
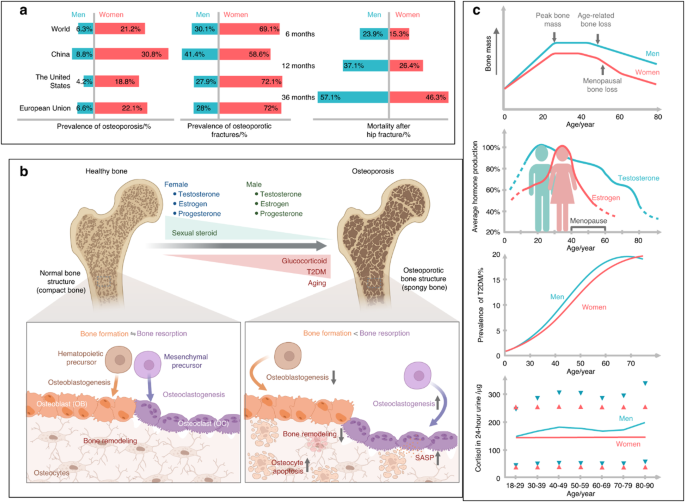
記住我

Normal CB tissues were obtained from nine surgical patients with degenerative disc disease. Altogether, 12 458 cells identified in the CB samples were subjected to further analysis. Single-cell transcriptomes of six conventional OS samples were obtained from the GEO dataset GSE152048.20 After quality filtering, we obtained 45 238 cells for subsequent analysis. The cells were classified into six distinct cell lineages annotated with canonical marker gene expression. As a result, mesenchymal, T, B, myeloid, osteoclast, and endovascular cells were identified (Fig. 1a and Fig. S1a). The distribution of the cells from each sample in the uniform manifold approximation and projection (UMAP) is shown in Fig. 1b and Fig. S1b. Compared with cells in CB tissues, mesenchymal and myeloid cells were increased and T and B cells decreased in OS samples (Fig. S1c, d).
Fig. 1
Single-cell atlas of osteosarcoma (OS) and normal cancellous bone (CB) samples. a UMAP plot of OS and CB transcriptomes, color-coded for six phenotypes identified by graph-based clustering. b UMAP plot color-coded for each OS and CB sample. c UMAP plot of T and B cells, color-coded for eight phenotypes identified by graph-based clustering. d UMAP plot of myeloid cells, including osteoclasts color-coded for five phenotypes. e UMAP plot of endovascular cells color-coded for two phenotypes. f UMAP plot of mesenchymal cells color-coded for 11 phenotypes and the UMAP plot of CSC-like clusters color-coded for 4 phenotypes. g Heatmap of expression of subset-specific markers across cell subsets. h Reproducible cell subset distributions across samples. Fractions of cells in each cluster derived from OS (red) or CB (blue) samples are shown
Clustering analysis was further performed for each cell type. T cells were divided into five clusters: regulatory T cells (T-regs), CD4+ T cells, CD8+ T cells, natural killer T (NKT) cells, and dendritic cells (DCs). B cells were mainly composed of three different subgroups: naïve B cells, active B cells, and plasma cells (Fig. 1c). Myeloid cells, including osteoclasts, fell into five subsets: monocytes, M1 macrophages (M1Φs), M2Φs, tumor-associated macrophages (TAMs), and osteoclasts (Fig. 1d). Two clusters (endothelial and mural cells) were identified among endovascular cells (Fig. 1e). Mesenchymal cells were divided into BMSCs, osteoblasts, adipocytes, chondroblasts, and seven OS tumor cell clusters, which we named CSC-like, OS-A1, OS-A2, OS-B1, OS-B2, OS-C1, and OS-C2. The CSC-like clusters were further subdivided into four subclusters (CSC, CSCL1, CSCL2, and CSCL3) based on clustering (Fig. 1f). Thus, 29 clusters were revealed in CB and OS tissues.
