig. 5
The AGO1 levels and evaluation of miRNA-mediated target cleavage in TuMV-infected plants. a Symptoms of TuGR- and TuGK-infected Col-0 plants at 12 dpi. Bar, 1 cm. The red arrowheads indicate the curing and serrated systemic leaves after TuGR infection. b Endogenous AGO1 and HC-Pro levels in mock, TuGR-, and TuGK-infected plants. The CP levels were used to confirm TuMV infection. RUBISCO (asterisk) was used as loading controls. c miRNA target gene expression of AGO1 (miR168), ARF16 (miR160), and MYB33 (miR159) in mock, TuGR-, and TuGK-infected plants. The bars represent the standard errors (n = 3). The statistical significance was assessed based on Student’s t test. ** indicates P values < 0.01
TuMV infection inhibits the RISC activityWe subsequently evaluated whether RISC activity might be affected during TuMV infection (Fig. 6a). We collected compatible amounts of AGO1-IP from mock, TuGR-, and TuGK-infected plants (Fig. 6b). The RISC assay demonstrated that the MYB33-230 substrate in TuGR-infected plants has lower cleavage efficiency compared with mock or TuGK-infected plants (Fig. 6c). As shown in Fig. 6c, 20 ng AGO1-IP from mock plants had 50.7% RISC activity, whereas 20 ng AGO1-IP from TuGR- and TuGK-infected plants had 38.5% and 43.9% RISC activity, respectively (Fig. 6a and c). These data demonstrated that severe TuMV infection inhibits RISC activity, which is consistent with the RISC results obtained in P1/HC-ProTu plants.
Fig. 6
Evaluation of the in vitro RISC cleavage efficiency using TuMV-infected plants. a MYB33-230 RNA substrates were cleaved by AGO1-IP from mock, TuGR-, and TuGK-infected Col-0 plants. The numbers indicate the amounts of AGO1-IP that were used for cleavage. b The AGO1-IP amounts from mock, TuGR-, and TuGK-infected Col-0 plants were evaluated by western blotting with α AGO1 IgG to normalize the RISC cleavage efficiency. In: input; IP: immunoprecipitation; FT: flow through. c Normalized RISC cleavage efficiency of AGO1-IP from mock, TuGR-, and TuGK-infected Col-0 plants
DiscussionGain-of-function and loss-of-function genetic studies are fundamental approaches to understanding gene function. Agrobacterium-mediated transformation is popular for creating ectopic transgenic plants or T-DNA insertion mutants. However, a few studies have identified the T-DNA insertion position. Because NGS is more efficient and economical, identifying franking sequences of T-DNA insertion becomes achievable and with several benefits, including whether the insertion is on a gene body and which chromosome is inserted. In addition, transgenic plants with a transparent genetic background can introduce another mutation by crossing or CRISPR-Cas approaches. The T-DNA of P1/HC-ProTu plants is inserted on chromosome 1 without breaking any gene body; thus, the atg8a mutant that localizes in chromosome 4 can be introduced into the transgenic plant to generate the P1/HC-ProTu/atg8age plants [11, 16]. We also identified the other T-DNA insertions in these P1/HC-Pro plants. The insertion data with the locations of the RNA silencing component provide information for crossing strategy decisions. For instance, the T-DNA of the P1/HC-ProZy plant is inserted on chromosome 4, which cannot introduce the atg8a mutant into this transgenic plant by crossing, and this deficiency might be overcome by gene editing. However, for introducing the other silencing mutants shown in Fig. 3b, the crossing approach can be applied to P1/HC-ProZy plants. Moreover, we can generate double transgenic plants by crossing two homozygous VSR transgenic plants, e.g., P1/HC-ProTu plants × P1/HC-ProZy plants, to study whether the synergistic effect would cause silencing suppression.
An in vitro RISC assay enables the direct analysis of AGO1 biological function. Baumberger et al. was the pioneering group that developed an in vitro RISC assay via IP of Flag-tagged AGO1 [17]. This system should be conducted in the ago1-36 background to eliminate the impact of endogenous AGO1. Hence, establishing the Flag-AGO1 approach might be time-consuming for genetic screening and limit the utility of in vitro RISC assay to other mutants or transgenic plants when the simultaneous introduction of two elements, including the Flag-AGO1 transgene and ago1-36 locus, is needed. Moreover, the T-DNA insertion of Flag-AGO1 is unclear and might have a genetic linkage with the other mutants. T-DNA Finder can obtain the insertion information for Flag-AGO1/ago1-36 plants. In contrast, the polyclonal α-AGO1 antibody targets the 240 aa N-terminus of AGO1 and specifically immunoprecipitates endogenous AGO1. We can rapidly take advantage of the in vitro RISC assay to verify the cleavage efficiency in any mutant and plant. AGO1 has 56 different mutant alleles, and some alleles might lose certain functions [19]. For instance, ago1-42 is mutated in the PAZ domain, which might affect the miRNA loading; whereas the ago1-27 mutant might lose miRNA duplex binding [13, 24]. The AGO1-IP from these mutants can be used to perform small RNA profiling, RISC activity, and the RISC complex assembly. In contrast, viral-infected Arabidopsis can also be applied to investigate various VSRs in RISC inhibition efficiency. Virus infection of various ago1 mutants can be used to investigate the interaction between VSRs and AGO1 through AGO1-IP with an α-AGO1 antibody.
