
記住我


We hypothesize oxidative stress, specifically mitochondrial ROS, is a primary mechanism through which IO causes cell death in H9c2 cells. We show that MitoNEET overexpression in H9c2 protects against IO-induced cell death by reducing iron content in the mitochondria and preventing mitochondrial ROS.
2. Materials and Methods 2.1. ReagentsFeSO4 (Ferrous Sulfate Heptahydrate, 215422), N-acetyl-L-cysteine (NAC, A7250) and DCF-DA (2′,7′-dicholorodihydrofluorescein diacetate dye, D6883), Ampicillin (A9518) LB broth (L3522), G418 (A1720) were from MilliporeSigma (Burlington, ON, Canada). Hoechst nuclear dye (H10325), CellROXTM Deep Red Reagent (C10422), Image-ITTM DEAD GreenTM Viability Stain (I10291), MitoSOXTM Red (M36008), MitoTrackerTM Deep Red FM (M22426) and CellEvent Caspase 3/7 Green Detection Reagent (C10423), Subcloning EfficiencyTM, ReadyProbesTM Cell Viability Imaging Kit (R37609), DH5α Competent Cells (18265017), Pierce Protease and Phosphatase Inhibitor Mini Tablets (A32959) were from Thermo Fisher (Waltham, MA, USA). Gibco brand Dulbecco’s Modified Eagle’s Medium (DMEM), fetal bovine serum (FBS) and 1% penicillin/ streptomycin were from Thermo Fisher (Waltham, MA, USA). 10-(6’-plastoquinonyl decyltriphenylphosphonium (SkQ1, HY-100474) was from MedChemExpress (Monmouth Junction, NJ, USA). FerroOrange was from Dojindo Molecular Technologies (Rockville, MD, USA). Anti-TOM20 antibody (sc-17764) and anti-Cytochrome c antibody (sc-25304) were purchased from Santa Cruz Biotechnology (Dallas, TX, USA). The CytoscanTM-LDH Cytotoxicity Assay kit (786-210) was purchased from G-Biosciences (St. Louis, MO, USA). PBS containing calcium and magnesium was from Wisent Bioproducts (311-011-CL, Wisent Inc., St-Bruno, QC, Canada). BSA (ALB001, Bioshop, Burlington, ON, Canada).
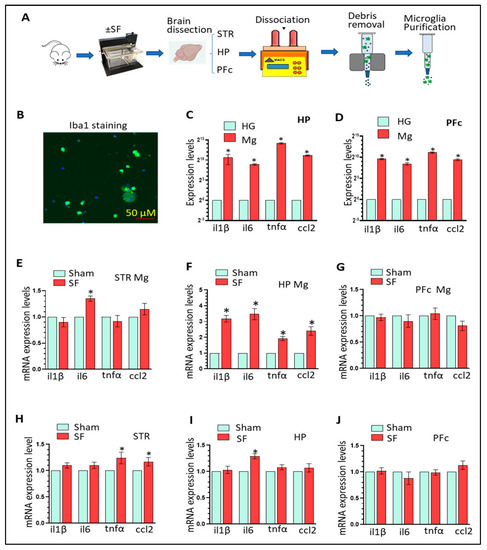
2.2. Cell CultureH9c2 rat embryonic cardiomyoblasts (ATCC, Manassas, VA, USA) were grown in DMEM supplemented with 10% FBS and 1% penicillin/ streptomycin (P/S) at 37 °C and a 5% CO2 atmosphere. The H9c2 cell line is a subclone from an original clonal cell line derived from embryonic BD1X rat heart tissue by Kimes and Brandt [16] and resemble immature embryonic cardiomyocytes [17]. Iron overload was induced by incubation of H9c2 in DMEM with 0.5% FBS and 1% P/S, supplemented with 100 µM FeSO4 for 24 h when cells reached approximately 70% of confluency. This concentration of FeSO4 is within the range typically used by researchers in the field to modulate the activity of iron regulatory proteins (IRP1 and IRP2) and trigger homeostatic processes to iron overload. NAC (500 nM) and SkQ1 (20 nM), a general antioxidant or mitochondrial specific antioxidant, respectively, were supplemented where indicated, 30 min prior to iron treatment and for the remainder of the 24 h. For assays requiring microscopy, cells were cultured on cover slips or imaging chambers at approximately 70% confluency for durations as specified in each assay. 2.3. Creation of Stable MitoNEET-Overexpressing Cell LineEscherichia coli DH5α were transformed using heat shock with plasmids containing either the MitoNEET (pMitoN) gene or empty vector (EV) (Genescript, clone ID Ora10938 and SC1217, respectively). Cultured and transformed E. coli colonies were selected on an LB/ampicillin plate and further sub-cultured in LB/ampicillin media. pMitoN and EV plasmids were purified using the QIAGEN Plasmid Midi Kit and confirmed with visualization with ethidium bromide by agarose gel electrophoresis relative to a DNA molecular weight ladder (Abcam, Cambridge, UK). H9c2 cells at 70% confluency were then transfected with pMitoN and EV