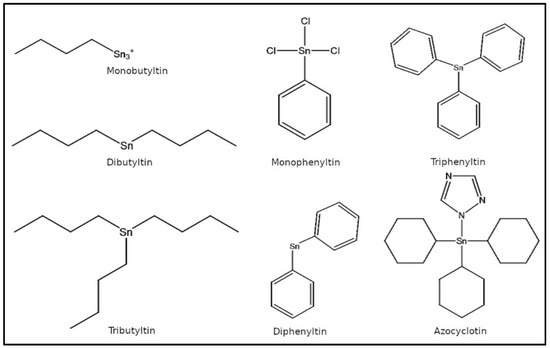
1. IntroductionOrganotin compounds (OTCs), also called as stannanes, are a large class of organometallic compounds having at least one tin atom covalently bound to a carbon atom with a general formula ‘RSnX’ where ‘R’ represents an organic group, i.e., alkyl, phenyl, etc., and ‘X’ an anion such as chloride, fluoride, oxide, etc. [
1,
2,
3,
4,
5]. Tin metal and its alloys have been used by humans for more than 6000 years, however, the first OTC was synthesized only about 175 years back in the year 1853, and, for nearly another 100 years, OTCs were not found to have any industrial utility for want of commercial applications [
5,
6]. As of now, there are more than 800 OTCs known, and, barring a few, all of them are anthropogenic [
7]. OTCs constitute one the largest group of organometallic compounds in commercial use globally, and the United States and China are the largest consumers of OTCs, followed by Western Europe, the Middle East, and others [
8]. The downside of the increased industrial applications and use has been that considerable amounts of OTCs have entered various ecosystems, and OTCs are now ubiquitous in the environment. The worldwide higher use and demand have resulted in an increase in global production of OTCs during the last several decades. Overall, the maximum utilization of OTCs is as polyvinyl chloride (PVC) stabilizers (about 70–80% of global consumption) and remaining as biocides [
5,
6,
9].OTCs are classified according to the number of organic functional groups on the tin atom or the number of ‘carbon-tin’ bonds, e.g., mono-, di-, tri-, and tetra-organotin compounds [
5]. Generally, mono- and di-substituted OTCs, e.g., dibutyltin (DBT), diphenyltin (DPT), etc., are used as heat stabilizers for PVC plastics, lubricating oils, hydrogen peroxide, and polyolefins and as catalysts in the production of polyurethane foam, polymers, esters, plastisol prints, rubber, adhesives, etc., with applications in apparel and footwear industries [
2,
5]. Tri-substituted OTCs, e.g., tributyltin (TBT), triphenyltin (TPT), etc., have been used extensively as biocides, such as antifouling paints for boats, timber preservatives, preservatives in textiles, leathers and synthetic leathers, fungicides in crops including potatoes, sugar beets, and pecans, and as pesticides [
2,
5,
10]. OTCs are also utilized in silicone-based finishes having elastomeric and water repellency properties. The seven most common OTCs commercially available for a variety of industrial applications are TBT, DBT, monobutyltin (MBT), TPT, DPT, monophenyltin (MPT), and azocyclotin (ACT).Environmental contamination with OTCs has been reported to be mainly through agricultural runoffs and marine aquatic environment due to leaching from antifouling paints, treated timber, PVC pipes, and other contaminated material [
4,
5,
10]. Biodegradation half-lives of organotins are generally shorter in tri-substituted organotins compared to di-substituted organotins, whereas monosubstituted organotins have the longest half-lives. The half-lives are longer in both seawater and soil/sediment than in fresh water and can range from six months to 15 years [
5]. The ‘C-Sn’ bond of organotins is comparatively stable, as the anionic group is reported to dissociate easily on hydrolysis in water. Human exposure occurs through drinking water and eating food, especially aquatic food such as fish and other marine animals. The global terrestrial and aquatic contamination and the adverse effects on non-target organisms including abnormalities in shell calcification in oysters and the masculinization of female gastropods—imposex has resulted in outlawing the use of OTs, especially tributyltin in marine paints in 2008 under the International Convention on the Control of Harmful Antifouling Systems on Ships and World Health Organization [
5,
10,
11]. However, developing nations that are not members of the International Maritime Organization are still permitted to use OTCs for industrial and agricultural purposes. Organotins have been found in household dust in many countries, including the United States and Germany [
12,
13]. Due to an increase in the usage of antifouling paints containing organotin compounds, organotin contaminations have also been observed in Saudi Arabia in recent years in fishing harbors of various coastal sites in the Eastern Province (Jubail, Khobar, and Qatif) [
14,
15]; the estimations of the contamination have shown more phenyltins than other organotins. The detection of high concentrations of hazardous triorganotins in the commercially important fish species caught from Arabian Gulf is of particular concern for human health in Saudi Arabia [
