
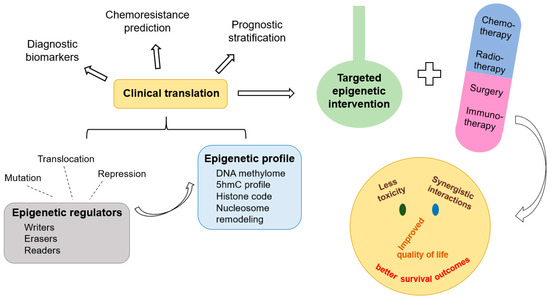
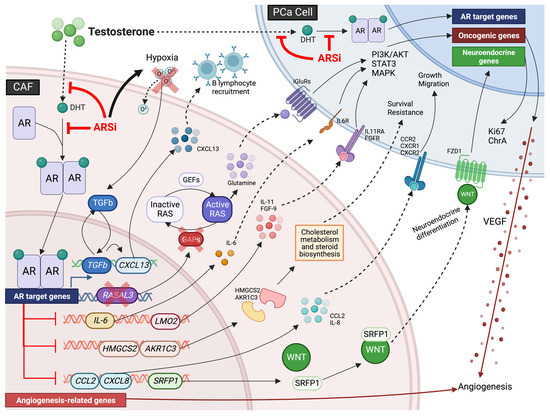
記住我
ncRNAs have been increasingly observed participating in hippocampal neurogenesis and functional reconstruction, including NSC regeneration, neural circuit repair, and recovery of learning and memory. This review summarizes progress in the understanding of the roles of ncRNAs in the regulation of hippocampal neurogenesis, particularly the differentiation of NSCs, and describes potential applications in the treatment of hippocampus-associated diseases, thus providing directions for future research and clinical practice.
3. Regulation of lncRNAs and the Differentiation of Hippocampal NSCslncRNAs are defined as a class of RNA molecules that do not encode proteins and whose transcripts are longer than 200 nucleotides. lncRNAs usually have the following characteristics: (1) lncRNA sequences are usually long and have an mRNA-like structure. (2) lncRNAs are not highly conserved, but the local sequences, and consequently their secondary and tertiary structures, are conserved; thus. lncRNAs are functionally conserved. (3) Expression of lncRNAs is relatively low. (4) Most lncRNAs show clear spatial–temporal expression specificity during differentiation and development. (5) Unlike miRNAs, lncRNAs do not have a universal mode of interaction. They can bind RNAs, DNAs, and proteins, and this binding can be enhanced or inhibited [92].lncRNAs influence gene expression at the epigenetic, transcriptional, and post-transcriptional levels, and consequently are involved in various cellular processes, including cell division, survival, apoptosis, and motility [93]. Their localization enables diverse functions. For example, lncRNAs can function as miRNA “sponges” in the cytoplasm by binding miRNAs and releasing them from their 3′UTR binding sites; consequently, mRNA stability is modulated, and miRNA activity and translation in the nucleus are inhibited. [94,95]. Nuclear lncRNAs regulate chromatin and gene transcription [96]. The regulation of lncRNAs by transcriptional variable splicing is currently unclear, and its relationship to the regulation of neural differentiation requires further study. Table 2 summarized the lncRNAs reported to be related with neurogenesis. 3.1. Rik-201 and Rik-203The lncRNA C130071C03Rik (mouse homologue of LINC00461) contains five transcripts (splice variants), among which the lncRNAs Riken (Rik)-201 and Rik-203 control neural differentiation. The lncRNA C130071C03Rik has been reported to be specifically expressed in the ependymal region of the spinal cord at embryonic days 11.5 and 13.5 in mice, and has been found to be expressed at higher levels in neural tissues than other tissues [97]. In comparison to other tissues, the hippocampus has an elevated level of Rik-203, which increases throughout neural development [98]. Recently, Zhang et al. found that Rik-203 and Rik-201 are present at high levels in the brain and promote mESC differentiation into neurons [99]. In addition, through RNA pull-down experiments, miR-467a-3p and miR-96 have been found to bind Rik-203 and Rik-201, respectively, thus indicating that Rik-203 and Rik-201 may serve as competing endogenous RNAs that inactivate miR-467a-3p and miR-96, respectively. Analysis using online miRNA target predication tools has indicated that Sox6 is a target of both miR-467a-3p and miR-96, and luciferase reporter assays have indicated that miR-467a-3p and miR-96 bind and downregulate Sox6 expression. Sox6 specifically regulates dopamine neurons during neural development and plays a regulatory role in the early differentiation of hippocampal NSCs [100,101]. Furthermore, gain or loss of function experiments have shown that Rik-203 and Rik-201 knockdown and overexpression of miR-96 and miR-467a-3p both inhibit neural differentiation. Sox6 reverses the suppression of differentiation in NSCs by upregulating miR-96 and miR-467a-3p and downregulating Rik-203 and Rik-201. These results indicate that Rik-203 and Rik-201 act as competing endogenous RNAs