plasmids using the Lipofectamine 3000 transfection kit (Thermo Fisher, Waltham, MA, USA) for 48 h. Afterwards, transfected H9c2 cells were selected using 10% FBS DMEM containing 500 µg/mL G418 antibiotic for 5 days. Surviving cells were collected and sub-cultured in a 96-well plate at a low density in G418-containing DMEM + 10% FBS for 2 weeks. The 5 most confluent wells from the pMitoN plate were collected and characterized, whereas colonies from the EV plate were pooled and cultured. Following characterization, the colony lineage expressing the most pMitoN was selected for further expansion and use in this study. EV-transfected H9c2 cells were used as control.
2.4. Intracellular Iron QuantificationFerroOrange is a cell-permeant non-fluorescent dye in its unbound state but exhibits fluorogenic signal when bound to ferrous (Fe2+) ions. For live detection of intracellular iron, H9c2 were seeded at 70% confluence in DMEM + 0.5% FBS on µ-slide 8-chambered polymer coverslips (Ibidi, GmbH, Germany) and treated with or without FeSO4 (100 µM, for the indicated times) in the presence of FerroOrange (1 µM) and nuclei-staining dye Hoechst (100 nM) at 37 °C, 5% CO2. The cells were imaged every 5 min on a Nikon A1 Confocal Microscope fitted with an incubated chamber (37 °C, 5% CO2). Images were analyzed using NIS-Elements (Nikon).
2.5. Mitochondrial Iron QuantificationMitoFerroFluor, MFF (a kind gift from Dr John J. Lemasters), is a new fluorescent marker used for measuring mitochondrial iron [18]. MFF accumulates in the mitochondria where it exhibits a fluorescent signal that can be quenched by ferrous iron. For experimentation, H9c2 cells were seeded at 70% confluence in DMEM + 0.5% FBS on µ-slide 4-chambered polymer coverslips and treated with FeSO4 (100 µM, 24 h). At treatment endpoint, cells were washed with PBS three times, then incubated with MFF (2 µM) and Hoechst (100 nM) for 30 min in PBS at 37 °C protected from light. Cells were washed with PBS three times and imaged using Nikon A1 Confocal Microscope fitted with an incubated chamber (37 °C, 5% CO2). Mean MFF fluorescent intensity was quantified by ImageJ. 2.6. Reactive Oxygen Species DetectionCellROX Deep Red oxidative stress reagent is not fluorescent in a reduced state but exhibits a strong fluorescent signal when oxidized. For live detection of ROS production, H9c2 cells were seeded at 70% confluency in DMEM + 0.5% FBS on µ-slide 4-chambered polymer coverslips (Ibidi, GmbH, Germany), and pre-stained 15 min with Hoechst (100 nM) in DMEM + 0.5% FBS. At the start of imaging, CellROX Deep Red (2.5 µM) was added, alongside FeSO4 (100 μM), for the indicated times. Imaging was performed with 20× objective (Nikon A1 confocal microscope) at 5 min intervals in a 37 °C, 5% CO2 live-cell chamber. For fixed detection of ROS production, H9c2 cells were seeded onto glass coverslips and treated with FeSO4 (100 μM, 4 h). CellROX® Deep Red (2.5 µM) was added to each well for 15 min. Cells were washed briefly with PBS and fixed with 10% formalin, before being mounted onto to a glass slide with VECTASHIELD mounting medium plus DAPI. Images were taken at 40× objective (Nikon A1). Images were analyzed using NIS-Elements (Nikon). When using DCF-DA, cells were washed with warmed PBS following FeSO4 treatment (100 μM, 4 h), and incubated with DCF-DA (20 µM) in PBS for 30 min at 37 °C protected from light. Afterwards, the cells were washed 3 times with warmed PBS. Fluorescence from DCF-DA was measured at 490 nm excitation and 520 nm emission using a Varioskan LUX plate reader (Thermo Fisher, Waltham, MA, USA). The antioxidants NAC and SkQ1 were used, as indicated.