Moreover, we demonstrated that the structure of the RNA substrate is critical for the in vitro RISC assay. The assay with the substrates ARF10-230 and ARF10-191 showed different cleavage efficiencies. We hypothesized that a targeting site length of the 3'-end might be essential. A 3'-end target site that is too long might form a complicated secondary structure, which may be difficult for AGO1 to access, as in the case of the ARF10-230 substrate. Thus, the RNA structure needs to be considered before determining the substrate sequence. We observed that miR159 and miR398 have more cleavage efficiency than miR164 and miR160. The amounts of miRNAs incorporated into AGO1 might cause a difference in the cleavage efficiency. The application of AGO1-IP with small RNA NGS can identify the AGO1-containing miRNA and siRNA profiles, which allows us to answer this question. Furthermore, AGO-IP from virus-infected plants can also obtain the viral siRNA profile, which can be applied in viral cleavage evaluation. AGO2 has been demonstrated to be involved in resistance to viral infection [27]. Therefore, we can apply the same approach to generate AGO2-specific antibodies for AGO2-IP and in vitro RISC assays in comparative studies.
Pollari et al. demonstrated that a WG pair of HC-ProPVA plays a role for AGO1 subcellular colocalization and interaction [26]. Wei et al. also demonstrated that HC-ProTu colocalized with AGO1 but had no direct interaction, as revealed by Förster resonance energy transfer (FRET) analysis [11]. Indeed, the IP results from three P1/HC-Pro plants did not show that three HC-Pros directly interact with AGO1. However, Pollari et al. used a cross-linking approach to demonstrate the interaction of HC-ProPVA coupled with AGO1 [26]. It has been proposed that HC-ProTu accelerates the aggregation of AGO1 and the host proteins to form RNA granules, which would recruit the colocalization of AGO1 and HC-Pro to cytoplasmic foci [28]. To summarize, we hypothesized that HC-Pro might interact with bridge host factors, which could also interact with AGO1, through the WG pair of HC-Pro, suggesting that HC-Pro and AGO1 might have an indirect interaction.
Because comparable AGO1-IP amounts from three P1/HC-Pro plants, the more attenuated RISC activity of the P1/HC-ProTu sample might not cause by decreased AGO1 levels. Wei et al. demonstrated that HC-ProTu specifically inhibits HEN1 activity, resulting in 50% unmethylated miRNAs that could not be loaded into AGO1 (Fig. 7a) [11]. However, HC-ProZy and HC-ProTe did not inhibit HEN1, which might allow more methylated miRNAs to be loaded into AGO1 for target RNA cleavage (Fig. 7b). We hypothesized that reduced miRNA content, e.g., 50% methylated miR159, in AGO1-IP of P1/HC-ProTu plants resulted in lower RISC cleavage efficiency (Fig. 7a). Although P1/HC-ProTu/atg8age plants still contain approximately 50% unmethylated miRNAs [11]; however, restoring AGO1 in the plants allows them to carry more methylated miRNAs to restore the RISC activity (Fig. 4a and b). Moreover, P1/HC-ProTe plants containing 100% methylated miRNAs showed less RISC activity, suggesting that multiple RISC interference mechanisms might exist in different potyviruses. In summary, the reduced AGO1 amount and less methylated miRNAs in AGO1-IP are the unique abilities of HC-ProTu to inhibit RISC activity, which means that TuMV can inhibit RISC activity more significantly compared with Col-0 and ZYMV-infected plants. In contrast, TEV infection may inhibit RISC less significantly than TuMV (Fig. 7).
Fig. 7
The working hypothesis for various HC-Pros in inhibiting the RISC activity. a The model of RISC inhibition by HC-ProTu. b The model to explain no effect on RISC inhibition by HC-ProZy and HC-ProTe. The dark red lines represent as mature miR159. The blue lines represent other small RNAs (sRNAs). The gray lines indicate the miRNA*. The dark red color indicates the methylated miRNAs. The pink color indicates the unmethylated miRNAs. Me, the methylated miRNA
Shiboleth et al. demonstrated that the FINK motif mutation of HC-ProZy still retained vigorous RNA silencing suppression, suggesting small RNA sequestering is not the only way for HC-ProZy inhibition RNA silencing but might exist multiple suppression approaches [6]. Wei et al. also demonstrated that HC-ProZy and HC-ProTe still can slightly trigger AGO1 degradation [11]. Indeed, HC-ProTu did not have a small RNA binding ability but can enhance autophagic AGO1 degradation and RISC inhibition more than HC-ProZy and HC-ProTe because of HEN1 inhibition [11]. Therefore, we proposed that conserved and diverse VSR functions exist simultaneously in different viral HC-Pros.
留言 (0)