
記住我
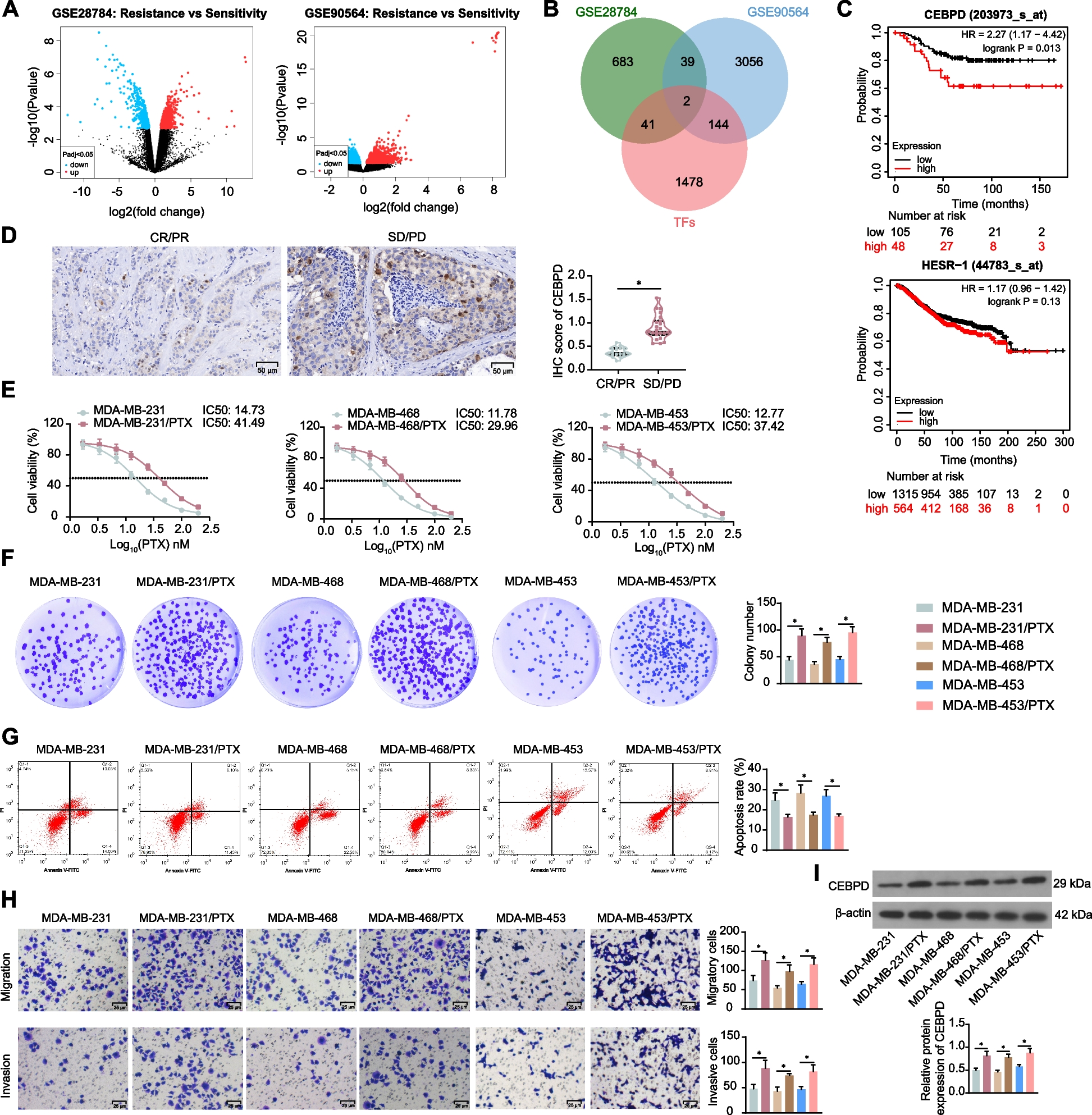




To investigate the effect of the NO donor S-nitroso-N-acetyl-DL-penicillamine (SNAP) on tumorigenesis in a mouse model, the murine Lewis lung carcinoma (LL2) tumor cell line was subcutaneously implanted in the right flank of C57BL/6 mice. Tumor-bearing mice were treated with a nitric oxide donor at various doses, and the treatment schedule is illustrated in Fig. 1A. High-dose SNAP (0.02 mg/kg) did not affect tumor progression in LL2 tumor-bearing mice (Fig. 1B). However, a low dose of SNAP (0.004 mg/kg) induced an antitumor response in LL2 tumor-bearing mice (Fig. 1C). The antitumor effect of low-dose SNAP was dose dependent. A 10-fold lower dose of SNAP (0.0004 mg/kg) did not affect tumor growth in LL2 tumor-bearing mice (Fig. 1D). In addition, low-dose SNAP treatment (0.004 mg/kg) also induced an antitumor response in the B16F1 melanoma orthotropic mouse model (Fig. 1E). The FDA-approved nitric oxide drug sodium nitroprusside (SNP) reduced tumor growth in the 0.1 mg/kg treatment group (Fig. 1F). Similarly, another FDA-approved nitric oxide donor drug, isosorbide-5-mononitrate (ISMN), inhibited tumor growth at the same low dose (0.004 mg/kg) in an LL2 tumor-bearing animal model (Fig. 1G). The results of three different nitric oxide donors indicated that a low dose of exogenous nitric oxide inhibited tumor growth. We then determined whether a low dose of nitric oxide donor increased NO levels or nitrosylated proteins in vivo and in vitro. The concentration of NO in mouse serum was increased 5 min after 0.1 mg/kg SNP treatment (p value = 0.09 in Supplementary Fig. 1 A). Due to an increase in NO levels after treatment with the low-dose NO-donating drug SNP in vivo, we further tested whether low-dose SNP affected protein nitrosylation in vitro. Therefore, THP-1 monocytic cells were treated with increasing doses of SNP, and protein nitrosylation was determined with an S-nitrosylated protein detection kit based on the biotin-switch method [28]. SNP enhanced levels of total nitrosylated protein in THP-1 cells even at the very low concentration of 1 μM (Supplementary Fig. 1 B). Additionally, low-dose SNP also enhanced the nitrosylation of heat-chock protein 90 (Hsp90), which is a known target of S-nitrosylation by NO [29] (Supplementary Fig. 1 C).