in the functional inhibition of miR-467a-3p and miR-96, respectively, and modulate the expression of Sox6, thereby further regulating NSC differentiation [99].Sevoflurane has been extensively used in clinical anesthesia, and early childhood exposure to sevoflurane has been associated with neural abnormalities in both humans and animals according to prior research [98,102]. Sevoflurane exposure potently inhibits NSC self-renewal and differentiation in vitro [103], thus leading to neuronal loss and cognitive impairment in young animals. Sevoflurane has also been found to decrease Rik-203 levels in mouse hippocampal tissue and neural progenitor cells. Suppression of Rik-203 levels in neural progenitors decreases not only Sox1 and Nestin expression, but also the number of Sox1 positive cells. miR-101a-3p strongly binds Rik-203, as evidenced by RNA-RNA pull-down experiments. Sevoflurane, Rik-203 suppression, and miR-101a-3p overexpression all decrease GSK-3β levels; consequently, a cascade of miR-101a-3p and GSK-3β may be involved in the Rik-203-mediated regulation of neural development [98]. Rik-203 also attenuates neuronal differentiation via inhibition of downstream miR-466l-3p. Sevoflurane decreases Rik-203 levels, thereby resulting in the release of miR-466L-3p from Rik-203. When miR-466L-3p is released, it specifically targets and decreases the concentration of BDNF and subsequently inhibits neuronal development [104]. The results above suggest that Rik-203 is a potential target that may be important in the prevention of anesthesia-induced neurotoxicity. 3.2. Peg13The lncRNA Peg13 was first reported to be differentially expressed in cerebral vascular endothelial cells in an ischemic glucose deficiency model in 2016, thus suggesting its potential role in nervous system injury [105]. Recently, Peg13 has been shown to regulate social and sexual interactions in mice, and mice lacking Peg13 exhibit same-sex attraction-like behavior. These mice also exhibit a deficiency in pup retrieval behavior, elevated anxiety, and diminished activity and curiosity [106]. Peg13 has been found to be involved in controlling mouse mating choices and modulating sevoflurane-associated neurotoxicity against NSCs [107,108]. Reports have indicated that an axis between miRNAs and target genes mediating Peg13 sponges is involved in the regulation of biological function. Exposure to sevoflurane significantly downregulates the expression of Peg13 and Sox13 and upregulates the expression of miR-128-3p in neural stem cells. Peg13 serves as a molecular sponge for miR-128-3p, thereby maintaining the expression of Sox13 in NSCs and decreasing sevoflurane-associated neurotoxicity [103]. Peg13 bound to miR-490-3p upregulates Psmd11, thus inactivating the Wnt/β-catenin pathway and alleviating the progression of epilepsy [107]. Mechanistically, Peg13 may function as a sponge for miR-20a-5p, thereby increasing the expression of XIAP and lessening serious brain injury caused by hypoxia/ischemia in newborn mice [109]. The Peg13/miRNA/target gene axis may have promising applications as a potential therapeutic target in neurological diseases. 3.3. lncRNA1230lncRNA1230 is a long intergenic non-coding RNA. Large-scale microarray data from Guttman et al. [107] indicate that the expression of lncRNA1230 is associated with differentiation of the neuroectoderm in mouse embryonic stem cells, thus suggesting its involvement in the control of neural lineage determination. Forced ectopic expression of lncRNA1230 significantly attenuates the ability of mouse embryonic stems to form neural cells, whereas knockdown of lncRNA1230 promotes the conversion of mouse embryonic stem cells toward certain NPCs. The lncRNA1230-induced mechanism of inhibition in the transformation of mouse ESCs to NPCs involves decreasing trimethylation of histone 3 lysine 4, thus inhibiting the binding of WD repeat domain 5 (WDR5) to the promoter regions of neurogenesis-related genes [108]. Notably, lncRNA1230 may play a crucial role in the neural fate of stem cells. 