2.7. Mitochondrial Reactive Oxygen Species DetectionMitoSOX Red is selectively targeted to mitochondria and exhibits a strong fluorescent signal upon oxidation. For live detection of mitochondrial ROS production, H9c2 cells were seeded at 70% confluency in DMEM + 0.5% FBS on a µ-slide 4-chambered polymer coverslip and treated with FeSO4 (100 μM, 4 h). At treatment endpoint, cells were washed with PBS three times, then incubated with MitoSOX Red (5 µM) and Hoechst (100 nM) for 30 min at 37 °C protected from light. Cells were washed times with PBS then imaged using a Nikon A1 confocal microscope, fitted with an incubated chamber (37 °C, 5% CO2). Mean MitoSOX intensity was quantified by ImageJ.
2.8. Caspase 3/7 Detection KitCellEvent Caspase 3/7 Detection Reagent is cell-permeant reagent and consists of the peptide (DEVD) conjugated to a nucleic acid-binding dye. Upon cleavage of the DEVD recognition sequence by caspases 3/7, the dye is released to bind cellular DNA increasing its fluorescent signal. For detection, H9c2 cells were plated at 70% confluency in 96-well plates (non-tissue culture coated) and treated with FeSO4 (100 µM, 24 h) with or without co-treatment with antioxidants NAC (500 nM) and SkQ1 (20 nM). At the treatment endpoint, cells were incubated with Caspase 3/7 Detection Reagent (10 µM) and Hoechst (100 nM) for 30 min at 37 °C protected from light. Afterward, images were obtained and analyzed to characterize cells with activated Caspase 3/7 pathways through the CellInsight CX7 High Content Screening platform (Thermo Fisher, Waltham, MA, USA), Absorption/Emission: 511/533 nm.
2.9. Flow CytometryAlexa Fluor® 488 Annexin V/Dead Cell Apoptosis Kit (V13241, Thermo Fisher, Waltham, MA, USA) was used to detect the early and late stages of apoptosis using annexin V and propidium iodide (PI). Annexin V detects phosphatidyl serine on the surface of the cell membrane surface during early stages of apoptosis. PI is a membrane impermeable nucleic acid binding dye used to detect late apoptotic cells, as per manufacturer’s instructions. H9c2 cells were plated on 6-well plates and cultured to reach 70–80% confluency. Treatment conditions for cells are outlined in the figure legends-conditions included 0.5% FBS media, FeSO4 100 µM, and co-treatment with NAC 500 nM or 20 nM SkQ1. Following treatments, cells were lifted off the culture dishes with trypsin-EDTA in PBS and immediately incubated with annexin V and PI diluted in 1x annexin-binding buffer (30 min at 4 °C), then mixed with additional 1x annexin-binding buffer. Cells were placed on ice until immediate flow cytometry analysis using the AttuneTM NxT Acoustic Focusing cytometer (Thermo Fisher, Waltham, MA, USA) with the BL1 and BL3 lasers for Annexin V and PI, respectively.