16].Limited epidemiological studies have been reported on the exposure of humans to organotin toxicity. In the United States, MBT, DBT, and TBT were detected in 70% of the human blood samples that were tested [
17]. The potential toxicological significance of OTCs to humans on the basis of animal experiments indicates diverse health problems including immunosuppression, endocrine problems, neurotoxicity, metabolic and enzymatic problems, ocular, cardiovascular, gastrointestinal, reproductive, developmental, and several other problems [
18]. OTCs cause morphological and functional changes in a number of tissues in animals that are involved in the regulation of endocrine function and metabolism in mammals, including the hypothalamus, pituitary, pancreas, gonads, adipose tissue, adrenal, and thyroid glands [
19,
20]. In regard to reproductive problems in humans, organotin exposure has been associated with congenital abnormalities in neonates such as cryptorchidism and hormone disbalance for LH and testosterone in newborn boys [
21]. Organotins have been also reported to cause metabolic dysfunction in animal models similar to polycystic ovarian syndrome (PCOS) in women [
22]. In animals, organotin exposure leads to phenotypic abnormalities, aberrant changes in hormones, transcriptome and proteome modifications, behavioral changes, and characteristic sexual organ masculinization called imposex, in which female gastropods develop penile structure and vas deferens [
23,
24]. In rats, organotins are associated with reduced sperm viability and motility, abnormalities of spermatids and spermatozoa, testicular necrosis, changes in the blood–testicular barrier, and hormone disbalance in males [
24,
25,
26], as well as delayed follicular development, irregular estrous cycles, alterations in steroidogenic enzymes, abnormal adipogenesis, decline in fertility, failure of implantation, lower chances of conception, and other reproductive abnormalities in females [
27,
28,
29].The available literature indicates that very few epidemiological or clinical studies have been reported on the effects of organotins on human health despite the overwhelming evidence of the adverse effects on laboratory and marine species. Computational methods have been increasingly used for the prediction of binding pose and molecular interactions of ligands with their target protein molecules for the characterization of interactions, designing novel inhibitors, and/or as an aid for designing experimental and clinical studies [
30,
31,
32]. The present study was performed to characterize the structural binding interactions of seven commonly available OTCs, viz., TBT, DBT, MBT, TPT, DPT, MPT, and ACT, against sex-steroid nuclear receptors, i.e., androgen receptor (AR) and estrogen receptors (ERα, ERβ). AR and ER are soluble proteins and function as intracellular transcription factors. Testosterone and dihydrotestosterone are the main steroid ligands of AR. In addition to the development of male sexual organs, maturation of male sexual organs and spermatogenesis, AR signaling plays a crucial role in a number of physiological and developmental processes related to male physiology. ER regulates the transcription of numerous genes, and its primary role is in the development of female phenotype, development of female reproductive tract, regulation of folliculogenesis, uterine development, and in other processes related to reproductive cycle. The suggested hypothesis is that OTCs may interfere with the natural interaction between ligands and proteins by binding to sex steroid nuclear receptors in the body, which might result in dysfunction of sex steroids target organs. 4. Discussion
The objective of this study was to characterize the structural binding interactions of seven commonly available OTCs, viz., TBT, DBT, MBT, TPT, DPT, MPT, and ACT, against sex-steroid nuclear receptors, i.e., AR, ERα, and ERβ. As already indicated, docking analysis showed that four of seven organotin ligands, i.e., TBT, DBT, DPT, and MPT, bound well within the binding sites of the three receptors. The remaining three organotin ligands, MBT, TPT, and ACT, showed a weak binding or did not bind with the receptors and, hence, further analysis of these ligands for molecular interactions and binding pose analysis was not considered. The dock score, binding energy, and dissociation constants for the four organotins with AR, ERα, and ERβ indicated good docking and binding within the ligand binding sites of the receptors. Similar binding poses of the organotin ligands and the respective bound native ligands along with close