Fig. 1
Low doses of nitric oxide donors induce an antitumor response that is immune dependent. (A) Schematic of the tumor-bearing mouse model and the timeline of treatment with the nitric oxide donor S-nitroso-N-acetyl-DL-penicillamine (SNAP). LL2 tumor-bearing C57BL/6 mice received (B) 0.02, (C) 0.004, and (D) 0.0004 mg/kg SNAP. (E) B16F1 tumor-bearing C57BL/6 mice were administered 0.004 mg/kg SNAP by intraperitoneal injection. (F) LL2 tumor-bearing mice were administered SNP (0.1 mg/kg). (G) LL2 tumor-bearing C57BL/6 mice were administered 0.004 mg/kg isosorbide mononitrate (ISMN). (H) LL2 tumor-bearing immunodeficient NOD-SCID mice were administered 0.004 mg/kg SNAP or (I) 0.004 mg/kg ISMN by intraperitoneal injection. In all mice, 2 × 105 Lewis lung carcinoma LL2 cells or B16F1 melanoma cells were implanted in the right flank. Tumor volumes were measured every 2-4 days beginning on the twelfth day after tumor implantation. The tumor results are presented as the means ± SEMs. *p < 0.05, **p < 0.01, ***p < 0.001. ns, no significant difference. All p-values were obtained by two-way ANOVA
We next examined whether immune modulation was essential in mediating the antitumor response induced by low-dose SNAP and ISMN. LL2 tumor cells were implanted in the right flank of immune-deficient mice (NOD-SCID), and the therapeutic effects of low-dose SNAP and ISMN (0.004 mg/kg) were monitored. Low-dose SNAP and ISMN (0.004 mg/kg) did not affect tumor growth (Fig. 1H and I). Therefore, the immune system was essential for the low-dose SNAP-mediated antitumor response.
Low-dose SNAP regulated splenic cytokines and the Th2 cell populationSystemic immune activation is important for the response to immunotherapy [30], and we determined whether cytokines were regulated by low-dose SNAP treatment. Cytokine expression levels were analyzed using a cytokine array. We isolated tumor tissues and splenocytes on Day 20 after tumor implantation. Tumor weight and tumor size were reduced (Supplementary Fig. 2 A) in the low-dose SNAP (0.004 mg/kg) treatment group. To stimulate cytokine secretion, we cocultured LL2 cell lysates with splenocytes for 24 hours, and the supernatants were collected for cytokine detection using a cytokine array. The cytokines IL-6 and IL-10 are important for Th2-mediated function in cancer progression and exhibited reduced levels in the low-dose SNAP treatment group. Additionally, the levels of the cytokines IFN-γ and TNF-α were higher in the low-dose SNAP treatment group compared with in the control group (Supplementary Fig. 2 B-D). To further determine whether Th2 cells were decreased by SNAP, splenocytes were collected from LL2 tumor-bearing mice at Day 20 after SNAP treatment. The proportion of Th2 cells, which may be associated with B cells, decreased after low-dose SNAP (0.004 mg/kg) treatment (Supplementary Fig. 2 E and F). Our results suggested that low-dose SNAP (0.004 mg/kg) influenced the Th2 T-cell population, which is associated with alterations in cytokines in the spleen.
