
記住我
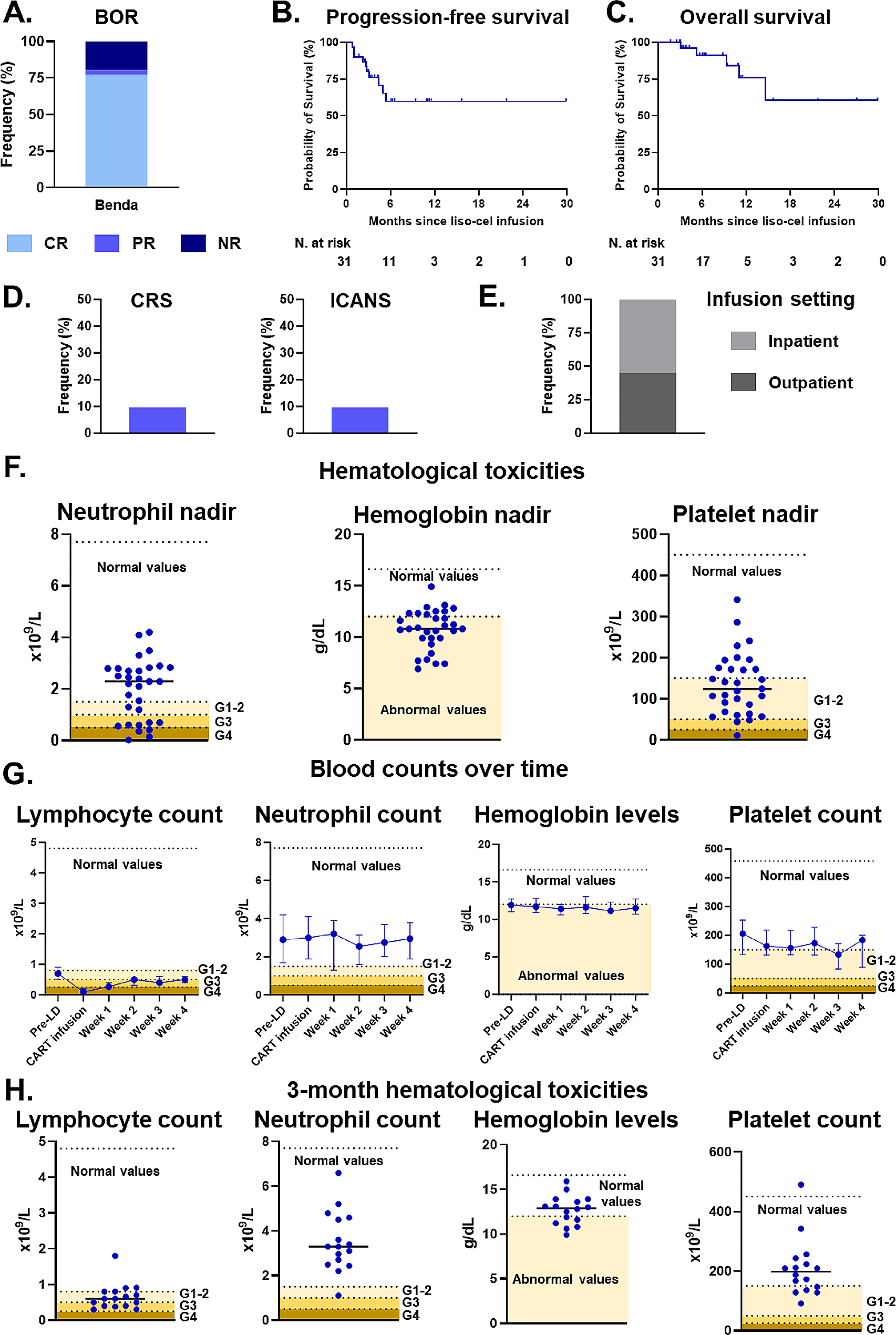



To identify pH-dependent anti-CD47 antibodies, we implemented a pH-dependent selection strategy to select antibodies from a human antibody phage display library [30]. Library phages were incubated with CD47 in pH 6.0 buffer, and subsequently eluted with pH 7.4 buffer (Fig. 1a). With this panning strategy, phages that bound to CD47 with high affinity at acidic-pH (pH < 7.4) and low affinity at physiological-pH (pH 7.4) were enriched, and two clones that bind to CD47 in a pH-dependent manner (BC2 and BC27) were obtained (Fig. 1b). The binding affinity of the two antibodies to CD47 was measured using SPR at pH 7.4 and pH 6.8: BC27 exhibited relatively higher pH-dependence than BC2, which defined as the ratio of KD at pH 7.4 to pH 6.8, and was therefore selected for further evaluation (Fig. 1c, d and Additional file 1: Fig. S1a). A flow cytometry-based blocking assay showed that BC27 was not able to completely antagonize the CD47-SIRPα interaction at a concentration as high as 1 µM even at acidic-pH condition (Additional file 1: Fig. S1b); this low blocking efficiency might reflect its low binding affinity at acidic-pH.