similarity in binding energies indicated good binding and thus provided credence to the docking accuracy. In addition, high overlapping/commonality of interacting residues of each receptor for the docked ligands and respective native ligands provided further support for good docking. This was especially true for the interacting residues of ERα, and ERβ for the organotin ligands and native ligands as ERα, and ERβ showed better structural binding characteristics compared to AR. The good docking and stability of docking complex was further supported by the results of MD simulation of a representative organotin/receptor complex (TBT/AR), with a RMSD plot showing a stable interaction of TBT with AR with low deviation of ligand within good acceptable range. In addition, RMSF and Rg plots showed maintenance of compactness of the complex along with no change in the SASA profile during the whole simulation period. Thus, the MD simulation also provided support and indicated good stability of the receptor–ligand complex. Taken together, all the above results suggested that the organotin ligands are bound tightly in the same ligand binding pockets of AR, ERα, and ERβ as their respective bound native ligands, testosterone and estradiol, especially for the ERα, and ERβ. Thus, the suggested hypothesis that OTCs may interfere with the natural interaction between sex steroids and their receptors by binding to sex steroid nuclear receptors was supported. Hence, the indicated organotins have the potential to interfere with the binding of testosterone and estradiol17β to their respective receptors and result in dysfunction of steroid receptor signaling.
Previous reports on the in silico studies of organotins with sex steroid nuclear receptors are not available to the best of our knowledge. However, organotins, TBT and TPT, were shown to have competitive binding antagonistic activity against human ER in vitro [
42]. The inhibitory effect on ER similar to other ER antagonists, such as 4-hydroxytamoxifen, was shown on ER-dependent reporter gene transcriptional activation by TBT and TPT at very low concentrations by interactions between human ERβ LBD and the co-activator SRC1 in a yeast two-hybrid detection system. In addition, TBT and TPT stimulated LA16 cells (that stably expressed androgen-responsive luciferase reporter gene and proliferates in response to androgen) and enhanced both AR-dependent transcription of luciferase gene and cell growth similar to dihydrotestosterone [
43]. Simultaneous treatment of LA16 cells with dihydrotestosterone and TBT or TPT caused synergistic effects on AR activation, but an androgen antagonist, flutamide, did not inhibit the TBT- or TPT-induced AR activation, suggesting a novel mechanism other than the ligand-binding site of AR. TBT and TPT were also shown to interact with other nuclear receptors, e.g., as nanomolar agonists of retinoid X receptor (RXR) and peroxisome proliferator-activated receptor γ (PPARγ). In this regard, TBT and TPT exposure to a RXR-transfected human choriocarcinoma cell line (JEG-3 cells) stimulated luciferase expression, indicating activation of RXR [
44]. In vivo and in vitro studies on organotins in marine invertebrates, fish, mammals, and laboratory animals have shown the profound endocrine disruption effects of organotins on reproductive and other functions. Development of imposex in mollusks even with exposure to very low TBT concentrations is the most sensitive and well known reproductive phenotypic effect of endocrine disruption [
6]. The endocrine disruption by organotins in various species is believed to occur through three main mechanisms, i.e., (1) increased testosterone—organotins cause an androgen abundance by inhibiting the aromatase in females [
45], (2) the APGWamide neuropeptide activation—organotins induce abnormally high levels of peptide APGWamide, which cause the development of male-like tissues subsequently producing androgens to promote the male-like sexual growth, and (3) the RXR agonism/activation—organotins cause abnormal activation of the RXR signaling pathway through ligand binding or by increasing the retinoid [
23]. RXRs in association with other nuclear receptors, such as PPAR, etc., regulate cellular development and differentiation, metabolism, and cell death. In addition, recently inactivation of UDP-glucuronosyltransferases (UGTs) was also proposed to be another mechanism by which organotins can cause abnormal reproductive health effects; UGTs regulate metabolic inactivation of many endogenous hormones [