Each cluster exhibited a distinct gene expression pattern (Fig. 1g and Table S1). Correlation analysis showed that clusters from the same cell lineage had higher similarity than those from other cell lineages (Fig. S1e), confirming the reliability of the clusters. The distribution of unique molecular identifiers in each cell lineage is shown in Fig. S1f. Each cluster included cells from multiple patients, showing clear distribution differences between OS and CB samples (Fig. 1h). The proportion of all cell clusters in each sample is also provided in Fig. S1d.
The distinct transcriptome program in mesenchymal cellsMesenchymal cells are the main constituent of OS samples (Fig. S1c), which is consistent with the mesenchymal origin of OS.21 Using the Monocle3 method, we observed that OS tumor cells included a CSC-like cluster and three branches with two clusters in each branch (Fig. 2a). Pseudotime trajectory analysis (monocle3 method) and velocity analysis both demonstrated the CSC-like cluster to be the origin of each branch during cell differentiation (Fig. 2b and Fig. S2a). We found that the clusters from each branch observed by the monocle3 method were also concentrated in different differentiation branches of the monocle2 method (Fig. S2b). Interestingly, OS cells from various samples were enriched in distinguished branches (Fig. 2c), indicating the heterogeneity of the OS samples. Cell proportion analysis demonstrated the OS cell clusters mainly derived from OS samples and the normal mesenchymal clusters from CB samples (Fig. 2d).
Fig. 2
Differences in cell composition and gene expression between OS and CB samples. a UMAP plot of mesenchymal cells by the Monocle3 method. b Pseudotime trajectory of mesenchymal cells. c UMAP plot of mesenchymal cells color-coded for each sample. d Differences in cell proportion between OS and CB samples in each mesenchymal cluster. e Dot-plot heatmap of the most significant genes of each cluster in mesenchymal cells. f Violin plots showing differential expression of stem cell, osteoblast, chondrocyte, adipocyte, and sarcoma markers in each mesenchymal cluster. g Differences in gene expression between CSC-like and BMSC clusters. h Comparison of gene expression between CSC-like and osteoblast clusters. i Differences in gene expression between the CSC-like cluster and total OS cells. j Differences in gene expression between total OS cells and osteoblasts. *P < 0.05
The most significant markers of each mesenchymal cluster are shown in Fig. 2e and Table S2, demonstrating clear distinctions between the clusters. Further analyses showed the stem cell markers to be highly expressed in BMSCs and CSC-like clusters and partly expressed in OS-A1/A2/B1/B2 clusters (Fig. 2f), suggesting that the OS cells in branch-A/B seemed to have stronger stemness than those in branch-C. In addition, chondrocyte and adipocyte markers were expressed at low levels in all OS cell clusters; OB markers ALP, THY1, and COL1A1 were expressed in OS cell clusters (Fig. 2f), suggesting the osteoblastic nature of OS cells.22 We observed that ALP and COL1A1 were upregulated in OS cells compared with OBs, indicating enhanced extracellular matrix formation ability in OS cells, which might induce abnormal osteogenesis in OS.22,23 Sarcoma markers were differentially expressed in OS cell clusters (Fig. 2f), suggesting the heterogeneity of the three branches. Some of the gene expression changes during the OS cell differentiation trajectories are shown in Fig. S2c. For example, MYC and CYR61 were upregulated during differentiation trajectory-A; CDK4 was gradually increased in branch-B, and expression of TIMP3 and MMP13 was gradually enhanced in branch-C.
We further compared gene expression between OS and normal mesenchymal cells and found that CSC-like cells expressed higher levels of the osteoblastic markers COL1A1, COL1A2, and SPARC than BMSCs (Fig. 2g and Table S3). Compared with OBs, CSC-like cells overexpressed SPP1 and SERPINA1 genes related to tumor progression (Fig. 2h and Table S4). Compared with other OS cells, CSC-like cells highly expressed chemoresistance genes, including FTL, XIST, and MT1G (Fig. 2i and Table S5). Overall, osteoblastic marker expression exhibited limited diversity between OS cells and OBs (Fig. 2j and Table S6). A comparison of gene expression between BMSCs and other normal mesenchymal cells (including adipocytes, chondroblasts, and OBs) is shown in Fig. S3a–c and Tables S7–9.