Single-cell RNA sequencing determines the landscape of the TME in response to low-dose SNAP treatmentTo further understand the complex changes in immune cells in the tumor microenvironment, we used single-cell RNA sequencing based on a microwell-based system (BD Rhapsody) to determine this profile. The changes in T cells and macrophages can be best detected by single-cell RNA sequencing given that these cell types are the most abundant immune cells in the tumor microenvironment. Tumor tissues were collected from LL2 tumor-bearing mice of the SNAP treatment group and control group and were processed into a single-cell suspension. Single-cell suspensions from three mice in each group were mixed for further analysis. Tumor-infiltrating immune cells (CD45+ and 7-AAD-) were isolated from LL2 tumor-bearing mice of the SNAP treatment group (0.004 mg/kg) and control group (PBS containing 0.004% DMSO). CD45+ cells were stained with the BD sample tag, which carries a specific sequence, to identify the different groups during sequencing (Fig. 2A). A total of 9391 cells were analyzed in the two groups (4538 cells for the control group and 4853 cells for the low-dose SNAP group).
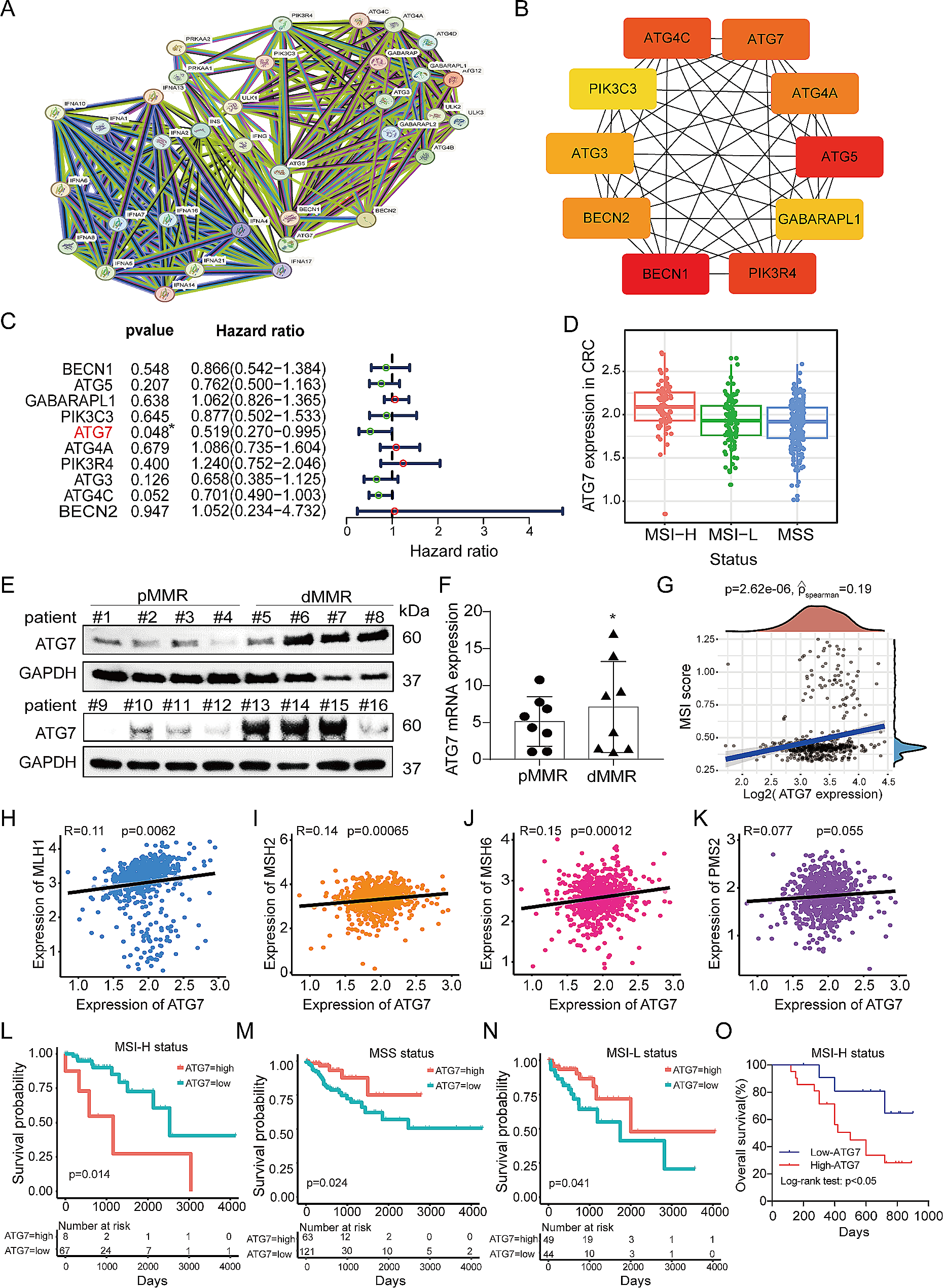
Fig. 2
The top 2 abundant infiltrating immune cells were tumor-associated macrophages and T cells. (A) Schematic of the study design. Single-cell RNA sequencing was used to analyze infiltrating immune cells isolated from tumor-bearing mice. (B) Clustering of intratumoral CD45+ cells and visualization of the subset. Uniform manifold approximation and projection (UMAP) of single-cell RNA sequencing data from 6563 cells (Control: 3387; Drug: 3176; upper) revealed six clusters determined by 14 specific markers (Supplementary Fig. 3). Each dot plot represents one cell. (C) Bubble heatmap determining the expression levels of specific marker genes in the CD45+ subsets and fibroblasts. (D) The percentages of CD45+ subsets and fibroblasts in the control and low-dose SNAP treatment groups
First, we classified cells into 11 clusters through clustering resolution selection (Supplementary Fig. 3 A). To identify cluster specificity, we categorized the marker genes and displayed the top five differentially expressed genes (DEGs) in each cluster (Supplementary Fig. 3 B and Supplementary Data 1). We further analyzed the gene expression intensities of 14 common markers to determine the immune cell type in these 11 clusters (Supplementary Fig. 3 C). Finally, we annotated each cluster and reduced the 11 clusters to 6 cell types (Fig. 2B) according to the intensity of the marker genes: tumor-associated macrophages (Clusters 0, 1, 2, and 3; Cd14+ and Cd68+), B cells (Cluster 4; Igkc+, and Cd79a+), T cells (Clusters 5, 8, and 9; Cd3e+ and Trac+), dendritic cells (Cluster 7; H2-Ab1+ and Itgax+), monocytes (Cluster 7; CD14+, S100a8+, S100a9+, and Fcgr1-) [31], and fibroblasts (Cluster 10; Cald1+ and Col3a1+). The expression levels of marker genes in the cluster were determined (Fig. 2C and Supplementary Fig. 3 D). We further analyzed the composition of CD45-positive cells between the control group and the low-dose SNAP treatment group. Tumor-associated macrophages (TAMs) were the major competent intratumoral immune cells in the two groups (69% and 68%). In addition, approximately 11-14% of cells were T cells (Fig. 2D). Our single-cell RNA sequencing results indicate that TAMs are the most abundant intratumoral immune cells. In addition, low-dose SNAP treatment did not remarkably alter the ratio of macrophage and T-cell populations in tumor microenvironments.