3.4. PnkyPnky is a highly conserved neuro-specific lncRNA localized primarily in the nucleus. In 2015, Ramos et al. [110] found that Pnky knockdown accelerates the differentiation of NSCs into mature neurons and significantly decreases the numbers of NSCs. Therefore, Pnky plays a critical regulatory role in NSC differentiation. Reports have indicated that Pnky inhibits the differentiation of NSCs by binding and interacting with polypyrimidine bundle binding protein (polypyrimidine tract-binding protein, PTBP1) and consequently inhibiting the splicing and expression patterns of key mRNAs in neuronal stem cells [111]. Pnky has also been shown to be a key regulator of NSC migration through modulation of the splicing and export of target mRNAs [112].Although lncRNAs play important roles in cell biology, few have been shown to regulate in vivo development, particularly cis and trans-regulation. In one example, after knockout of Pnky in developing cortical cells, the expression of POU Class 3 Homeobox 2 (Pou3f2) decreased and differentiation of NSCs was promoted, although the expression of Pou3f2 was not completely lost with the deletion of the Pnky gene. Pnky promotes the expression of Pou3f2 and regulates the differentiation of NSCs through trans-regulatory mechanisms [113].In-depth knowledge of Pnky may potentially allow for its use in the treatment of neural disorders. Lin et al. [114] designed and synthesized an MRI visualization nano-drug that immobilizes a mixture of siRNAs targeting Pnky on micelle surfaces and promotes the directional differentiation of NSCs into neurons by downregulating the level of Pnky, thus repairing cerebral infarctions. These results indicate the great potential of nanomedicines targeting Pnky in NSC-based therapies, particularly for stroke. 3.5. Neat1The overexpression and knockdown of the lncRNA Neat1 respectively promotes and inhibits both spinal cord NSC migration and differentiation into neurons [115]. Neat1 has been reported to be the key regulator of Wnt/β-catenin [116], the key pathway affecting the proliferation of NSCs [117]. The expression of Neat1 is regulated by miR-124, thus resulting in activation of the Wnt/β-catenin pathway during spinal cord injury regeneration [115]. Recent studies have indicated that the lncRNA Neat1 mediates the proliferation of neural stem cells via the Neat1-let 7 b-P21 axis [118]. Neat1 may also mediate several pathways regulating neurogenesis. Beyond promoting NSCs to differentiate into neurons, Neat1 promotes NSCs to differentiate into oligodendrocytes. In 2019, Katsel et al. found the diminished expression of Neat1 in the brains of patients with schizophrenia, with a greatly diminished number of oligodendrocytes also thus observed [119]. In terms of the above reports, there exists a feedback loop between lncRNA and miRNA, forming a mutual regulation relationship. As a competing endogenous RNA, miR-124 is the target of Neat1 and conversely controls its expression, and together they regulate downstream gene expression and signal activity. 3.6. GAS5The lncRNA GAS5 has been extensively studied in tumors and is thought to potentially function as an anti-oncogene [120]. Although research on the importance of GAS5 in neurological disorders such as AD is scarce, GAS5 levels are significantly higher in patients with AD than controls [121]. The function of GAS5 in neurogenesis has also been investigated, with GAS5 being found to promote hippocampal NSC differentiation into neurons. In rats with cholinergic injury, overexpression of GAS5 enhances learning and memory in vivo [122]. GAS5 was found to be regulated by Lhx8, which has a specific effect on the development of the cholinergic nervous system and promotes cholinergic differentiation of hippocampal NSCs. Therefore, it has been hypothesized that an elevated GAS5 level in the hippocampus promotes NSCs in the hippocampal DG to differentiate into neurons.Table 2. Effects and mechanisms of various lncRNAs on NSCs.
Table 2. Effects and mechanisms of various lncRNAs on NSCs.