Specific molecular features of the three differentiation branches of OS cellsWe performed GSVA in the seven clusters of OS cells to further analyze the features of the three branches of OS cells (Fig. S2d). Total GO pathway enrichment increased gradually in the CSC-like, OS-A1/B1/C1, and OS-A2/B2/C2 clusters (Fig. 3a). Because quiescence is a critical feature of stem cells,24 this result suggests the primitiveness of the CSC-like cluster and OS-A1/B1/C1 clusters as the primary stage during OS cell differentiation.
Fig. 3
Transcriptional distinction between the three differentiation trajectories of OS cells. a Differences in total GO pathway enrichment between the seven OS clusters. b Differences in the proportion of cell cycle phases in the 11 mesenchymal clusters. c Differences in gene expression between the OS-A2 and OS-B2 clusters. d Differences in gene expression between the OS-A2 and OS-C2 clusters. e Differences in expression between the OS-B2 and OS-C2 clusters. f Heatmap showing differences in the activation of pathways related to targeted therapies in OS-A2/B2/C2 clusters calculated by the GSVA method. g Heatmap showing differences in activation of pathways between the seven OS clusters. h Heatmaps of differences in single-cell copy number between the seven OS clusters and normal endovascular cells. i Heatmap showing differences in TF activity between the seven OS clusters. j TF activity in the seven OS clusters. The top 8 activated TFs were marked in each cluster
We further studied each cluster’s cell cycle stage distribution and found that BMSCs and CSC-like cells were enriched in the S and G2/M stages, indicating activation of cell proliferation, which is consistent with their stem cell nature (Fig. 3b). Interestingly, we found that OS-B1/B2 cells were concentrated in the S stage but that OS-C1/C2 was mainly in the G1 stage (Fig. 3b), indicating that OS cells in branch-B might have strong proliferative activity and that the proliferation of cells in branch-C may be possibly slow. The differentially expressed genes in the OS-A2/B2/C2 clusters are illustrated in Fig. 3c–e and Tables S10–12. We observed that several bone metabolism-related genes, including SPP1, IBSP, and MMP13, were upregulated in the OS-C2 cluster compared with the other two clusters.
When focusing on specific pathway activation among the OS cell clusters, CSC-like and OS-A1 exhibited stronger pluripotency than other clusters, and pathways related to osteoblast development and bone morphogenesis were activated in OS-A2/B2/C2 clusters, possibly revealing the maturation of these clusters (Fig. 3g). In addition, we analyzed pathways related to targeted therapies in OS-A2/B2/C2 clusters and found obviously different pathway enrichment between the OS-A2/B2/C2 clusters, such as activation of cell cycle- and ERBB2-related pathways in OS-B2 and enrichment of bone resorption and immune activation pathways in OS-C2 (Fig. 3f). Expression levels of target genes involved in targeted therapies were also diverse in the OS-A2/B2/C2 clusters. For example, VEGFRs (FLT1 and KDR) and CDK4 were overexpressed in the OS-B2 cluster, and PD-L1 (CD274) was upregulated in the OS-C2 cluster (Fig. S2e). These results suggest that OS cells in distinct differentiation branches might be sensitive to different drugs.