46]. Inhibition of UGTs, including UGT2B15 (regulates catalysis of dihydrotestosterone glucuronidation) and UGT1A1 (regulates catalysis of estradiol-3-O-glucuronidation) by organotins such as TBT and TPT, may interfere with glucuronidation of endogenous sex hormones and thus inhibit their termination, resulting in abnormal endocrine actions [
46]. The endocrine disruption mechanism of organotins in humans is not clear but has been postulated to most likely involve increased testosterone [
4]. In addition, the potential for TBT and other organotins to act as obesogens in humans and to interfere with endocrine regulation of adipogenesis such as stimulation of preadipocyte differentiation into adipocytes in a PPARγ-dependent manner is also very concerning.Very limited epidemiological studies have been reported on the association of organotin exposure and human reproductive problems. Several reports have mentioned general toxic effects associated with acute human organotin exposure, such as seizures, visual disturbances, paraparesis, forgetfulness, fatigue, weakness, loss of motivation, depression, and attacks of rage; some symptoms persist for at least three years [
2,
47,
48]. Information associated with long term exposure is not reported. In other studies, severe toxicity of TPT showed neurotoxic symptoms, such as cerebellar syndrome, hearing impairment, and loss of consciousness with paroxysmal activity on electroencephalography [
49]. In general population, significant levels of the organotins TBT and TPT have been detected in the blood of human male and female volunteers [
50]. In another study, triorganotins, such as TBT and TPT, were detected in 37% and 99% of the placental tissues of women in Denmark and Finland, respectively, and a positive correlation between organotin (DBTCl) levels in the human placenta and incidence of cryptorchidism was reported in newborns in Denmark [
21]. In addition, in the same study, the blood LH levels in four-month-old boys were negatively correlated, whereas the inhibin B levels were positively correlated with TBTCl in the placenta of women from Finland.Several studies have reported adverse effects of organotins in male reproductive function in laboratory species [
4,
27]. In this regard, rat testes were reported to accumulate tin after three days of TBT exposure, resulting in reproductive abnormalities [
25]. TBT induced weight gain and improved the food efficiency of male rats [
51]. In another study [
52], TPT resulted in decreased sperm count and motility in a dose-dependent manner, an impaired sperm histone–protamine replacement process, and significantly increased incidence of sperm deformities, as well as impaired proliferation of spermatogonia in adult male rats. In a recent study, TBT exposure decreased the number of Leydig cells and inhibited androgen production in rats [
53]. In male mice, TBT exposure early after birth was associated with reduction in the weight of testis, epididymis, prostate, and seminal vesicles [
54,
55]. In zebra fish, TBT exposure during early development induced male bias and reduced or completely inhibited sperm motility, caused the absence of flagella, and resulted in only abnormal spermatozoa in semen in the exposed male offspring [
56]. Similarly, exposure of ACT to adult male and female zebra fish altered the gene expression related to reproductive function, such as for LH in pituitary and for aromatase in gonads, along with reductions in estrogen in both sexes [
57]. In addition, ACT exposure was associated with impaired spermatogenesis in males.In addition to in vivo studies, in vitro studies have shown that TPT exerted inhibitory effects on the activity of human blood steroidogenic enzymes, i.e., 5-alpha-reductase type 2, cytochrome P450 aromatase, 17-beta-HSD type 3, 3-beta-HSD type 2, and 17-beta-HSD type 1 through the interaction with critical cysteine residues [
50]. TBT, DBT and TPT salts were shown to inhibit human 5-alpha-reductase type 1 and 5-alpha-reductase type 2 enzymes, which are required for activation of androgens [
58]. In vitro acute or short-term exposure of bovine sperm to TBT was associated with decrease in total motility, progressive motility, curvilinear velocity, and beat-cross frequency, along with a lower mitochondrial membrane potential [
26]. In addition, TBT-exposed sperm resulted in a reduced cleavage rate and a lower rate of 8-16 cell morula development compared to embryos from unexposed sperms. DBT decreased the production of androgens from rat immature Leydig cells both under basal and LH stimulated conditions [
59]. In another study [