Low-dose SNAP treatment increases CD8+ T cells and natural killer (NK) cellsGiven that CD8+ T cells are one of the key immune cells responsible for antitumor immunity, we first aimed to study the effect of low-dose SNAP treatment on the T cell population. To efficiently identify CD4+ and CD8+ cells, we used CD4 and CD8 Ab-seq antibodies, which carry the antibody and a specific barcode for single-cell RNA analysis. To determine the populations of T cells, we subclustered the T lymphocytes that were identified by CD3e and Trac and used uniform manifold approximation and projection (UMAP) for dimension reduction (Supplementary Fig. 4 A). A total of six clusters were identified. There were two CD8+ T-cell clusters: Cluster 0, central memory T cells (Tcf7, and S1pr1) and Cluster 1, CD8+ cytotoxic T cells (Cd8a, CD8 (Ab), Gzmb, Nkg7, and Prf1); one CD4+ T-cell cluster: Cluster 2, CD4+ regulatory T cells (CD4 (Ab), Ctla4, Il2ra); two CD3e+ T cells: Cluster 4, proliferating T cells (Mki67, Top2a, Pclaf, Stmn1) and Cluster 5, CD14+ T cells; and Cluster 3, natural killer cells (CD3e-, Gzma, Nkg7, Prg1, and Gzmb) (Fig. 3A and Supplementary Figure 4B and C and Supplementary Data 2). Notably, during low-dose SNAP treatment, the population of Cluster 1 CD8+ cytotoxic T cells (Effector T cells; TE) increased from 21.8% to 24.2%, and Cluster 3 natural killer (NK) cells increased from 16% to 19%. In contrast, central memory T cells (TCM) in Cluster 0 were significantly decreased from 36.5% to 23.9% (Fig. 3B). Analysis of the top 5 expressed genes in TCM identified Cd69 (Supplementary Data 2); therefore, Cd69 and Tcf7 were analyzed in 5 T cell populations and NK cells. Coexpression of Tcf7 and Cd69 was observed only in TCM (Supplementary Fig. 5A), closely matching the exhausted T cell progenitor 1 (Texprog1) populations reported recently [32]. Additionally, biological process analysis of upregulated genes in Cluster 1 CD8+ cytotoxic T cells (avg_log2FC > 0.5) indicated that several gene groups were enriched in leukocyte activation and cytotoxicity (Fig. 3C). Moreover, KEGG pathway analysis suggested that the upregulated genes were related to the NK cell-mediated immune response (e.g., Gzmb, Prf1, Klrd1, Klrk1, Itgal, and Itgb2) (Fig. 3D). To further investigate which mechanisms were altered in Cluster 1 CD8 cytotoxic T cells after low-dose SNAP treatment, 1117 DEGs derived from the low-dose SNAP group (Supplementary data 3) were used for gene set enrichment analysis (GSEA). GSEA results indicated that the DEGs of Cluster 1 CD8+ cytotoxic T cells exposed to low-dose SNAP treatment were enriched in negative regulation of hydrolase activity (Fig. 3E). GSEA results implied that hydrolase inhibition might correlate with the low-dose SNAP-induced antitumor response. Indeed, previous studies indicated that T cell activation resulted in a decrease in adipocyte triglyceride lipase, a triacylglycerol hydrolase [33]. These results suggested that low-dose SNAP increased the number of Cluster 1 CD8+ cytotoxic T cells and that this cluster exhibited an expression pattern associated with potentially increased cytotoxicity. GSEA results also suggest that low-dose SNAP treatment might regulate hydrolase activity to enhance T cell activation. On the other hand, GSEA analysis of 937 DEGs in NK cells indicated cluster 5 NK cells contained the interferon-gamma response gene signature (Supplementary Fig. 5B and supplementary data 4).