Differences in gene expression in cells are often caused by changes in chromosome copy number variation (CNV). According to the CNV calculation results based on scRNA-data, we found that CSC-like cells had different CNVs compared with other OS clusters. In addition, the clusters in the same differentiation branch exhibited similar CNVs, but different differentiation branches had distinct CNVs. For example, extensive chromosomal gains were observed in 6p, 8q, 16q, 17p, and 19p of branch-A clusters, 3p, 6, 10, and 19 of branch-B clusters, and 3p, 9q, and 19 of branch-C clusters. Extensive chromosomal losses were found in 10, 12q, and 19q of branch-A clusters, 8q, 12p, 14, 20, and 22 of branch-B clusters, and 12q of branch-C clusters (Fig. 3h). These results highlight the heterogeneity between the three differentiation branches of tumor cells in OS.
We performed transcription factor (TF) analysis for the seven OS cell clusters to reveal the transcriptional program during OS cell differentiation. The results highlighted the relative activation of EZH2 and CTCF in CSC-like cells (Fig. 3i), which correlated with tumor cell stemness, proliferation, and drug resistance.25,26 Moreover, clusters in the same differentiation branch shared similar TF activation, whereas those in different branches exhibited different TF activation, demonstrating that the transcriptional programs in the three differentiation trajectories are different (Fig. 3i, j). TF activation of the four normal mesenchymal clusters (BMSCs, osteoblasts, chondroblasts, and adipocytes) is shown in Fig. S3d, e.
OS classification based on the differentiation branches of OS cellsDue to the clear differences in gene expression, TF activation, and CNV between the three branches of OS cells, we considered that the OS samples could be classified into three groups based on the differentiation branches of tumor cells. The intersections between the marker genes of OS-A2/B2/C2 were quite limited (Fig. 4a), suggesting that OS-A2/B2/C2 markers can be used to identify differentiation branches in OS samples. We selected the 44 most specific markers of the OS-A2/B2/C2 clusters, including 24 markers for OS-A2, 9 for OS-B2, and 11 for OS-C2 (Fig. 4b). Interestingly, we observed that the markers of OS-B2 correlated significantly negatively with the survival of OS patients based on the TARGET dataset (Fig. 4d); conversely, the markers of OS-A2 and OS-C2 predicted good prognosis in OS patients (Fig. 4c, e).
Fig. 4
Classification of OS samples from the TARGET dataset based on the markers of the three differentiation branches of OS cells. a Venn diagram showing the similarities and differences of the calculated markers of OS-A2/B2/C2 clusters. b Heatmap showing the expression of the 44 selected marker genes of OS-A2/B2/C2 in all 29 clusters. Kaplan–Meier curves of overall survival for patients with different expression levels of total OS-A2 (c), OS-B2 (d) or OS-C2 (e) markers in OS patients from the TARGET dataset. f Consensus clustering matrices with the 44 marker genes of OS-A2/B2/C2 in TARGET OS samples for k = 3. g Heatmap showing the expression of 44 marker genes in the three clustered groups of TARGET OS samples. h Kaplan–Meier curves of overall survival for the three clustered groups of TARGET OS patients. i Heatmap showing activation differences of pathways related to targeted therapies in the three subgroups of TARGET OS samples. j Heatmap showing expression levels of genes related to targeted therapies in the three subgroups of TARGET OS samples
To explore the possible application of OS-A2/B2/C2 markers in OS sample classification, we first divided 88 OS samples in the TARGET dataset into three groups by resampling selected tumor profiles using the 44 selected markers for OS-A2/B2/C2. The three subtypes achieved a good discrimination effect for OS samples (Fig. 4f), and the markers of OS-A2/B2/C2 were overexpressed significantly in Group A/B/C samples (Fig. 4g). Consistent with the relevance between the markers of OS clusters and the prognosis of OS patients, Group B patients exhibited a significantly worse survival rate than the other two groups (Fig. 4h). Pathway enrichment analysis showed that pathways related to targeted therapies were differentially activated in the three groups of OS samples, which was partly similar to the pathway activation in the OS-A2/B2/C2 clusters, such as activation of cell cycle pathways in Group B and activation of bone resorption and immune activation pathways in Group C (Fig. 4i and Table S13). The target genes of targeted therapies were also differentially expressed in the three group samples: VEGFRs (FLT1, KDR, FLT3, and FLT4) and CDK4 were overexpressed in Group B samples, and PD-L1 (CD274) was upregulated in the Group C cluster (Fig. 4j). These findings suggest that the classification system based on OS-A2/B2/C2 markers may be used to guide clinical treatment for OS patients.