Cells were stained with MitoSOX Red assay and monobromobimane (mBBr) at treatment endpoints as per the manufacturer’s protocol. Following staining, cells were washed and lifted off the culture dishes with trypsin-EDTA in PBS and suspended in 2% FACS buffer on ice until immediate flow cytometry analysis on the AttuneTM NxT Acoustic Focusing cytometer (Invitrogen, Waltham, MA). The VL2 and BL1 lasers were used for mBBr and MitoSOX Red, respectively. Flow cytometry data were analyzed using FlowJo® software (Tree Star Inc., Ashland, OR, USA).
2.10. ImmunocytochemistryH9c2 cells were grown on glass coverslips in 24-well plates. They were allowed to reach 70% confluency and treated with FeSO4 (100 µM, 24 h) in DMEM + 0.5% FBS without or without the co-treatment of antioxidants 500 nM NAC and 20 nM SkQ1. At the treatment endpoint, the coverslips were washed briefly with PBS (PBS with added Mg2+ and Ca2+) and fixed with 10% formalin solution for 30 min. Following 3 more washes, the cells were permeabilized using PBS containing 0.5% Triton-X detergent for 30 min and blocked with PBS containing 3% BSA for 1 h. Afterwards, the cells were incubated in 3% BSA in PBS solution containing mouse anti-TOM20 (Santa Cruz, for mitochondria detection) and rabbit anti-Cytochrome c (Abcam, Cambridge, UK) at 1:500 dilution overnight at 4 °C. Cells were then washed 3 times with PBS containing 0.5% Tween-20 and incubated in 3% BSA in PBS solution containing anti-rabbit IgG Alexa Fluor 555 and anti-mouse IgG Alexa Fluor 488 (Thermo Fisher, Waltham, MA, USA) at 1:500 dilution for 1 h at room temperature. Afterwards, cells were washed 3 times with PBS containing 0.5% Tween-20. Coverslips were mounted onto slides in mounting medium containing a 1:1 mixture of VECTASHIELD anti-fade mounting medium with DAPI and imaged using the Nikon A1 Confocal Microscope. Images were analyzed using NIS-Elements (Nikon). For 3D images, z-stacks were captured, then rendered on IMARIS 9.9.0 using the surfaces tool.
2.11. Cell Death AssaysCell rupture was quantitatively evaluated by measuring the released lactate dehydrogenase (LDH) with CytoScanTM LDH Cytotoxicity Assay (G-Biosciences, 786-210). H9c2 cells were treated with FeSO4 (100 μM, 24 h) in DMEM + 0.5% FBS with or without antioxidants NAC (500 nM) and SkQ1 (20 nM). The cultured media were mixed in 1:1 ratio with the mixed substrate solution followed by incubation for 30 min in the cell culture incubator (37 °C, 5% CO2) protected from light. The assay was quantified by absorbance at 490 nm subtracted by 680 nm and plotted as a relative value over the control condition.
Image-IT™ DEAD Green™ Viability Stain was used to detect dead cells with compromised plasma membrane. H9c2 cells were plated at 70% confluency in DMEM + 0.5% FBS in 96-well plates (non-tissue culture coated) and treated with FeSO4 (100 μM, 24 h) with or without co-treatment with antioxidants NAC (500 nM) and SkQ1 (20 nM) for. At treatment endpoint, the Viability Stain (100 nM) and Hoechst (100 nM) were added to the cell media for 30 min (37 °C, 5% CO2) protected from light. Afterwards, treatment media were removed, and cells were washed briefly with PBS and fixed with 10% formalin solution. The images were obtained and analyzed to characterize dead compromised cells through the CellInsight CX7 High Content Screening platform in the FITC (green) channel.
ReadyProbes™ Cell Viability Imaging kit was used to determine viability of cells. H9c2 cells were seeded at 70% confluence in 24-well plates and treated with FeSO4 (100 μM, 24 h) in DMEM + 0.5% FBS. At treatment endpoint, cells were stained according to manufacturer’s protocol, then imaged using EVOS FL Auto 2 Imaging system (Thermo Fisher, Waltham, MA, USA).