Fig. 1
Generation and characterization of pH-dependent anti-CD47 antibodies. a Schematic diagram depicting the pH-dependent selection strategy. Library phages were binding to CD47-ECD in pH 6.0 buffer, and subsequently eluted by pH 7.4 buffer; enriched phages were rescued for the next round of panning or, single clones were selected for characterization. b Characterization of the pH-dependent binding of BC2 and BC27. Serial dilutions of antibodies (in phage-scFv form) binding to CD47-ECD at pH 7.4 and pH 6.0, measured by ELISA. c Representative SPR sensograms of pH-dependent antibodies binding to CD47-ECD at pH 7.4 and pH 6.8; CD47-ECD was diluted from the indicated starting concentrations. Both original sensorgrams (colored lines) and fitting curves (black lines) are shown. d Summary of SPR measurements of pH-dependent antibodies, and a non-pH-dependent control Hu5F9; the affinity measurement results (KD) shown represent the mean and standard deviation of three replicated experiments. pH-dependence is defined as the ratio of KD at pH 7.4 to pH 6.8. The position of the mutated residues and their corresponding sequences within the HCDR3 and LCDR3 of BC27 (and its variants) are shown. e Top, serially diluted antibodies binding to CD47-ECD from pH 6.2 to pH 7.6, measured by ELISA. Bottom, the calculated EC50 for the data of the top panel
We next sought to improve both the binding affinity and pH-dependent properties of BC27. Random mutations were introduced into both the CDR3 of the heavy chain (HCDR3) and the light chain (LCDR3) to construct phage display sub-libraries. These sub-libraries were subsequently selected using the pH-dependent strategy described above. After several rounds of intensive optimization-selection, about 15 variants with improved affinity and/or pH-dependence were obtained. Among these variants, BC31M4 (KD = 10.7 nM) had about 12-fold higher affinity than the parental antibody BC27 (KD = 127.6 nM) at pH 6.8, thus achieved a significant increase of pH-dependence from 9.2-fold to 42.7-fold. We also obtained a weak pH-dependent variant BC31M5 that had a pH-dependence of about 1.9-fold (similar affinity to BC31M4 at pH 6.8, much higher affinity at pH 7.4) (Fig. 1c, d). Remarkably, only one amino acid is different between these two antibodies: residue 108 in the HCDR3 is threonine (T) in BC31M5 but alanine (A) in BC31M4.
To further characterize the pH-dependent binding property of BC31M4 and BC31M5, we measured their bindings to CD47 at serial pH conditions (from 6.2 to 7.6) by ELISA. BC31M4 exhibited an apparent pH-dependent binding pattern with 72.5-fold binding increase from pH 6.8 to 7.6 (an EC50 of 0.03 nM at pH 6.8 and 2.12 nM at pH 7.6) (Fig. 1e); when the pH was lower than 6.8, the affinity did not increase further. Thus, BC31M4 binds most efficiently with CD47 at pH 6.8 or below, matching with the acidic conditions of most solid tumor microenvironments. In contrast, BC31M5 exhibited slight pH-dependent binding ability. The aforementioned anti-CD47 antibody Hu5F9, which has high binding affinity but no pH-dependence (Fig. 1d and Additional file 1: Fig. S1a), did not exhibit pH-dependent binding ability in these assays. Given their different pH-dependent binding properties, we chose BC31M4 as the therapeutic antibody candidate and BC31M5 as a weak-pH-dependent antibody control for further investigation.
Histidine residues contribute to the pH-dependent binding of BC31M4To understand the structural basis of this pH-dependent binding property, we solved the crystal structure of BC31M5 (Fab form) in complex with the CD47 extracellular domain at pH 6.5 (2.8 Å resolution; Fig. 2a and Additional file 1: Table S1). Recalling that BC31M4 and BC31M5 have only one amino acid difference, they likely bind to similar epitopes in CD47. The complex structure reveals 15 epitope residues in CD47 at the interaction interface (Table 1). Most of these residues overlap with the previously reported SIRPα binding sites [35], suggesting that these antibodies will maximally antagonize the CD47-SIRPα interaction. In addition, residue T108 in the HCDR3 of BC31M5 forms polar contacts with Q1 and T102 of CD47 (Fig. 2a, b); however, it is expected no similar polar contacts between BC31M4 and CD47 as this residue is A108 in BC31M4.