Fig. 3
Low-dose SNAP increases CD8+ effector T cells, and the cytotoxicity pathway is enriched in these cells. (A) Subclustering of T lymphocytes and visualization of the subsets. Dimension reduction by UMAP displaying six subsets of T lymphocytes (814 cells). (B) Distribution of each subset of T lymphocytes in the control and SNAP treatment groups. (C) Biological process and (D) KEGG pathway analyses of 92 DEGs (avg_log2-fold change > 0.5) in CD8+ T cells. The GeneRatio indicates the number of significant genes associated with the Gene Ontology (GO) term divided by the total number of genes in the related pathway in the database. (E) MSigDB ontology gene set analysis of 1117 differentially expressed genes (DEGs) in Cluster 1 CD8+ cytotoxic T cells after low-dose SNAP treatment; the list of DEGs is shown in the Supplementary Data. 3. DEGs were identified using the “FindMarker” function in the Seurat package. The red bar represents the upregulated pathway after low-dose SNAP treatment. All pathways were statistically significant (p-value < 0.05). The p-value of GSEA was calculated by permutation
CD8+ T cell-mediated immunity is required for tumor inhibition by low-dose nitric oxide donorsImmunosuppressive cytokines, such as IL-6 and IL-10, interfere with the antitumor effects of CD8+ and CD4+ T cells in the tumor microenvironment [34, 35]. Low-dose SNAP (0.004 mg/kg) treatment decreased IL-6 and IL-10 secretion by splenocytes (Supplementary Fig. 2E). Moreover, single-cell RNA sequencing analysis indicated that low-dose SNAP treatment altered CD8+ T cells with activation gene signatures. Therefore, we further analyzed the percentages of CD8+ T cells in the tumor microenvironment after low-dose SNAP treatment using flow cytometry. Mice were treated with low-dose SNAP (0.004 mg/kg) and sacrificed on the 20th day after LL2 tumor implantation. The low-dose SNAP group exhibited smaller tumor sizes and weights than the control group (Fig. 4A). In addition, another NO donor, ISMN, also reduced tumor volume and weight upon low-dose treatment (0.004 mg/kg) (Fig. 4B). Infiltrating CD8+ T cells were increased by low-dose SNAP or ISMN (Fig. 4C and D). These results demonstrated that an increase in CD8+ T cells may play a role in the antitumor effect induced by SNAP or ISMN treatment.
Fig. 4
CD8+ T cells are essential for the repression of tumor growth mediated by low-dose SNAP. (A) Tumor tissues were isolated from LL2 tumor-bearing mice after SNAP treatment (left), and the tumor sizes (middle) and the tumor weights (right) were analyzed on the 20th day. (B) Tumor tissues were isolated from LL2 tumor-bearing mice after ISMN treatment (left), and the tumor sizes (middle) and the tumor weights (right) were analyzed on the 20th day. (C) FACS analysis of intratumoral CD8+ T cells between the control group and low-dose SNAP treatment group. (D) FACS analysis of intratumoral CD8+ T cells between the control group and the low-dose ISMN treatment group. The mice were sacrificed, and tumor-infiltrating lymphocytes were analyzed on the 20th day after implantation of LL2 tumor cells. (E) Violin plot revealing Ccr7 expression levels in dendritic cells. Each dot represents one cell. (F) Flow cytometry analysis of intratumoral mature dendritic cells between the control group and low-dose SNAP treatment group. (G) Schematic of CD8+ T-cell depletion in mice (upper). Representative histogram of CD8+ expression on splenocytes (lower). (H) Simplified diagram of CD8+ T-cell depletion in the LL2 tumor-bearing mice (upper). LL2 tumor-bearing mice were treated with low-dose SNAP (0.004 mg/kg) and anti-CD8 antibody (200 μg/time) (lower). Anti-IgG antibody was used as the control. Both antibodies and SNAP were administered via intraperitoneal injection. The column scatter dot plot represents the mean values ± SEMs. *p < 0.05, **p < 0.01, versus the control group. P-values of the tumor results and violin plots were obtained by two-way ANOVA and the Wilcox test, ***p < 0.001, ns, no significant difference. The tumor volumes were measured every 2 days beginning on the 10th day after tumor implantation