2.12. Quantitative Polymerase Chain Reaction (qPCR)H9c2 cells were plated on a 6-well plate. When the confluency reached to 70%, cells were treated with FeSO4 (100 μM, 24 h) in DMEM + 0.5% FBS. At the treatment endpoint, cells were trypsinized and washed 3 times with PBS at 4 °C. Cells were then lysed and processed to collect RNA by using the RNeasy Mini Kit (QIAGEN). The extracted RNA was quantified using the plate reader-based µDropTM Plate (Thermo Fisher, Waltham, MA, USA) and approximate 400 ng of total RNA per sample was used in reverse transcription using the RevertAid RT Reverse Transcription Kit (Thermo Fisher) using random hexamer primer to obtain total cDNA. The qPCR solution was prepared with cDNA, the iQTM SYBR® Green Supermix (Bio Rad, Hercules, CA, USA), and the respective primers (outlined in Table 1) and the reaction was performed using a Bio Rad qPCR machine. The samples were placed at 95 °C for enzyme activation, 95 °C for denaturation, 60 °C for annealing, 72 °C for extension, followed by a plate read. Data was quantified and analyzed using the delta-delta Ct method normalized over the 18S rRNA. 2.13. Western BlotFollowing treatment endpoint (100 μM FeSO4 in DMEM + 0.5% FBS for 24 h), H9c2 cells were lysed in RIPA lysis buffer containing a protease and phosphatase inhibitors. After denaturing lysates in Laemmli sample buffer for 10 min at 90 °C, samples were resolved on an SDS-PAGE gel and transferred onto PVDF membrane. The membranes were blocked with 3% BSA for 1 h at room temperature. Membranes were then incubated overnight at 4 °C in primary antibodies against: Cleaved caspase-3 (Asp175, 1:1000, Cell signaling, #9661), CISD1/MitoNEET (D5M4C, 1:1000, Cell signaling technology (CST), Danvers, MA, USA, #83775), GAPDH (14C10, 1:1000, CST, #2118). Membranes were then washed and incubated in the corresponding horse-radish peroxidase (HRP)-linked secondary antibody for 1 h at room temperature: anti-mouse IgG, HRP-linked antibody (1:10,000, CST, #7076) and anti-rabbit IgG, HRP-linked antibody (1:10,000, CST, #7074). The antibody-antigen complex was reacted with enhanced chemiluminescence (Bio Rad, Hercules, CA, USA) reagents before developing and the signal was detected by using CL XPosure Film (Thermo Fisher, Waltham, MA, USA). The density of each band was quantified using Image J, NIH Image.
2.14. StatisticsData are given as mean ± SEM, and one-way analysis of variance was used for comparison in GraphPad Prism 8.0. All experiments were performed with a minimum of 3 biological replicates (n = 3), unless otherwise stated. Differences were significant at * p < 0.05, ** p < 0.01, *** p < 0.001 vs. Control. # p < 0.05, ## p < 0.01 vs. FeSO4.
4. DiscussionCardiomyopathy and heart failure are common pathologic endpoints for many patients with metabolic syndrome [20]. Such outcomes have been thought to have multi-factorial causes, including hypertension, insulin resistance, inflammation, and cardiomyocyte death [21]. IO-related cardiomyopathy and heart failure have been a primary focus of studies examining genetic traits that either perturb iron metabolism or medical conditions that require therapeutic transfusions [1,2]. In the last decade, however, there has been increasing interest in the intersection between IO pathologies and the metabolic syndrome, since serum ferritin levels (which can be used as a marker of IO) have been strongly associated with the severity of insulin resistance and metabolic syndrome [19,20] —sparking a re-emergence of the terminology “dysmetabolic iron overload syndrome (DIOS)” [22,23]. Recent evidence pinpoints the heart as a key organ affected in these concurrent diseases due to heightened oxidative stress and insulin resistance [24].In our current study, we examined cell death by IO-induced oxidative stress in H9c2 cells cultured with FeSO4 for up to 24 h to simulate an acute model of IO. Using FerroOrange staining we observed significant intracellular iron accumulation within 2 h of treatment time, that was paralleled by significantly increased cellular ROS levels within the same time frame (tracked by CellROX or DCF-DA) and this could be moderated by the general antioxidant NAC. Longer incubation times with FeSO4 upregulated expression of the cytoplasmic and mitochondrial isoforms of the iron-storing proteins, Ferritin. Heightened ROS was shown to trigger the apoptosis program (Cleaved-caspase 3/7, Cytochrome C loss from mitochondria, increased PM phosphatidylserine detection with Annexin V) and increased the incidence of lytic cell death (e.g., LDH release) within the 24 h time-frame studied. Having observed that iron accumulation upregulated mitochondrial Ferritin, and the MFF detection of iron elevation within the mitochondria, the role of mitochondria-specific ROS generation was examined. Using MitoSOX, we observed increased production of mitochondrial ROS in IO, which was prevented by treatment with the mitochondrial antioxidant, SkQ1. IO-induced mitochondrial ROS induced apoptosis and cell death which were also reduced or prevented by SkQ1, thus implicating mitochondrial ROS as a causative factor.