Verification of the classification system in clinical OS samplesTo further examine the clinical significance of the classification system in OS samples, we performed Immunohistochemical (IHC) staining for six gene makers selected from the OS-A2/B2/C2 clusters (OS-A2: ALKBH5 and TOM1L2; OS-B2: CDK4 and LMO7; OS-C2: COL6A3 and THBS2) in 138 osteoblastic OS samples (Fig. 5a). OS patients were divided into three groups according to the strongest expressed marker of each sample. Groups A/B/C highly expressed the markers of the OS-A2/B2/C2 clusters, which parallels Group A/B/C in the TARGET OS cohort. We observed some correlations between the expression of markers for the same cluster (Fig. 5b). By using K-M curves, we found that the OS patients in Group B exhibited significantly worse overall survival than those in the other two groups (Fig. 5c), similar to the survival analyses in the TARGET dataset (Fig. 4h). Except for alive status, no significant difference in clinical characteristics was observed among the three groups of OS patients (Table S14). These results suggest that the classification of OS samples can be performed through IHC staining, which may help in the prognostic evaluation of OS patients.
Fig. 5
Group classification of OS samples based on IHC staining. a IHC staining of ALKBH5, TOM1L2, CDK4, LMO7, COL6A3, and THBS2 in three OS samples from each group (No. 13, No. 68, and No. 79 of OS samples). b Statistical analysis of expression of six gene markers in each OS sample. c Kaplan–Meier curves of overall survival for the three groups of OS patients
Identification and characterization of OS stem cellsAs CSCs play a critical role in tumor development and progression,27 we further studied the composition of the CSC-like cluster and found that they can be further classified into four clusters: CSC, CSC-like 1 (CSCL1), CSCL2, and CSCL3 (Fig. 6a). Pseudotime trajectory analysis identified three differentiation branches of CSCs that correlated with CSCL1, CSCL2, and CSCL3 (Fig. 6b). The CSC cluster contained 369 cells, which accounted for 0.82% of all cells in OS (Fig. 6d). CSCL1 cells were mainly from samples enriched in OS-A1/A2 cells (OS1 and OS2), over 50% of CSCL2 cells were from samples enriched in OS-B1/B2 cells (OS3 and OS6), and most CSCL3 cells were from samples enriched in OS-C1/C2 cells (OS4 and OS5) (Fig. 6c). In addition, the gene markers of CSCL1/2/3 were partly similar to OS-A2/B2/C2 clusters, such as MT1G and SERPINA1 in CSCL1, as well as SPP1 and MMP13 in CSCL3 (Figs. 2e and 6h and Table S15). These results suggest that CSCL1/2/3 might be the progenitors of OS-A2/B2/C2.