53], TBT exposure of immature rat Leydig cells in vitro reduced androgen production, cell viability, and cell cycle progression, while increasing reactive oxygen species (ROS) and apoptosis. Similarly, exposure of rat Leydig cells and Sertoli cells to TPT induced a significant decrease in the expression of steroidogenic and apoptotic indicators; similar exposure did not affect spermatogonia cells [
52].In female animals, organotin exposure was associated with toxicity of female reproductive system [
3,
27]. The most well known effect of many organotins, such as TBT and TPT, is the development of imposex in female gastropods, which refers to the development of male sexual organs in females, such as the penis and vas deferens [
60,
61,
62]. In many laboratory mammalian animal models, TBT exposure was associated with reproductive, metabolic, and cardiovascular abnormalities, including hyperandrogenism, cystic ovarian follicles, irregular estrous cycle, elevated levels of LH, obesity, abnormal lipid profiles, glucose metabolism, and insulin resistance similar to those found in polycystic ovarian syndrome (PCOS) in women and animal models of PCOS [
22,
63,
64,
65]. In this regard, TBT exposure in female rats induced body weight gain and adiposity [
66]. Exposure to TBT disrupts the proper functioning of the HPG axis of the female rats, probably in part through causing abnormal KISS and GnRH action, which regulate the reproductive axis of the hypothalamus and pituitary [
64]. In this regard, TBT exposure in female rats caused irregular estrous cycles, downregulated hypothalamic GnRH mRNA expression, decreased exogenous KISS response, decreased basal and surge levels of LH, reduced exogenous GnRH responsiveness, decreased pituitary expression of both ERα and Erβ, increased testosterone, ovarian and uterine fibrosis, and decreased fertility. In mice and rats, TBT disrupts ovarian reserve, development of germ cells, folliculogenesis, steroidogenesis, ovulation, and CL formation [
67]. In addition, in utero TBT exposure was associated with abnormal number and morphology of gonocytes with lipid droplets accumulating in the endoplasmic reticulum in the female rat offspring. In this regard, the placenta has been shown to accumulates the organotins, as exposure to TBT resulted in rat placental TBT levels that were five times higher than those found in maternal blood and ten times higher than those found in milk [
68]. In other studies, TBT caused irregular estrous cycles, disturbed ovarian development, including increased presence of atretic and cystic follicles, fewer CLs, antral follicles and increased levels of atretic follicles, hyperandrogenism, high levels of serum LH, and decreased levels of serum sex hormone-binding globulin, in addition to an increase in the RXR/PPAR signaling pathway and other proteins that are involved in androgen biosynthesis [
65,
69]. Exposure to ACT in adult female zebra fish was associated with alterations in the reproductive related gene expression, reduced estrogen, and increased testosterone in females [
57]. Increased accumulation of ACT in F1 eggs and embryonic abnormalities were also found after parental exposure.In vitro studies have also demonstrated effects of organotin on female hypothalamic-gonadal-axis and ovarian cells similar to in vivo effects. TBT exposure of ovarian theca cells from five species (human, sheep, cow, pig, and mouse) affected cholesterol trafficking, luteinization, and steroidogenesis in all five species [
70]. The effect was, in part, through modulation of RXR, as shown by RXR antagonist and RXRα knockdown. In human granulosa-like tumor cell line KGN, TBT exposure reduced the mRNA expression of aromatase and its activity by 30% compared to control cells [
71]. In addition, exposure to higher TBT concentrations resulted in KGN cell death within 24 h, whereas lower TBT concentration promoted apoptosis. Exposure of TBT in bovine cumulus–oocyte complex cultures reduced estrogen and testosterone levels along with expression of aromatase and 3-beta-HSD mRNA [
72]. In addition, TBT exposure inhibited LH stimulated estrogen synthesis in follicular granulosa cells. TBT and DBT acted as partial competitive inhibitors of aromatase enzyme in human placenta and showed inhibition of human 3-beta HSD type I activity [
73].
Taken together, the results of our in silico structural interactions of organotin ligands with sex steroid receptors support the previously reported adverse effects, as discussed above, for in vivo and in vitro studies in human and laboratory animals. The morphophysiological, hormonal, and molecular impairments discussed support our suggested hypothesis of perturbation of natural interaction between native ligands and sex steroid nuclear receptors leading to impaired reproductive function.


留言 (0)