Fig. 2
Structural characterization of the pH-dependent binding of BC31M4. a Crystal structure of the BC31M5-CD47 complex, depicted as ribbons. T108 (orange) in VH and histidines (red) in VL are shown as sticks. b Detailed view of the BC31M5-CD47 interface. Gray dashed lines indicate electrostatic interactions between BC31M5 and CD47. Side chains of contacted residues are shown as sticks. Yellow dashed lines indicate π-contacts. c The three histidines (H38, H55, or H107) in the VL of BC31M4 (in phage-scFv form) were mutated into any other amino acids individually (site saturation mutagenesis). These mutants binding to CD47 at pH7.4 and pH 6.5 were measured by ELISA. d Serial dilutions of BC31M4 mutants (in hIgG1 form) with the indicated double or triple histidine-to-arginine substitutions binding to CD47-ECD at pH7.4 and pH 6.8, measured by ELISA
Table 1 Contact residues between BC31M5 and CD47We next explored how particular amino acids contribute to the pH-dependent binding of BC31M4. The side chain of histidine has an acidic ionization constant (pKa) value of around 6.5 in most proteins [36], this property supports differential antibody binding around this pH range. There are three histidine residues (H38, H55, and H107, defined by IMGT numbering) located within the CDRs of the variable region of light chain (VL). The complex structure shows that H38 (LCDR1) and H107 (LCDR3) form electrostatic contacts with E97 and E104 in CD47, respectively (Fig. 2b and Table 1). These data suggest that H38 and H107 support the pH-dependent binding of BC31M4.
We subsequently conducted site saturation mutagenesis for these histidines individually. The mutagenesis was performed with phage-scFv form, and binding to CD47-ECD was evaluated by ELISA. Many of the mutations dramatically decreased the binding affinity of BC31M4 at pH 6.5. At positions H38 and H107, mutations by replacing with other positively charged amino acids—arginine (R) or lysine (K)—which maintained binding activities comparable to wild-type BC31M4 or retained weak binding activities—confirming that electrostatic contacts formed through protonation of amino acids at these positions promotes BC31M4 binding to CD47. Additionally, at position H55 (LCDR2), a histidine-to-arginine (H-to-R) mutation retained a relatively weak binding activity. And at H55 and H107, mutation to aromatic amino acids—phenylalanine (F), tyrosine (Y), or tryptophan (W)—retained weak binding activity (Fig. 2c). Note that the complex structure shows that H55 and H107 form intra-chain π–contacts with the antibody’s W56 and W116 residues, respectively (Fig. 2b); similar π–contacts may form with mutant variants bearing aromatic (π–π) or positively charged (cation–π) amino acids.
To characterize the contribution of individual histidine to the pH-dependent binding of BC31M4, we generated a set of BC31M4 mutants in which two of the three histidines were substituted with positively charged arginine. These substitutions were performed in hIgG1 form, and binding to CD47 was evaluated by ELISA, with pH-dependent binding assessed as binding deviation between pH 7.4 and pH 6.8. The 38R/55R and 55R/107R substitutions retained pH-dependent binding to CD47 at a similar level to wild-type BC31M4; in contrast, the 38R/107R substitution dramatically diminished the pH-dependent binding; and the 38R/55R/107R substitution completely abrogated pH-dependent binding while also greatly reducing overall binding (Fig. 2d). Collectively, these results support that H38 and H107 are directly involved in the pH-dependent binding between BC31M4 and CD47; whereas H55 contributes minimally to the pH-dependent binding, apparently acting indirectly.
BC31M4 blocks the cell-surface CD47-SIRPα interaction and promotes macrophage phagocytosis of tumor cells in a pH-dependent mannerTo determine whether BC31M4 binds to the cell-surface CD47 and blocks the CD47-SIRPα interaction in a pH-dependent manner, a flow cytometry-based binding and blocking assay was performed to evaluate the binding and blocking activity of antibodies to different cells, including a CHO cell line stably expressing full-length human CD47 (CHO-hCD47), Raji (B lymphoma), Jurkat (T lymphoma) and MDA-231 (breast cancer) cells, at pH 6.8 and pH 7.4 conditions separately. Compared to BC31M5, BC31M4 bound to these cells and blocked the binding of SIRPα to these cells with much higher efficiency at pH 6.8 than at pH 7.4 (Fig. 3a). We further investigated the binding of antibodies (BC31M4, BC31M5, and Hu5F9) to human T cells at pH7.4 and pH 6.8. Compared to Hu5F9, both BC31M4 and BC31M5 exhibited pH-dependent binding to CD4+ and CD8+ T cells in human PBMCs, evidenced by the apparent higher binding efficiency at pH 6.8 than at pH 7.4 (Additional file 1: Fig. S2a).