NameBiological FunctionMechanismReferencesSox2OTInhibit NSC proliferation and promote neuronal differentiationLink with Sox2, interact with YY1[123]RMSTPromote the formation and development of nerve cellsTarget Sox2[124,125]Kdm2bPromote the formation and development of nerve cellsCombine with hnRNPAB and activate Kdm2b expression[126]PauparPromote the formation and development of nerve cellsCombine with local genes Pax6 and KAP1[127]Gm21284Inhibit NSC proliferation while promoting NSC differentiationInteract with miR-30e-3p, miR-147, and miR-431[128]1604Enhance neural differentiationmiR-200c/ZEB1/2 axis[129]Rik-201Promote neural differentiationRegulated by C/EBPβ and target miR-96/Sox6[99]Rik-203Promote neural differentiationRegulated by C/EBPβ and target miR-467a-3p/Sox6 and miR-101-3a/GSK-3β[98,99]Malat1Enhance neural differentiationActivate ERK/MAPK, inhibit PPAR/p53[130]PnkyInhibit neural differentiation and the formation and development of nerve cellsInteract with PTBP1[110]IncR492Inhibit neural differentiationInteract with HuR and activate Wnt signaling[131]BDNF-ASInhibit eNSC-derived neurite outgrowthTarget TrkB signaling pathway[132]UCA1Promote NSC differentiation to astrocytesmiR-1/Hes1[133]OPCBoost oligodendrogenesisRegulated by OLIG2[134]IncOL1Boost oligodendrogenesisForm a complex with Suz12[135]Inc158Boost oligodendrogenesisPromote NFIB expression[136]Neat1Boost oligodendrogenesisActivate the Wnt/β-catenin pathways[115]Pcdh17itFunction as an oligodendrogenesis markerUnknown[137]OLMALIN/-ASModulate oligodendrocyte maturationMay affect several genes, Target multiple genes, such as HDAC9, SOX4, GPR126 and EGR1[138]MAG3Inhibit neurogenesisAffect Notch or Wnt/β-catenin signaling pathway, miR-128-3p/ATRA/cAMP/CREB axis[139,140]lncRNA1230Attenuate NSCs’ ability to form neural cellsInteract with Wdr5[141]LINGO-1Promote neurogenesisDownregulate miR-15b-3p and promote Wnt5a expression[142]Peg13Promote neurogenesisSponge microRNA-128-3p to preserve Sox13 expression[103]GAS5Promote hippocampal NSC differentiation into neuronsUnknown[121,122] 4. Regulation of circRNAs in Hippocampal NSCscircRNAs are a newly described RNA species formed by the reverse splicing of linear genes. These stable closed circular RNA molecules are widely conserved and expressed in eukaryotes [143]. circRNAs act as molecular “sponges” for miRNAs and regulate gene transcription, affect protein synthesis and function, and participate in the regulation of intercellular signaling pathways [144]. Clustering of circular RNA sequencing data from different regions of the mouse brain (olfactory bulb, prefrontal cortex, hippocampus, and cerebellum) has indicated that expression of these RNAs is brain region-specific [145], and circRNAs are highly correlated with CNS diseases [146,147].lncRNAs, miRNAs, and circRNAs are inextricably linked; for example, circRNAs and lncRNAs can negatively regulate the expression of miRNA through an miRNA sponge mechanism, and can further interfere with the complementary base pairs of target mRNAs [148], thus modulating gene expression and interfering with the occurrence and development of disease [149]. Many ncRNAs do not play independent roles in cells but mutually interact and collaboratively influence biological functions. Table 3 summarized the circRNAs reported to be related with NSCs. 4.1. circHIPK2Wang et al. showed that circHIPK2 plays a negative role in the neural differentiation of NSCs. In vitro silencing of circHIPK2 causes direct differentiation of NSCs to neurons but not to astrocytes. Transplantation of NSCs with downregulated circHIPK2 has been found to facilitate neural plasticity and functional recovery after stroke; therefore, circHIPK2 may be a potential target for stroke treatment [150]. RNA sequencing has indicated that circHIPK2 targets and increases the expression of Smox. Silencing any of them facilitates neuronal differentiation in vitro but does not affect differentiation into astrocytes [150]. Thus, the effect on neuronal differentiation after silencing of circHIPK2 is potentially mediated by inhibition of the expression of Smox, thereby further promoting nerve recovery after stroke. However, the identities of the intermediary molecules between circHIPK2 and Smox remain elusive.miR-124 was proposed to have an intermediary role after Huang et al. discovered that circHIPK2 functions as an endogenous sponge for miRNA-124—a miRNA that regulates autophagy and endoplasmic reticulum stress [146]. Interestingly, miRNA-124 is involved in the regulation of neural differentiation [151,152], and Smox has been found to be the target gene of miRNA-124 in the development of H. pylori-associated gastric cancer [153]. The circHIPK2/miR-124/Smox axis is believed to play an important role in neurogenesis, although this possibility has not been confirmed experimentally. 