MitoNEET is a recently discovered mitochondrial protein [25,26,27], originally characterized by its interaction with pioglitazone [28]. While there has been ample literature studying MitoNEET’s function, related to its binding of iron–sulfur complexes and its similarities to other (NEET) proteins containing CDGSH domains [27], studies investigating its roles in biological systems, especially those employing gain- or loss-of-function genetic alterations, are only just emerging [12,29]. It was from these studies that MitoNEET was found to regulate the mitochondrial respiration rate when overexpressed, likely by reducing the iron levels in the mitochondrial matrix [12]. Based on this evidence, we created a H9c2 cell line, stably overexpressing MitoNEET to examine how it may partition intracellular iron content. Under IO treatment, we found that these cells accumulated less iron in their mitochondria using the mitochondrial iron-binding fluorescent dye, MFF [18]. This observation correlated with less cellular and mitochondrial ROS overproduction and reduced biomarkers of apoptosis-related cell death and reduced cell lysis. These results established the potential of MitoNEET as a safeguard against IO at the cellular and mitochondrial levels.IO-induced oxidative stress is well characterized as an important contributor to the pathogenesis of IO cardiomyopathy, myocardial injuries and heart failure [30,31]. From our current model, events underpinning IO cardiomyopathy could be attributed to IO within the mitochondria. By inhibiting the effects of mitochondrial iron accumulation (increased mitochondrial ROS) with the SkQ1 antioxidant or iron accumulation within this organelle, with MitoNEET expression, the deleterious effects of iron overload could be mitigated.Regulation of mitochondrial iron by MitoNEET’s has not been fully elucidated. The transfer of its iron–sulfur clusters to acceptor proteins can be dependent on its own oxidative status. Specifically, in an oxidized form, this transfer occurs rapidly while it is inhibited when MitoNEET is reduced [32]. MitoNEET itself is reducible by biological antioxidants such as glutathione [27,33]. MitoNEET also exhibits lower iron–sulfur transfer kinetics when interacting with the anti-diabetic drugs in the thiazolidinedione class, such as pioglitazone, as these compounds stabilize MitoNEET structure and its binding affinity to the iron–sulfur clusters [32]. In a separate study of a spinal cord injury model, this interaction was shown to be essential in pioglitazone’s therapeutic effects [33]. Given that pioglitazone is insulin-sensitizing and iron overload has been shown to result in insulin resistance in the heart [24], pioglitazone’s interaction with MitoNEET and potential involvement in mitochondrial iron regulation may be a basis of part of its therapeutic impact.Our study is limited by utilizing an in vitro model of iron overload in H9c2 cells to examine its consequences on cell viability. However, with a rising prevalence of metabolic syndrome and diabetes worldwide, secondary IO and the associated DIOS could be significant risk factors of cardiomyopathy, heart failure, and death [34]. Thus, our study points to prevention or therapeutic intervention against IO may prove advantageous in treating these maladies. Our study showed the beneficial role played by MitoNEET in counteracting IO and its consequences at the cellular level. While MitoNEET overexpression may not be clinically feasible, therapies utilizing antioxidants and thiazolidinediones may enhance MitoNEET activity and is worthy of further investigation.
留言 (0)