Fig. 3
BC31M4 blocks the cell-surface CD47-SIRPα interaction and promotes macrophages phagocytosis of tumor cells. a Top, the binding and blocking activity of antibodies to different tumor cells at pH 7.4 and pH 6.8. Bottom, the calculated EC50 and IC50 for the data of the top panel. “~” indicates estimated values; “-” indicates EC50 or IC50 values are not measurable. b, c Macrophages phagocytosis of Raji cells induced by different antibodies at pH 7.4 and pH 6.8. All antibodies used are of hIgG1 isotype except BC31M4-F(ab′)2. Antibody concentration is 1 μg/ml for Rituximab, and 5 μg/ml for BC31M4 and BC31M4-F(ab′)2. Phagocytosis index is determined as the number of phagocytosed Raji cells per 100 macrophages. d Schematic diagram of tumor inoculation and antibody treatment in human tumor xenograft models. e Tumor growth of mice treated as in (d). f Survival of mice in (e). n, number of mice
We next sought to determine the ability of BC31M4 to promote the phagocytosis of tumor cells, an antibody-dependent cellular phagocytosis (ADCP) analysis was performed using Raji cells as targets and bone marrow-derived macrophages (BMDMs) from the C57-hCD47/hSIRPα transgenic mice as effectors. In the C57-hCD47/hSIRPα mice, the IgV domains of both CD47 and SIRPα (responsible for the CD47-SIRPα interaction) are replaced with the corresponding human sequences. Raji cells opsonized with different concentrations of antibodies were incubated with BMDMs at pH 6.8 and pH 7.4 conditions separately; antibodies used here were converted into hIgG1 isotype that mediates strong Fc effector functions in both human and mouse [37,38,39]. BC31M4 did not induce macrophage phagocytosis of Raji cells at pH 7.4, but significantly promoted phagocytosis at pH 6.8 in a dose-dependent manner. BC31M5 promoted macrophage phagocytosis of Raji cells to a similar extent at pH 6.8 and pH 7.4 (Fig. 3b).
Subsequently, we evaluated the ability of BC31M4-F(ab′)2 fragment (lacks Fc) alone or in combination with a tumor-specific monoclonal antibody in inducing phagocytosis. Using Rituximab-hIgG1 (binds to CD20 on Raji cells) as the tumor-specific antibody, Raji cells as target cells, we observed that BC31M4-F(ab′)2 in combination with Rituximab induced a significantly higher level of phagocytosis than Rituximab alone did at pH 6.8; however, BC31M4-F(ab′)2 alone failed to induce phagocytosis of Raji cells (Fig. 3c).
Additionally, using E.G7-hCD47 cells (a mouse-derived T lymphoma cell line that stably expresses human CD47) as the target cells, BC31M4-F(ab′)2 alone or BC31M4-mIgG1 (a mouse isotype with weak Fc effector functions [38, 39]) also did not induce the phagocytosis of E.G7-hCD47, whereas BC31M4-mIgG2a (a mouse isotype with strong Fc effector functions [38, 39]) induced phagocytosis of E.G7-hCD47 cells (Additional file 1: Fig. S2b). Collectively, these results suggest that BC31M4 promotes macrophages phagocytosis of tumor cells requires both blockade of the CD47-SIRPα inhibitory pathway and activation of the Fc-mediated effector functions.
BC31M4 inhibits tumor growth in human tumor xenograft modelsGiven that BC31M4 can promote macrophage phagocytosis against tumor cells in vitro, we next examined whether BC31M4 has antitumor effects in vivo using human tumor xenograft models. Raji lymphoma cells were subcutaneously (s.c.) inoculated into NOD-SCID mice, and treated with antibodies (in hIgG1 form) intraperitoneally (i.p.) (Fig. 3d). Compared to the PBS control, both BC31M4 and BC31M5 significantly inhibited tumor growth and significantly prolonged the survival of mice (Fig. 3e, f). In addition, the antitumor efficacy of BC31M5 was more potent than BC31M4 in these xenograft models, which may be attributed to the higher binding affinity of BC31M5 with CD47, about threefold higher than BC31M4 at pH 6.8 (Fig. 1c, d), albeit with only 1.9-fold pH-dependency. Note that human CD47 expression is absent in these immunocompromised mice, any human CD47-specific antibodies will only bind to tumor cells in these models, regardless of whether they have pH-dependent binding ability or not; and antibodies with higher binding affinity (BC31M5), but not better pH-dependence (BC31M4), are more likely to have better antitumor activity; however, human CD47 is ubiquitously expressed on healthy cells in patients. Therefore, the advantage of BC31M4’s pH-dependent binding cannot be appropriately assessed in these xenograft tumor models.