4.2. circ-TTC3circRNA tetratricopeptide repeat domain 3 (circ-TTC3) has been studied in a limited number of diseases but is known to be upregulated in hypoxic cardiomyocytes and to protect against myocardial infarction-induced cardiomyocyte apoptosis via the miR-15b-Arl2 regulatory cascade [154]. Additionally, circ-TTC3 binds miR-449a, thus activating the NFⱪ-B and PI3K/AKT pathways and ameliorating hypoxia-induced damage in HaCaT cells [155]. Moreover, during acute kidney injury caused by sepsis, circ-TTC3 alleviates inflammation and oxidative stress via the miR-148a/Rcan2 axis [156].The effect of circ-TTC3 on NSC differentiation has been demonstrated by Yang et al. [157]: depletion of circ-TTC3 in NSCs increases proliferation and neuronal differentiation. circ-TTC3 appears to sponge miR-372-3p and then target and enhance Toll-like receptor 4 (TLR4) expression in NSCs. Evidence also indicates that TLR4 is involved in promoting NSC differentiation into astrocytes and neurons during stroke progression [158]. Depletion of circTTC3 was found to significantly decrease neurological scores, brain water content, and cerebral infarction in a middle cerebral artery occlusion/repression stroke model. Moreover, the effect of circTTC3 on stroke and NSC biology has been confirmed to be mediated through the miR-372-3p/TLR4 axis. 4.3. Acbd6The circRNA Acbd6 is spliced from exons derived from Acbd6 on the positive strand of chromosome 13 (from 73, 239, 821–73, 265, and 813 bp). Virtually no reports have investigated circAcbd6, and few studies have examined the host genes of circAcbd6. However, members of the ACBD family have been reported to play important roles in regulating viral replication, organelle organization, self-renewal of stem cells, and protein acylation [159]. A modular protein in mammalian cells, acyl-coenzyme A binding domain-containing member 6 (Acbd6) is found in the spleen, placenta, and embryonic-like stem cells produced from the placenta, bone marrow, cord blood, and circulating CD34+ progenitors [159,160]. In a recent study, a particular region of the SGZ that harbors a subpopulation of mature NSCs showed a substantial expression of circAcbd6 in neural tissues. Moreover, the finding from our lab shows that the forced expression of circAcbd6 promotes hippocampal NSC differentiation into neurons and even cholinergic neurons [161]. In vivo results suggest that circAcbd6 may greatly enhance learning and memory ability, indicating its potential value in the treatment of AD.Table 3. Effects and mechanisms of various circRNAs in NSCs.
Table 3. Effects and mechanisms of various circRNAs in NSCs.
NameBiologic FunctionMechanismReferencescircHIPK2Inhibit proliferation and differentiation of NSCsCombine miR-124 and regulate Smox expression[146]circRNA TTC3Inhibit proliferation and differentiation of NSCsRegulate miR-372-3p/TLR4 axis[157]hsa-circ-0002468Regulate neuronal differentiationModulate miR-561/E2F8 axis[162]circRNA Acbd6Promote NSC differentiation into cholinergic neuronsModulate miR-320-5p-Osbpl2 axis[161]circ-0005835Inhibit NSC proliferation and differentiation to neuronsSponge miR-576-3p[163] 6. Limitations and ProspectsCurrent research has utilized the characteristics and advantages of ncRNAs (especially miRNAs and circRNAs) as new biological markers to screen for several diseases. These ncRNAs have major roles in disease initiation and progression and great value in disease diagnosis, treatment, and prognostication. Interfering with the regulation of ncRNAs can help control disease progression; therefore, regulation of the expression of key ncRNAs may provide hope for disease prevention and treatment and the identification of targets for new drug development. At present, the future of ncRNA research is promising. Currently, in the treatment of neurological illnesses, ncRNAs can be regulated through a range of physical techniques, such as cranial magnetic resonance and electroacupuncture. However, recent studies have shown that the development of NSCs is a precisely controlled process whose influencing factors are not regulated by single signaling pathways but are instead closely associated with a variety of different cellular signaling pathways and cellular electrophysiological characteristics. The regulatory network is complex, involving chromatin remodeling, histone modification, DNA methylation, X chromosome inactivation, and regulation of ncRNA at the translational level. It is also associated with changes in the extracellular microenvironment. Recent studies have shown that ncRNAs play important roles in the differentiation of NSCs. However, several limitations must be noted. Research on the effects of ncRNAs on NSC differentiation has been relatively simple and limited to examining the effect of a certain type of ncRNA on NSC differentiation. The specific mechanisms remain to be fully elucidated, and some influencing factors remain hypothetical. In addition, few reports have described the formation of common regulatory networks among ncRNAs, mRNAs, and the microenvironment affecting the differentiation of NSCs. Furthermore, the effects of ncRNAs on the differentiation of NSCs involve multiple molecules. Although in recent years lncRNAs have been confirmed to act as miRNA sponges, few studies have assessed the effects of ncRNAs on the nervous system. Moreover, the construction of lncRNA-miRNA-mRNA regulatory networks is imperfect, and further research in this field is urgently needed. Nonetheless, ncRNAs are important regulators affecting the differentiation of NSCs and may enable the development of potentially valuable novel treatments for nervous system diseases.
留言 (0)