To investigate the potential mechanisms by which low-dose SNAP regulates CD8+ T cell activation, we collected the expression profiles in the subset of T lymphocytes (Clusters 0, 1, 2, 4, and 5), excluding Cluster 3 NK cells. We further gated CD8+ T cells (n = 302) with Ab-seq antibodies (CD8 (Ab) and CD4 (Ab)) using SeqGeq software, a platform for single-cell analysis (Supplementary Fig. 6A). In addition, we selected 338 significant genes among 8369 genes in CD8+ T cells (average expression > 0, p value < 0.05, percentage of expressing cells > 10, SNAP/Control fold change >1.5; Supplementary Data 5) for pathway analysis by Metacore software. Alterations in gene expression in CD8+ T cells were highly related to the Wnt/beta-catenin pathway (Supplementary Fig. 6B), which was recently reported to regulate CD8+ T-cell activation [36]. Therefore, we further isolated intratumoral CD8+ T cells by magnetic beads and measured the mRNA levels of beta-catenin-related genes by quantitative real-time polymerase chain reaction (RT‒qPCR). Beta-catenin (Ctnnb1) was significantly downregulated, and its targeted transcriptional factor 7 (Tcf7) was slightly downregulated upon treatment with low-dose SNAP (Supplementary Fig. 6C and D). In addition to the Wnt/beta-catenin pathway, cross-presentation by mature dendritic cells also contributes to CD8+ T cell activation [37], and the expression of C-C chemokine receptor 7 (Ccr7) in conventional dendritic cells (cDCs) plays an important role in delivering intact tumor antigen to tumor-draining lymph nodes [38] and is also a marker in the maturation of dendritic cells [39]. Single-cell RNA sequencing data revealed that Ccr7 expression levels in dendritic cells were increased after low-dose SNAP treatment (Fig 4E). Therefore, we next examined whether low-dose SNAP increased the number of mature and activated dendritic cells in the tumor microenvironment. Flow cytometry analysis indicated that the number of cells double-positive for the dendritic cell marker CD11c and maturation marker CD86 was increased after low-dose SNAP treatment (Fig 4F). A previous study indicated that nitric oxide treatment induced immunogenic cell death (ICD) to control tumor progression [20]. ICD is a process in which dying cells release damage-associated molecular patterns (DAMPs) to activate the maturation of dendritic cells and induce CD8+ T cell activation to kill cancer cells [40]. To investigate whether the low-dose SNAP-mediated antitumor effect was correlated with ICD, we isolated tumor cells from LL2 tumor-bearing mice through CD326 magnetic beads and measured levels of the ICD marker, surface calreticulin. However, surface calreticulin was only weakly enhanced by low-dose SNAP treatment (Supplementary Fig. 7). These results demonstrated that low-dose SNAP treatment increased dendritic cell maturation and CD8+ T cells in the tumor microenvironment. To further determine whether CD8+ T cells play an essential role in the antitumor response, CD8+ T cells were depleted with an anti-CD8 antibody. Two doses of anti-CD8 antibody depleted approximately 90% of CD8+ T cells, as demonstrated by flow cytometry (Fig. 4G). The low-dose SNAP-mediated antitumor response was almost abolished by CD8+ T cell depletion (Fig. 4H). These results demonstrated that CD8+ T cells were required for the antitumor effects induced by low-dose SNAP.
The results above indicated that the low-dose SNAP-mediated antitumor response was dependent on CD8+ T cells and correlated with mature dendritic cells. In addition, SNAP treatment influenced the Wnt/beta-catenin pathway of CD8+ T cells, which may be related to CD8+ T-cell activation.
Low-dose SNAP increased a TAM subtype that coexpressed M1- and M2-related markersNO is an important molecule for macrophage differentiation [41]; therefore, low-dose NO donor SNAP treatment may regulate TAM polarization. To further investigate whether low-dose SNAP treatment modulated TAM differentiation, we analyzed alterations in the expression levels of M1 and M2 macrophage markers. TAMs were subclustered into 7 clusters based on the following markers: TAM1, Ppbp, Arg1, and Pf4; TAM2, Cd74, H2-Aa, and H2-Ab1; TAM3, C1qa, C1qb, and C1qc; TAM4, Il1a, Cxcl2, and Il1b; TAM5, Ifit47, Ifit3, and Ifit1; TAM6, Sell, Plac8, and Chil3; and TAM7, Atf3, Ubc, and Rhob (Fig. 5A). TAM1 expressed M2 markers Arg1 and Mrc1. TAM2 expressed major histocompatibility complex 2 (MHC-II) and macrophage migration inhibitory factor (MIF) receptor CD74. TAM3 expressed the complement component 1 complex genes C1qa, C1qb, and C1qc TAM4 expressed the proinflammatory genes Il1a, Il1b, and Ptgs2. TAM5 expressed the interferon-stimulated genes Rsad2, Isg15, and Ifit1. TAM6 expressed Plac8 and Chil3. TAM7 expressed Klf2 and Egr1 (Supplementary Fig. 8).