BC31M4 selectively accumulates in tumors and exhibits superior PK properties in C57-hCD47/hSIRPα miceGiven that BC31M4 and BC31M5 do not recognize mouse-CD47 (mCD47), xenograft models are unsuitable for evaluating their therapeutic effects in a physiological context wherein CD47 is widely expressed. Thus, and seeking to evaluate the therapeutic efficacy and safety of antibodies in immunocompetent syngeneic mouse models that are more reflective of the CD47 expression profile in humans, we established syngeneic tumor models using the aforementioned C57-hCD47/hSIRPα mice. Because mCD47 does not cross-react with human-SIRPα (hSIRPα) [40], the mouse-derived tumor cell lines were humanized via stable transfection with full-length hCD47 (Cell-hCD47) (Additional file 1: Fig. S3a). Furthermore, to minimize the risk of eliciting anti-drug antibodies in the immune competent C57-hCD47/hSIRPα mouse, antibodies used were converted (unless otherwise noted) into the mIgG2a isotype.
To verify whether BC31M4 selectively binds to tumors in vivo, we measured the biodistribution of antibodies in vivo using whole-body near-infrared fluorescence (NIRF) imaging. Antibodies were labeled with fluorophores (Additional file 1: Fig. S3b). E.G7-hCD47 lymphoma cells were s.c. inoculated into C57-hCD47/hSIRPα mice; when tumors reached volumes about 500 mm3 (Additional file 1: Fig. S3c), these mice were treated i.p. with antibodies. Previous studies showed that adopting a low priming dose before a high maintenance dose (prime-plus-maintenance) can alleviate the side effects of anti-CD47 antibodies in vivo [8]. To reduce the risk of causing severe side effects, antibodies were administered with a priming dose of 1 mg/kg, followed by a single maintenance dose of 5 mg/kg two days later (Fig. 4a). After the priming dose, BC31M4 exhibited more obvious intratumoral accumulation than Hu5F9 and BC31M5 (Additional file 1: Fig. S3d). After the maintenance dose, BC31M4 obviously accumulated at tumor sites at both 24 h and 72 h; in contrast, no similar intratumoral accumulation was observed for BC31M5 and Hu5F9 (Fig. 4b).
Fig. 4
BC31M4 selectively accumulates in tumors and exhibits superior PK properties in C57-hCD47/hSIRPα mice. a Schematic diagram showing the tumor inoculation and antibody treatment of mice in (b, c). C57-hCD47/hSIRPα mice were s.c. inoculated with E.G7-hCD47 cells on the right lower flank. Mice were i.p. treated with Cy7-labeled antibodies or PBS. b Antibody distribution in mice, monitored by in vivo fluorescence imaging at the indicated time points after the maintenance dose (5 mg/kg). Hair on the back of mice was removed before imaging. Green circles indicate the location of tumors. c Antibody distribution in tumors and different organs of mice (n = 5 per group); tumors and organs were isolated for ex vivo fluorescence imaging at 3 h and 72 h after the maintenance dose. Representative images from one mouse in each group are shown. d Summary of the antibody distribution in tumors and organs of (c), the average quantified radiant efficiency is shown. e Summary of the tumor-to-organ fluorescence intensity ratio of (c). f PK analysis of antibodies in C57-hCD47/hSIRPα mice (n = 2–4 per group). Mice were treated i.p. with a priming dose of 1 mg/kg two days before delivery of a single dose of 20 mg/kg. The antibody concentration in serum was monitored, and the half-life of each antibody is indicated
We next profiled the antibody distribution in different organs and tissues. Additional mice given the same treatment as above were euthanized at 3 h or 72 h post maintenance dose, tumors and organs (spleen, liver, kidney, and lung) were isolated for fluorescence imaging and quantification of the fluorescence intensity (Fig. 4c, d and Additional file 1: Fig. S2e). Comparing the fluorescence intensities in tumors showed that the BC31M4 signal significantly increased between the 3 h and 72 h sampling time points, whereas the BC31M5 signal remained at a similar level and the Hu5F9 signal appeared to decrease slightly between these time points. The organ analysis showed that the fluorescence intensity of all antibodies was markedly decreased between the 3 h and 72 h sampling time points for all examined normal organs. At 3 h, all antibodies exhibited similar fluorescence intensity in tumors; whereas compared to BC31M5 and Hu5F9, the BC31M4 signal was slightly lower (not significant) in spleen, and significantly lower in other normal organs. At 72 h, the BC31M4 signal was significantly higher than BC31M5 and Hu5F9 in tumors; whereas in normal organs, comparing to BC31M5, the BC31M4 signal was significantly higher in liver, lung, and kidney, but not in spleen; comparing to Hu5F9, BC31M4 signal was significantly higher in spleen and liver, but not in lung and kidney (Fig. 4c, d).