Fig. 6
Molecular features of OS stem cell CSC-like cells. a UMAP plot of CSC-like cells using the Monocle3 method. Four subclusters were identified in CSC-like cells by graph-based clustering. b Pseudotime trajectory of CSC-like cells. c UMAP plot of CSC-like cells color-coded for each OS sample. d Mean cell numbers of the four subclusters of CSC-like cells in OS samples. e Total GO pathway enrichment among the CSC, CSCL1, CSCL2, CSCL3, and OS-A1/B1/C1 clusters. f The cell proportion of each cell cycle phase in four subclusters of CSC-like cells. g Violin plots showing expression of proliferative markers and stem cell markers in the four subclusters of CSC-like cells. h Dot-plot heatmap of the most significant genes in the four subclusters of CSC-like cells. i Differences in gene expression between CSCs and other CSC-like subclusters. j Differences in gene expression between CSCL1 and other CSCL2 clusters. k Differences in gene expression between CSCL1 and other CSCL3 clusters. l Differences in gene expression between CSCL2 and other CSCL3 clusters. m TF activity in the four subclusters of CSC-like cells. The top 8 activated TFs are marked in each cluster
Total GO pathway enrichment in four CSC-like clusters was lower than that in OS-A1/B1/C1 (Fig. 6e). Cell cycle analysis showed the CSC cluster to be arrested in G2/M phase compared with the CSCL1/2/3 clusters (Fig. 6f). As G2/M arrest has been found in some cancers and CSCs, we considered that the G2 checkpoint is a possible therapeutic target for anti-CSC therapy of OS. Furthermore, CSCs overexpressed the cell proliferation markers MKI67, CDC20, CDKN3, and CCNB1 and expressed more stemness markers than CSCL1/2/3 clusters (Fig. 6g). Differences in gene expression between the CSC and CSCL1/2/3 clusters are shown in Fig. 6i–l and Tables S16–19, demonstrating overexpression of genes related to the mitotic cell cycle in CSCs, such as TOP2A, CENPE, CENPF, and CCNB1. Pathway analysis also revealed DNA binding and chromosome condensation to be activated in the CSC cluster (Fig. S4a and Table S20). These results suggest that the CSC cluster might comprise a primary stem cell subset.
TF analysis was performed to detect specific TF activation in CSCs. It was found that TF was related to drug resistance, DNA methylation, and MSC differentiation, including EZH2, CTCF, PITX1 and HOXB7,28,29,30,31,32 which were obviously activated in CSCs compared with in CSCL1/2/3 clusters (Fig. 6m). We further detected activation and expression of EZH2, CTCF, PITX1, and HOXB7 in all mesenchymal clusters and found them to all be significantly overexpressed and activated in CSCs compared with in other mesenchymal clusters (Fig. S4b). These results describe the particular transcriptional program in CSCs.
Distribution change and molecular features of macrophages and osteoclasts in OSMacrophages and osteoclasts play crucial roles in tumor immunity and tumor-bone interactions in OS.33,34 Five classical clusters (monocytes, M1Φs, M2Φs, TAMs, and osteoclasts) were identified in this cell lineage (Fig. 7a). Each cluster was composed of cells from multiple samples (Fig. 7b). Monocle analysis revealed two differentiation trajectories of monocytes, differentiating into osteoclasts or macrophages (Fig. 7c), which is consistent with the known differentiation directions of monocytes.35 Monocytes, M2Φ, TAMs, and OCs were increased and M1Φ decreased in OS compared with CB tissues (Fig. 7d), possibly promoting the malignant behavior of OS, as M1Φs function as a tumor suppressor by activating the antitumor immune response.36 Each cluster was characterized by a distinct gene expression pattern with known markers, such as CD14 and FCGR3A in monocytes; IL1A and IL1B in M1Φs; CD163 and MRC1 in M2Φs; CD81 and CCL2 in TAMs; and TNFRSF11A, CTSK and ACP5 in OCs (Fig. 7e, f and Table S21).
Fig. 7
Activation of tumor-associated macrophages (TAMs) and osteoclasts (OCs) in OS. a UMAP plot of myeloid cells, including OCs, using the Monocle3 method. b UMAP plot of myeloid cells color-coded for each OS and CB sample. c Pseudotime trajectory of myeloid cells. d Differences in cell proportion between OS and CB samples in myeloid cell clusters. e Violin plots showing differences in expression of monocyte, M1Φ, M2Φ, TAM, and OC markers in each myeloid cluster. f Dot-plot heatmap of the most significant genes in the five myeloid clusters. g Heatmap showing differences in pathway activation between the five myeloid clusters calculated by the GSVA method. h Violin plots showing differences in activation of important pathways between the five myeloid clusters, including OC development, bone resorption, macrophage activation involved in immune response, regulation of antigen processing and presentation, tricarboxylic acid metabolic process, positive regulation of sprouting angiogenesis, positive regulation of fibroblast migration, and regulation of bone development pathways (part of the pathway is abbreviated in the figure)
GSVA was performed to detect the function of macrophages and osteoclasts. Consistent with the known roles of each cluster, pathways of bone resorption were activated in osteoclasts and pathways of the immune response in TAMs (Fig. 7g, h and Table S22). We also found that pathways related to the regulation of angiogenesis, fibroblast migration, and bone development were activated in osteoclasts (Fig. 7g, h), suggesting that osteoclasts may play a role in promoting angiogenesis and tumor progression in OS.