Fig. 5
Low-dose SNAP treatment induces alterations in macrophage subsets. (A) Subclustering of tumor-associated macrophages (TAMs) by UMAP and identifying the top three DEGs in each subset of TAMs. The top three DEGs were identified using the “FindAllMarker” function in the Seurat package. (B) The distribution of each TAM defined by gene expression patterns in the control or low-dose SNAP treatment groups. (C) The expression levels of M1-related and M2-related genes in the TAM subset. (D) Ridge plots determining the expression distributions of M1-related genes (Cxcl3) and M2-related genes (Arg1) in the TAM subsets. The X-axis represents log-normalized expression levels
To investigate the effect of low-dose SNAP on the TAM subtype, we calculated the proportion of each TAM subset. Low-dose SNAP dramatically altered the amounts of two TAM subsets, TAM1 and TAM2 (Fig. 5B). TAM1 was increased from 18.4% to 25.4% after low-dose SNAP treatment, whereas TAM2 was decreased from 23.9% to 16.1%. Given that M1 and M2 macrophages frequently exert opposite effects on tumor progression, we analyzed M1-related genes and M2-related genes in the TAM subtypes [42,43,44,45,46,47,48]. TAM4 cells expressed more M1-related genes (Ccl3, Nos2, and Il1b), and TAM3 cells exhibited higher mRNA expression of M2-related genes (Apoe, C1qa, C1qb, and C1qc). However, TAM2 cells exhibited reduced mRNA expression levels of M1- and M2-related genes (Fig. 5C). Of note, low-dose SNAP treatment increased a particular subtype of macrophages (TAM1) that coexpressed M1- and M2-related genes, such as Cxcl3 and Arg1 (Fig. 5D), suggesting that low-dose SNAP regulated the inflammatory response in TAMs.
Coexpression of the M1-like gene expression signature and Arg1 in a subpopulation of macrophages induced by SNAPArg1-expressing M2 macrophages in general play an immunosuppressive role through the consumption of extracellular L-arginine [49]; however, CD206-expressing M2 macrophages can be converted into cells that enhance adaptive and innate antitumor immune responses [50]. Our single-cell analysis revealed that low-dose SNAP increased the proportion of TAM1 cells that coexpressed M1- and M2-related genes (Fig. 5C). In addition, TAM1 cells did not express IL-10 (Supplementary Fig. 9), which is a key cytokine associated with immunosuppression. Therefore, it is important to further characterize the unique macrophage subpopulation induced by low-dose SNAP. We subclustered TAM1 cells, and 5 subsets were identified (TAM1-0 to TAM1-4) (Fig. 6A). The top 5 averaged genes in each subtype of TAM1 were calculated and visualized in a heatmap (Fig. 6B). TAM1-0 cells expressed Pf4 (Cxcl4) and Selenop, which induced M2 macrophage differentiation [51], and TAM1-1 cells had high expression of heat shock protein family A members (Hspa1a and Hspa1b) and prostaglandin-endoperoxide synthase 2 (Ptgs2). TAM1-2 cells expressed higher levels of thrombospondin 1 (Thbs1). TAM1-3 cells expressed MHC-class II molecules (H2-Eb1, H2-Aa, and H2-Ab), and TAM1-4 cells exhibited increased prostaglandin reductase 1 (Ptgr1) expression. Notably, TAM1-1 cells had lower Mrc1 (CD206) expression levels and higher expression levels of Ptgs2, a gene involved in the inflammatory response in M1 macrophages [52]. Low-dose SNAP treatment reduced the TAM1-0 population but increased the TAM1-1 population. On the other hand, the TAM1-2, TAM1-3, and TAM1-4 populations were not affected by SNAP (Fig. 6C). To further determine whether M1-related genes were expressed in Arg1-expressing macrophages, we analyzed M1- and M2-related genes in subsets of TAM1 cells. TAM1-0 cells expressed more M2 macrophage-related genes, and TAM1-1 cells exhibited increased expression of M1 macrophage-related genes (Fig. 6D). Indeed, flow cytometry analysis also indicated that low-dose SNAP treatment increased the population of Arg1+ IL7R+ macrophages and Arg1+ CD80+ macrophages (Fig. 6E and F). These data suggested that low-dose SNAP treatment decreased M2-like macrophages (TAM 1-0). In addition, a unique subset of Arg1-expressing macrophages (TAM1-1) that highly expressed M1-related genes was significantly increased after low-dose SNAP treatment.