To better determine the extent of antibody accumulation in tumors relative to that in normal tissues, we subsequently calculated the fluorescence signal intensity ratio of the tumor to each of the other organs (tumor-to-organ ratio). Compared to Hu5F9, BC31M4 exhibited relatively high intratumoral accumulation (tumor-to-organ ratio > 1) at 3 h based on all the evaluated organs, and this accumulation became more substantial (tumor-to-organ ratio > 9) at 72 h (Fig. 4e). Additionally, although the pH-dependent binding property of BC31M5 is poor, it also exhibited some level of intratumoral accumulation but significantly lower than BC31M4 especially in tumor-to-lung ratio at 3 h and tumor-to-spleen ratio at 72 h. These results support that pH-dependent binding endows antibodies with selective binding capacity for tumor cells while sparing the healthy cells.
Poor PK due to antigen sink effect is a concern for anti-CD47 antibodies [8]. The weak binding of BC31M4 to normal tissues should reduce antigen sink effect and improve its PK properties. We therefore performed a single-dose PK analysis of antibodies using C57-hCD47/hSIRPα mice. BC31M4 exhibited much better PK properties with a half-life of 113 h, compared with 19.3 h for BC31M5 and 9.7 h for Hu5F9 (Fig. 4f). Collectively, these results support that BC31M4’s pH-dependent binding property endows it with superior PK properties in immunocompetent syngeneic mouse models. It is worth noting that the PK profiles of the three antibodies suggest that the higher accumulation of BC31M4 in most normal organs at 72 h (Fig. 4c, d) is likely due to its higher serum concentration, but not its binding to healthy cells.
BC31M4 causes minimal side effects in C57-hCD47/hSIRPα miceVarious side effects have been reported for anti-CD47 therapies in pre-clinical and clinical studies [8, 16,17,18]. We next adopted different treatment strategies to compare the safety of our antibodies in vivo (Fig. 5a). Briefly, a body temperature drop was the most commonly observed symptom in treated C57-hCD47/hSIRPα mice, which often occurred around 3 h after administration of the first maintenance doses (Fig. 5a); additionally, body temperature drop was less observed after administration of the rest maintenance doses (data not shown). Moreover, the mice that displayed severe temperature drop (≥ 1 °C) were also lethargic and were at obviously increased risk of treatment-related-death (Fig. 5b). Mouse death mostly occurred involving the administration of BC31M5 or Hu5F9 in doses higher than 10 mg/kg (Fig. 5b).
Fig. 5
BC31M4 causes minimal hematotoxicity in C57-hCD47/hSIRPα mice. a Top, schematic diagram of mouse treatment and body temperature measurement. ‘0’ indicates no priming dosing. Bottom, body temperature changes compared to the average temperature in the corresponding control group; each histogram represents one mouse. b Summary of the treatment-related-death of the antibody treated mice. c Hemagglutination test of antibodies in vitro. RBC aggregation was analyzed by flow cytometry. Dot plots of the FSC-A and SSC-A values are shown; percentage of events in the gate (red frame) is denoted. d, e, g–i Summary of the complete blood count test of antibody-treated mice. The parameters of RBC count (d), red cell distribution width (e), platelet count (g), mean platelet volume (h), and the proportion of individual leukocytes in WBCs (i) are shown. f Spleen weight of mice used in the complete blood count test. Antibodies used in (a–f) are mIgG2a isotype unless otherwise noted
When mice were directly treated with a high dose (20 mg/kg) of antibody i.p. without priming, both BC31M5 and Hu5F9 caused severe body temperature drop in more than half of the mice, and this was accompanied with very high treatment-related-death rates (≥ 50%); however, similar side effects occurred in only 6% (1/16) of the BC31M4-treated mice. After adopting the prime-plus-maintenance treatment strategy comprising a priming dose of 1 mg/kg and maintenances doses of 10 mg/kg, no side effects were observed in the BC31M4 treated mice. In contrast, the proportions of mice displaying severe body temperature drop and/or treatment-related-death rates were still very high upon either BC31M5 (38%) or Hu5F9 (29%) treatment under the maintenance dose of 10 mg/kg. When the maintenance dose was further decreased to 1 mg/kg, no treatment-related-death occurred for Hu5F9-treated mice, but 1 out of 10 animals displayed a severe temperature drop. Moreover, when antibodies were converted into the mIgG1 isotype (weak Fc effector functions), neither BC31M4-mIgG1 nor Hu5F9-mIgG1 caused any side effects in mice treated with maintenance doses of 10 mg/kg (Fig. 5a, b). These results suggest that the tolerability of anti-CD47 antibody is Fc-effector function dependent.