Perturbation of lymphocytes and endovascular cells in OSEight clusters were identified in T and B lymphocytes, including T-reg, CD4+ T, CD8+ T, NKT, DC, naïve B, active B, and plasma cells. Each cluster comprised of cells from multiple samples (Fig. S5a). The clusters were characterized by distinct gene expression (Fig. S5c and Tables S23 and 24), and the reported markers of lymphocytic clusters were verified to be expressed in specific subgroups (Fig. S5e). Nearly all T- and B-cell clusters were decreased in OS compared with in CB tissues (Fig. S5d), suggesting possible immunosuppression in OS.
Two clusters (endothelial and mural cells) were identified in endovascular cells (Fig. S5b), with both being supported by known and novel markers (Fig. S5c, e and Table S25). Possibly due to a sufficient blood supply of the tumor, both clusters of endovascular cells were significantly increased in OS compared with CB tissues (Fig. S5d).
Ligand‒receptor mediated intercellular interactions in the OS microenvironmentCellPhoneDB analysis was performed to detect interactions between clusters. Overall, intercellular interactions in OS samples were activated compared with those in CB samples; endothelial, osteoclast, and TAM cells exhibited relatively abundant interactions with OS cells (Fig. S6a). Part of the ligand‒receptor interactions between the clusters in OS and CB tissues are depicted in Fig. 8a, b. It was found that the tumor cells in OS samples exhibited a stronger ability to regulate angiogenesis, macrophage activation, and bone resorption than the osteoblasts in CB samples. For example, OS cells secreted VEGFA, GRN, and TNFSF11, which bind to KDR on endothelial cells, TNFRSF1A on TAMs, and TNFRSF11A on osteoclasts (Fig. 8c). However, lymphocytes were more activated in CB samples, such as the CXCL12-CXCR4 interaction between monocytes and lymphocytic subsets in CB samples (Fig. 8c), suggesting immunosuppression in OS.
Fig. 8
Ligand-receptor interactions between clusters in OS and CB samples. a Dot-plot heatmap shows part of the ligand-receptor interactions between the clusters calculated by the CellPhoneDB method. b Dot-plot heatmap showing part of the ligand-receptor interactions between clusters in CB samples. c Schematic representation of differences in the microenvironment between OS and CB samples. d Schematic representation of differences in intercellular interactions between OS-A2/B2/C2 and other cell clusters in OS samples
In addition, communications between OS-A2/B2/C2 and other subsets were obviously different. For example, OS-A2 and OS-B2 expressed more VEGFA/B, which activate their receptors on endothelial cells and might promote angiogenesis. OS-B2 expressed CXCL12 and might activate TAMs and CD8 T cells by binding to CXCR4. OS-C2 markedly expressed TNFSF11 and CSF1, which promotes osteoclast formation through activated TNFRSF11A and CSF1R. Self-interactions of TIMP1-FGFR2 and FGF7-FGFR1 were also observed in OS-B2 and OS-C2, respectively (Fig. 8a, d). The ligand‒receptor interactions between OS-A2/B2/C2 and endothelial cells, TAMs, and osteoclasts are shown in Fig. S6b. These results suggest that the tumor microenvironment (TME) is different between the three subgroups of OS samples, which might provide clues for therapy for the three subgroups.
留言 (0)