Fig. 6
Low-dose SNAP induces a subset of macrophages displaying an M1-related gene expression signature and Arg1 expression. (A) Subclustering of TAM1 (978 cells) and visualization of the subsets. UMAP embedding of single-cell RNA sequencing data revealed five clusters using the “FindCluster” function. Each dot represents one cell. (B) Average expression levels of the top 5 DEGs in each subset of TAM1 cells. (C) Distribution of each TAM1 subset in the control and SNAP treatment groups. (D) Bubble heatmap showing the expression levels of M1-related genes and M2-related genes in the subset of TAM1 cells. (E) Flow cytometry analysis of intratumoral TAM1-1 cells and (F) CD80+ Arg1+ macrophages between the control and low-dose SNAP treatment groups. (G) MSigDB hallmark gene set enrichment assay (GSEA) analysis of DEGs in the TAM1-0 (left) and TAM1-1 (right) clusters. TAM1-0 and TAM1-1 DEGs were determined using the “FindMarker” function in the Seurat package and are displayed in Supplementary Data 6. All pathways were significant (p-value < 0.05). Tumor tissues were collected Day 20 after tumor implantation and used for flow cytometry analysis
To further understand the potential roles of TAM1-0 and TAM1-1 in the tumor microenvironment, DEGs were used (800 genes in TAM1-0 cells, 991 genes in TAM1-1 cells) for hallmark gene set enrichment analysis (GSEA) (Supplementary Data 6). TAM1-0 cells had enhanced levels of coagulation and complement pathways and reduced levels of glycolysis, hypoxia, and TNF-α signaling and IFN-γ response pathways. TAM1-1 cells exhibited opposite trends based on GSEA. TAM1-1 cells displayed enrichment in proinflammatory response pathways, including TNF-α signaling, the IFN-γ response, and IL-6-JAK-STAT3 signaling (Fig. 6G). Based on GSEA, TAM1-0 cells may function as protumor macrophages; on the other hand, TAM1-1 macrophages may possess antitumor properties. Therefore, the antitumor response of low-dose SNAP was correlated with a decrease in protumor macrophages and an increase in a unique set of Arg1-positive macrophages with an antitumor gene expression signature.
Low-dose SNAP enhanced the therapeutic effect of cisplatin in tumor-bearing mouse modelsFinally, combination treatment with chemotherapeutic drugs and immunomodulation agents is frequently employed in cancer patients to prolong their survival [53]. For example, immune checkpoint blockade plus cisplatin has been used to treat NSCLC [54]. Our results indicated that low-dose SNAP inhibited tumor growth via regulation of the immune system; therefore, we further tested the concept that low-dose SNAP treatment might enhance the therapeutic effect of the chemotherapeutic drug cisplatin. Mice were treated with low-dose SNAP (0.004 mg/kg) and/or cisplatin (5 mg/kg), and tumor growth was measured. The combination treatment protocol is illustrated in Fig. 7A. In the LL2 tumor-bearing mouse model, a combination of low-dose SNAP and cisplatin enhanced the cancer therapeutic effect and prolonged survival compared with cisplatin or low-dose SNAP alone (Fig. 7B and C). Similarly, a low dose of SNP, another nitric oxide donor, enhanced the therapeutic effect and extended the survival of tumor-bearing mice (Fig. 7D and E). To further investigate whether the effective combination therapy effect can be observed in another mouse strain, BALB/C mice were implanted with CT26 colon tumor cells and treated with the same doses of SNAP, cisplatin, or both. Increased antitumor effects were observed in the CT26 tumor-bearing mouse model upon combination treatment with low-dose SNAP and cisplatin (Fig. 7F). These results indicated that low-dose SNAP enhanced the antitumor efficacy of a common chemotherapeutic drug, cisplatin. To investigate the potential mechanism responsible for the enhancement of therapeutic effects in the combination study, we performed single-cell RNA sequencing on live tumor-infiltrating immune cells from four groups (control, low-dose SNAP, cisplatin, and combination). A total of 6 clusters (TAMs, monocytes, dendritic cells, B cells, T cells and fibroblasts) were identified based on DEGs and 14 common markers (Supplementary Fig. 10A and B). T cells and NK cells were clustered into 5 clusters based on DEGs: CD3+ CD4- CD8- double negative T cells, CD4 regulatory T cells (Cd4, Il2ra, and Foxp3), CD8 cytotoxic T cells (Cd8, Gzmb, and Ifng), proliferating T cells (Mki67, Top2a, and Pclaf), and NK cells (Cd3e negative, Klrk1, Nkg7, and Gzma), and alterations in these clusters were assessed among the four groups of mice (Supplementary Fig. 10C). Combination treatment with low-dose SNAP and cisplatin increased the number of NK-annotated cells based on gene expression patterns. Similar percentages of CD8+ cytotoxic T cells were observed between the cisplatin group and the combination group. However, interestingly, combination treatment inhibited the androgen response in CD8+ cytotoxic T cells in GSEA (Supplementary Fig. 10D), which may inhibit CD8+ T-cell function [55]. These results demonstrated that an increase in NK cells is associated with an enhanced therapeutic effect in combination treatment. Additionally, the additive effect of combination treatment might correlate with the downregulation of the androgen response in CD8+ T cells.
留言 (0)