As a consequence of the relatively high expression of CD47 on RBCs and platelets, hematotoxicity—which includes anemia, thrombocytopenia, hemagglutination, and neutropenia—has been a major concern among the various side effects observed in clinical studies of anti-CD47 therapies [16,17,18]. We next sought to determine the basis of the severe side effects observed in mice during antibody treatment and further evaluate the safety of BC31M4. We first performed an in vitro hemagglutination assay using RBCs from the C57-hCD47/hSIRPα mice. The RBC aggregation was further assessed by flow cytometry. Compared to Hu5F9, BC31M4 caused obviously less RBC aggregation, represented by the lower percentage of cell populations with high FSC-A and SSC-A values (Fig. 5c).
We next evaluated the in vivo hematotoxicity of BC31M4. Given that both BC31M5 and Hu5F9 caused severe side effects (to a similar extent) (Fig. 5a, b), we selected Hu5F9 for comparison in the following assay. Healthy mice were treated with a single dose of 20 mg/kg antibodies (mIgG2a isotype) or PBS i.p., and hematologic parameters were assessed at 3 h after treatment. Compared to the PBS control, we detected significant decreases in RBC count, hemoglobin (Hgb), and hematocrit (HCT) in mice treated with Hu5F9 (Fig. 5d and Additional file 1: Fig. S4a, b), features indicating acute anemia; and the RBC distribution width (RDW) was significantly increased (Fig. 5e), indicating increased variation in RBC volume that could result from RBC aggregation. This was consistent with the above finding that Hu5F9 caused more severe RBC aggregation than BC31M4 in vitro. In contrast, mice treated with BC31M4 did not exhibit any alteration in these indices. Besides, the significant increase of the spleen weight of mice treated with Hu5F9 (Fig. 5f) is consistent with the commonly observed splenomegaly in hemolytic anemia due to accumulation of macrophages and CD47−/− RBCs in mice [1].
Significant drops in platelet counts were observed in mice treated with BC31M4 and Hu5F9 (Fig. 5g). However, a significant increase in mean platelet volume (MPV, indicating platelet aggregation) was observed for Hu5F9-treated but not for BC31M4-treated mice (Fig. 5h). Moreover, compared to the proportion of individual leukocytes in PBS controls, BC31M4 treatment did not change the proportion of individual leukocytes assessed; whereas Hu5F9 treatment caused a significant decrease in neutrophils (~ 22%) that was accompanied by relative increases in lymphocytes and basophils, which indicated neutropenia of Hu5F9-treated mice (Fig. 5i). Collectively, these results showed that BC31M4 causes minimal hematotoxicity during treatment.
BC31M4 efficiently promotes adaptive immune responses against tumors in syngeneic mouse modelsWe next evaluated the therapeutic efficacy of antibodies in syngeneic mouse models. The anti-CD47 antibodies (mIgG2a isotype) alone did not exert any antitumor effect in the syngeneic mouse models (Additional file 1: Fig. S5a–e). Previous studies showed that the activation of the adaptive immune system was required for the antitumor effect of anti-CD47 therapy in immunocompetent syngeneic mouse models [21, 22]. Thus, we explored whether BC31M4 promotes antitumor responses against syngeneic tumors in mice with a stimulated adaptive immune system (combination therapy). The E.G7 cell line is derived from EL4 lymphoma cells and expresses ovalbumin (OVA); the OT-I T cells are OVA-specific CD8+ T cells and can be isolated from OT-I transgenic mice [41]. To determine whether Fc effector function is required in this combination therapy, BC31M4 or Hu5F9 was also converted into mIgG1 isotype and included for evaluation in these experiments.
We subsequently established the E.G7-hCD47 syngeneic models as described above, and treated these mice i.p. with a priming 1 mg/kg antibody dose (or PBS) on day 3, followed by two maintenance doses of 10 mg/kg on day 5 and 11; OT-I T cells were intravenously (i.v.) transfused into all mice on day 8 (Fig. 6a). Compared to the OT-I T cell monotherapy (i.e., PBS priming and maintenance), the BC31M4 and OT-I T cell combination therapy significantly delayed tumor growth and significantly prolonged the survival of mice. Although BC31M5 also significantly inhibited tumor growth compared to the OT-I T cell monotherapy, it is not as potent as BC31M4, which reflects its decreased tumor selectivity and faster PK than BC31M4 in vivo. Furthermore, combination therapies using the Hu5F9 or BC31M4-mIgG1 did not affect tumor growth and survival (Fig. 6b, c). We also observed in a similar combination therapy experiment that Hu5F9-mIgG1 did not confer any antitumor effects (Additional file 1: Fig. S6). These results suggest that BC31M4 is more potent than the other antibodies in promoting adaptive antitumor immune responses in syngeneic mouse models, and the antitumor activity of BC31M4 depends on the Fc effector function.
Fig. 6
BC31M4 efficiently promotes the adaptive immune responses against tumors in syngeneic mouse models and promotes the development of immune memory. a Schematic diagram of tumor inoculation and treatment. C57-hCD47/hSIRPα mice were s.c. inoculated with E.G7-hCD47 cells on the right lower flank. Antibodies or PBS were i.p. injected, OT-I T cells were i.v. transfused. b The tumor growth of mice treated as in (a); Treatment-related-death mice were excluded from summary statistics. c Survival of mice in (b). d Schematic diagram of tumor inoculation and treatment. e The tumor growth of mice treated as in (d). f Survival of mice in (e). g Schematic diagram of tumor rechallenge. C57-hCD47/hSIRPα mice were initially inoculated and treated as in (a), cured mice were rechallenged with E.G7-hCD47 or EL4-hCD47 cells about 140 days after the initial tumor inoculation. The rechallenged tumor cells were s.c. inoculated on the left lower flank. h, i Tumor growth in rechallenged mice is shown individually. Age-matched, naïve, C57-hCD47/hSIRPα mice were included as controls. Proportions of tumor free mice are indicated. n, number of mice. Antibodies used in (a–g) are mIgG2a isotype unless otherwise noted
Recalling that BC31M4 exhibited intratumoral accumulation after the 1 mg/kg priming dose (Additional file 1: Fig. S3d), it is possible that, deployed as combination therapy, low dosages of BC31M4 may confer similarly potent antitumor activity as high dosages. To test this proposition, the E.G7-hCD47 lymphoma model was treated with a combination of BC31M4 and OT-I T cells in a similar manner as described above. Mice were treated i.p. with a priming 1 mg/kg BC31M4 (or PBS) on day 4, followed by 4 maintenance doses of 1 or 10 mg/kg on days 6, 10, 14, and 18. OT-I T cells were transfused into all mice on day 8 (Fig. 6d). Strikingly, compared to the OT-I T cell monotherapy, the antitumor efficacy under the maintenance doses of 1 mg/kg was as potent as the maintenance doses of 10 mg/kg, both of which significantly inhibited tumor growth (Fig. 6e, f). These results suggest that the tumor selectivity of BC31M4 confers it with high efficiency in promoting the antitumor immune responses in syngeneic mouse models.
BC31M4 promotes the activation of native T cells and the development of immune memoryWe next sought to determine whether the surviving mice after the combination therapy developed immune memory. Mice that survived a similar experiment—in which E.G7-hCD47 lymphoma model animals were treated with the combination of anti-CD47 antibody and OT-I T cells—were rechallenged with the E.G7-hCD47 cells or the parental EL4-hCD47 cells that did not express OVA, about 4 months after the initial tumor challenge (Fig. 6g). Previously untreated age-matched C57-hCD47/hSIRPα mice were challenged with the same tumor cells as controls.
All survived mice were resistant to the either E.G7-hCD47 rechallenge or EL4-hCD47 rechallenge. Although tumor growth was observed in one (1/5) mouse rechallenged with E.G7-hCD47 and two (2/6) mice rechallenged with EL4-hCD47 cells, the growth was much slower than in control mice, and most of the survived mice completely rejected the rechallenge cells. In contrast, in the control mice, rapid tumor growth was observed in six (6/7) mice challenged with E.G7-hCD47 and six (6/7) mice challenged with EL4-hCD47, (Fig. 6h, i). These results suggest that most of the survived mice have immune memory. Moreover, these results indicate that most of the memory cells were specific to the antigen(s) common in E.G7-hCD47 and EL4-hCD47 cells, rather than the dominant antigen OVA in E.G7-hCD47 cells, a finding implicating the activation of native T cells in the observed immune responses. Collectively, these results suggest that BC31M4 promotes the activation of native T cells against tumors and the development of immune memory in syngeneic mouse models.
留言 (0)